Full Length Research Paper

ABSTRACT
The foliar anatomy of 12 species of Asteraceae around Ile Ife in South Western, Nigeria was described. The distinguishing characteristics of taxonomic value include; venation patterns, areole shapes, number of veinlet endings, trichome types, arrangement of vascular bundles. Venation types are actinodromous, craspedodromous or camptodromous, the presence of rectangular areoles are predominant in the species studied and this show family characteristics. However, the presence of crystal druses in the areoles of C. odorata is of diagnostic importance. The type of vascular bundle in the leaf midribs is classificatory as it divided the taxa studied into two groups; vascular bundles are amphicribal in Bidens pilosa, Chromolaena odorata, Launaea taraxacifolia, Crassocephalum crepidiodes, Tridax procumbens and Vernonia cinerea and bicollateral in Ageratum conyzoides, Aspilia africana, Emilia praetermissa, Synedrella nodiflora, Tithonia diversifolia and Vernonia amygdalina.
Key words: Asteraceae, foliar, taxonomic, venation, areole.
INTRODUCTION
Asteraceae (Compositae) is a very large cosmopolitan family whose members are highly advanced. It belongs to the sub-class Asteridae in the order Asterales. Asteraceae is the second largest family in the division Magnoliophyta with 1,100 genera and over 20,000 recognized species (Ming, 1999). The majority of Asteraceae species are herbaceous although an important component of the family consists of shrubs or even trees, many plants in the family Asteraceae are economically important as weeds, ornamentals, medicinals and green vegetables (Olorode, 1984).
The angiosperms are endowed with macromorpho-logical characters of significant taxonomic values which can be easily observed with the naked eye or simple hand lens. Morphological attributes of vegetative organs have often constituted the mainstay of taxonomic studies in plants (Polhill, 1968; Pilbeam and Bell, 1979; Adedeji, 2005) and are very important in classification.
The use of anatomical methods in taxonomic investigations cannot be over emphasized. Although no character is absolutely immutable, but some are more fixed than the others and it is on those that are less plastic that the systematic anatomist rely. Taxonomic decision based on epidermal studies are quite reliable because they are not really affected by environmental conditions (Barthlott, 1981), thus comparative plant epidermal studies have been found to be reliable in taxonomy and systematics (Stace, 1969; Ogunkunle and Oladele, 2000; Metcalfe and Chalk, 1950, 1979; Naik and Nigrude, 1981; Palmer and Tucker, 1981; Adedeji, 2004; Adedeji and Illoh, 2004) all the authors have all stressed the taxonomic importance of anatomical features which along with other characters form taxonomic, identification and classification of plants.The present study reported the use of foliar anatomy in establishing the taxonomic relationships between twelve species of Asteraceae.
MATERIALS AND METHODS
Leaf clearing (for venation studies)
Sizeable portions of the matured leaves of each species were taken from the standard median portion (midway between the tip and the base) of Ageratum conyzoides Linn., Aspilia africana (Pers) C.D Adams, Bidens pilosa Linn., Chromolaena odorata (Linn.) king & Robinson, Crassocephalum crepidioides Benth S. Moore, Emilia praetermissa Milne-Redhead, Launaea taraxacifolia Willd, Synedrella nodiflora Gaertn, Tithonia diversifolia (Hemsl) A. Gray, Tridax procumbens Linn., Vernonia amygdalina Del. Cent and Vernonia cinerea Linn. were obtained from the median parts of well expanded leaves.
These portions were decolorized by boiling in 90% ethyl alcohol to remove chlorophyll, and treated in 2% sodium hydroxide solution over night. They were rinsed in water thrice and transferred into 5% domestic bleach (JIK). They remained in the bleach until they became completely white, then rinsed in water thrice and preserved in 50% ethanol as described by Olatunji (1983). These were stained in 1% aqueous solution of Safranin O and mounted on a clean slide in 25% glycerol and the edges of the cover slips sealed with nail vanish.
Sectioning
Transverse sections of the leaf were cut at 20 µ thickness using Reichert Sledge Microtome and best sections preserved in 50% ethanol. The sections were stained in 1% aqueous solution of Safranin O for 5 min washed in 3 changes of water to remove excess stain and counterstained in 1% solution of Alcian blue for 5 min then washed in three changes of water and dehydrated by passing through series of ethyl alcohol: 50, 70, 80, 90 and 100% with two changes in 100% alcohol (dehydration process), and excess stain (differentiation process).
The dehydrated and differentiated sections were cleared in xylene to remove last trace of water, to clear the sections (making it more transparent) and to remove last traces of ethanol and since xylene is the solvent of the mountant (DPX) used, it prevent cloudiness of the slide. The sections were mounted in DPX mountant according to Akinloye et al. (2012). Photomicrographs of the leaf sections of the twelve species were taken with Amscope digital camera attached to a light microscope. All microscopic measurements were made with the aid of an ocular and stage micrometer. The drawings were prepared in the scientific illustration unit of the National History museum, Obafemi Awolowo University by the museum’s scientific illustrator with about twenty eight years experience in scientific illustrations. The slides were observed directly under LEICA DM500 binocular light microscope using different magnifications, the drawings were prepared without the aid of Camera Lucida but with painstaking replication to scale of the arrangement of the cells and structures in pencil line drawings before being transferred unto tracing films using Rotring pen size 0.1 with black Rotring ink. The drawings were later scanned into the computer for insertion into the appropriate sections in the text.
RESULTS
Herbarium survey
All plant species used in this work were collected authenticated in the Ife Herbarium and voucher specimens deposited in the same herbarium (Table 1).

Ageratum conyzoides Linn.
Venation is actinodromous, perfect, marginal, basal. Areoles arrangement is random. The shape is triangular, quadrangular and pentagonal; 38.4 - 86.3±1 µm long and 24.9 - 92.3±1 µm wide. The veinlet endings are singly divided and number ranges between 0 - 2. Lamina (Plate 1A and B): Epidermis uniseriate, cells rectangular. The cuticle on both surfaces are not prominent and where they are seen, they are non-striated. Palisade mesophyll one layer thick, consisting of cylindrical shaped parenchyma cells that are closely packed. Mean length of palisade mesophyll is up to 28.9±1.80 µm. Spongy mesophyll consist of parenchyma cells that are largely irregular in shape, irregularly arranged with intercellular spaces. Simple uniseriate multicellular trichome present. Midrib (Plate 1C and D): Epidermis uniseriate, the epidermal cells are irregular, occasionally polygonal. Vascular bundle, 1, bicollateral. Simple uniseriate multicellular trichome present.

Aspilia africana (Pers). C.D. Adams
Venation is actinodromous, basal to suprabasal. Areoles shape is rectangular and quadrangular; 36.2 - 92.7 µm ± 1 long and 49.7- 102.6 µm ± 1 wide. The veinlet endings are bifurcated number ranges between 0 - 2. Lamina (Plate 2A and B): Epidermis uniseriate, cells rectangular, cuticles are not prominent and non-striated. Palisade mesophyll one layer of cells thick consisting of tightly packed elongated cells. Mean length of palisade mesophyll up to 34.5±1.80 µm. Spongy mesophyll consist of parenchyma cells that are largely irregular in shape, cells irregularly arranged with intercellular spaces. Midrib (Plate 2C and D): Epidermis uniseriate, epidermal cells are largely irregular in shape, occasionally polygonal varying in size and arrangement. Vascular bundles 5, crescentiform with dissected xylem, bicollateral. Trichome present, largely simple uniseriate bicellular.

Bidens pilosa Linn.
Venation is pinnate, craspedodromous, that is, secondary veins terminating at the margin. Areoles are variables in size, polygonal to rectangular in shapes; 35.2 - 82.3 ± 1 µm long and 38.4 - 98.8 ± 1 µm wide. Veinlet endings are singly divided, number ranges between 1-3. Lamina (Plate 3A and B): Epidermis uniseriate, cells rectangular shaped. Not prominent and non-striated cuticles on both surfaces. Palisade mesophyll one layer thick, consisting of tightly packed elongated shaped parenchyma cells. Mean length of palisade mesophyll up to 29.8±1.70 µm. Spongy mesophyll consist of loosely packed varied, irregularly shaped parenchyma cells with intercellular spaces. Midrib (Plate 3C and D): Epidermis uniseriate, epidermal cells are irregular polygonal varying in sizes and arrangements. Vascular bundle 3, trapezoid amphicribal type. Simple uniseriate multicellular trichome present.

Chromolaena odorata Linn.
Venation is actinododromous, that is, 3 or more primary veins diverging radially from a single point, suprabasal. Areole shapes are rectangular, polygonal and triangular; 79.1 - 149.2±1 µm long and 64.4 - 93.8±1 µm wide. Veinlet endings are singly divided, bifurcated and number ranges between 1 to 3. Crystal druses are present and prominent. Lamina (Plate 4A and B): Epidermis uniseriate, cells rectangular. The cuticles on both surfaces are not prominent, where they seen, they are non-striated. Palisade mesophyll one layer thick consisting of tightly packed cylindrical elongated parenchyma cells. Mean length of palisade mesophyll up to 27.5±1.80 µm. Spongy mesophyll consist of parenchyma cells largely irregular in shape, irregularly arranged with intercellular spaces. Simple uniseriate bicellular and multicellular trichomes are present. Midrib (Plate 4C and D): Epidermis uniseriate, epidermal cells are irregular variable in size and arrangement. Vascular bundle, 1, amphicribal, shield shaped. Trichome: Simple uniseriate multicellular trichome are present.

Crassocephalum crepidioides (Benth) S. Moore
Venation is pinnate, craspedodromous, that is, secondary veins terminating at the margin. Areole shapes are rectangular to quadrangular; 19.8 - 83.2 ± 1 µm long and 40.7 - 87.9 ± 1 µm wide. Veinlet endings are singly divided, bifurcated and number ranges between 0 - 2. Lamina (Plate 5A and B): Epidermis uniseriate, epidermal cells are irregular in sizes and arrangement. Cuticles are not prominent and non-striated on both surfaces. Palisade mesophyll one layer thick consisting of tightly packed cylindrical elongated parenchyma cells. Mean length of palisade mesophyll up to 29.7±1.80 µm. Spongy mesophyll consist of parenchyma cells largely irregular in shape, irregularly arranged with intercellular spaces. No trichome present. Midrib (Plate 5C and D): Epidermis uniseriate, epidermal cells are polygonal to rectangular shaped varying in size and arrangement. Vascular bundle, 2, amphicribal, dissected xylem. No trichome present.

Emilia praetermissa Milne-Redhead
Venation is pinnate, camptodromous, that is, secondary veins freely ramified towards the margin. Areoles are rectangular and hexagonal; 42.9 - 89.3±1 µm long and 36.2 - 68.9±1 µm wide. Veinlet endings are singly divided, bifurcated, number ranges between 0 - 2. Lamina (Plate 6A and B): Epidermis uniseriate, epidermal cells are rectangular shaped. Cuticles are not prominent and non-striated. Palisade mesophyll one layer thick consisting of tightly packed elongated cylindrical cells. Mean length of palisade mesophyll is up to 23.6±1.60 µm. Spongy mesophyll consist of parenchyma cells largely irregular in shape, irregularly arranged with intercellular spaces. No trichome present. Midrib (Plate 6 C and D): Epidermis uniseriate, epidermal cells are polygonal to rectangular in shape variable in size and arrangement. Vascular bundle, 1, bicollateral. No trichome present.

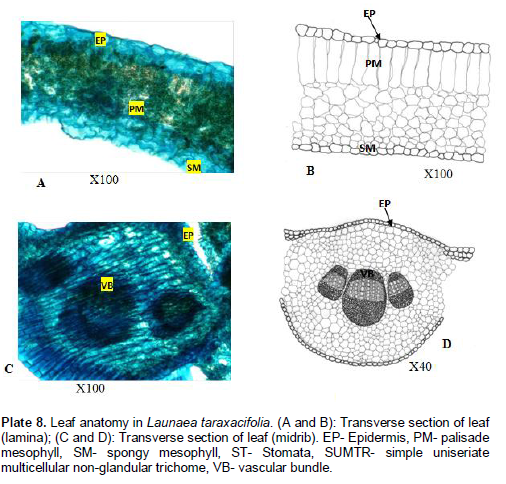
Launaea taraxacifolia Willd
Venation is pinnate, craspedodromous, that is, secondary veins terminating at the margin. Areole shapes are rectangular to polygonal; 50.9 - 99.2±1 µm long and 36.2 - 88.9±1 µm wide. Veinlet endings are bifurcated and number ranges between 0 - 2. Lamina (Plate7A and B): Epidermis uniseriate, epidermal cells more or less rectangular. Cuticles are not prominent and non-striated. Palisade mesophyll one layer of cells thick consisting of tightly packed elongated cylindrical parenchyma cells. Mean length of palisade mesophyll is up to 26.8±1.60 µm. Spongy mesophyll consist of parenchyma cells enclosing intercellular spaces. No trichome present. Midrib (Plate 7C and D): Epidermis uniseriate, epidermal cells are rectangular. Vascular bundle 3, bicollateral. No trichome present.

Synedrella nodiflora Benth
Venation is pinnate, actinodromous; that is, 3 or more primary veins diverging radially from a single point, perfect, marginal, basal. Areole shape is quadrangular; 31.6- 106.2±1 µm long and 28.7 - 96.7±1 µm wide. Veinlet endings are singly divided and number ranges between 0 - 2. Lamina (Plate 8A and B): Epidermis uniseriate, epidermal cells are largely irregular. Not prominent, non-striated cuticles. Palisade mesophyll two layers of cells thick, tightly short or elongated packed cylindrical parenchyma cells. Mean length of palisade mesophyll up to 26.8±1.65 µm. Spongy mesophyll consist of loosely packed cells enclosing large intercellular spaces. No trichome present. Midrib (Plate 8C and D): Epidermis uniseriate, epidermal cells are largely irregular in sizes and arrangements, occasionally polygonal. Vascular bundle 17, bicollateral. No trichome present.
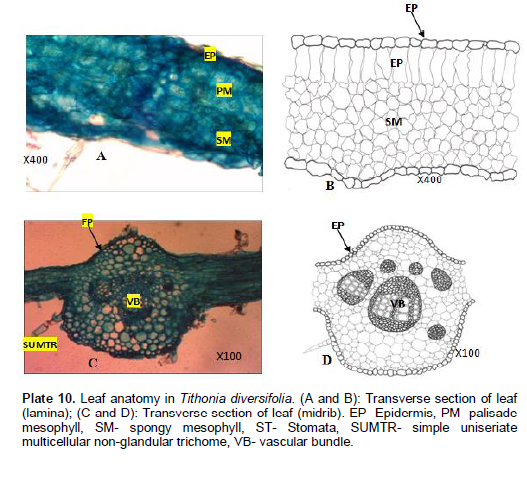
Tithonia diversifolia Hemsl A. Gray.
Venation is actinodromous, that is, 3 or more primary veins diverging radially from a single point, basal. Areole shape varies from rectangular to polygonal; 55.4 - 94.9±1 µm long and 45.2 - 87.9±1 µm wide. Veinlet endings are singly divided and number ranges between 0 - 2. Lamina (Plate 9A and B): Epidermis uniseriate, cells are irregular varying in sizes and arrangement. Cuticles are not prominent, non-striated. Palisade mesophyll one layer thick,tightly packed, cylindrical elongated cells. Mean length of palisade mesophyll up to 27.7±1.70 µm. Spongy meso-phyll layer consist of tightly packed cells enclosing inter-cellular spaces. No trichome present. Midrib (Plate 9C and D): Epidermis uniseriate, cells are irregular, cuticle not distinct. 2 large, 4 small, bicollateral bundles. Simple uniseriate multicellular trichome present.

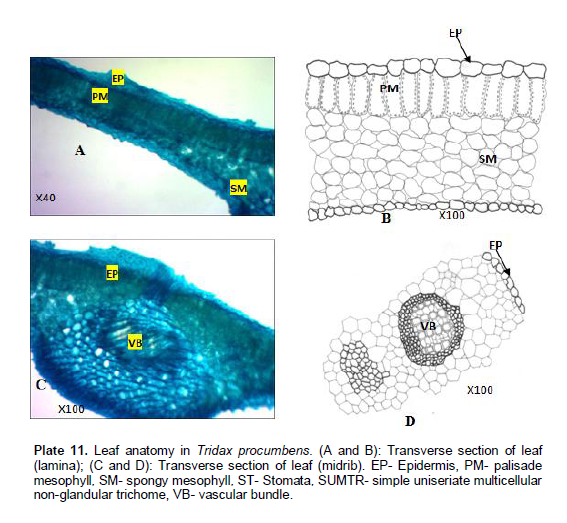
Tridax procumbens Linn
Venation is pinnate, camptodromous, cladodromous, that is, secondary veins freely ramified towards the margin. The shape varies from rectangular to polygonal; 54.5 - 97.7±1 µm long and 46.8 - 82.5±1 µm wide. Veinlet endings are singly divided and number ranges between 0 - 2. Lamina (Plate 10A and B): Epidermis uniseriate, cells more or less elongate or cylindrical. Cuticles are not prominent and where observed, they are non-striated. Palisade mesophyll one layer of cells thick, tightly packed, cylindrical and elongated cells. Mean length of palisade mesophyll up to 28.7±1.60 µm. Spongy mesophyll consist of loosely packed irregular cells enclosing intercellular spaces. No trichome present. Midrib (Plate 10C and D): Epidermis uniseriate, cells are irregular vascular bundle, 1, amphicribal. No trichome present.
Vernonia amygdalina Del. Cent
Venation is pinnate, camptodromous, brochidodromous, that is, secondary veins joined together in a series of prominent arches. The areoles are variable in size and shape rectangular to quadrangular in shape; 54.5 - 103.7±1 µm long and 35.6 - 97.4±1 µm wide. Veinlet endings are singly divided and number ranges between 0 - 2. Lamina (Plate 11A and B): Epidermis uniseriate, cells polygonal to irregular, variable in size and arrangement. Cuticles not prominent and non-striated. Palisade mesophyll, one layer of cells thick, tightly packed. Mean length of palisade mesophyll up to 29.6 ± 1.50 µm. Spongy mesopyhll cells consist of loosely packed irregular cells enclosing intercellular spaces. No trichome present. Midrib (Plate 11C and D): Epidermis uniseriate, epidermal cells variable in sizes and arrangement. Vascular bundle, 5, bicollateral. No trichome present.
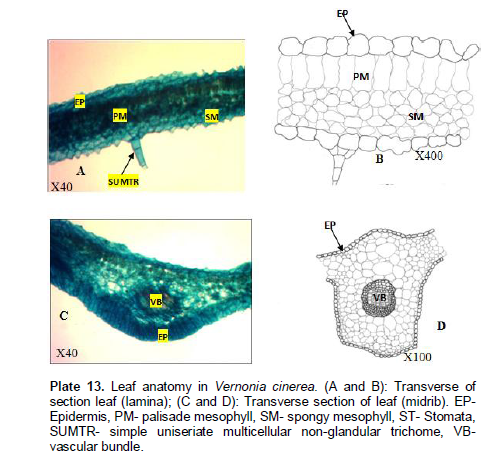
Vernonia cinerea Linn
Venation is pinnate, camptodromous, cladodromous, that is, secondary veins freely ramified towards the margin. Areoles are variables in size. The shapes vary from polygonal to rectangular; 65.5 - 118.7±1 µm long and 48.6 - 81.4±1 µm wide. Veinlet endings are simple, bifurcated and number ranges between 0 - 4. Lamina (Plate 12A and B): Epidermis uniseriate, cells are irregular to polygonal, variable in size and arrangement. Cuticles are not prominent and non-striated. Palisade mesophyll, one layer thick consisting of tightly packed elongated cylindrical parenchyma cells. Mean length of palisade mesophyll up to 28.4±1.70 µm. Spongy mesophyll cells consist of irregular cells enclosing intercellular spaces. No trichome present. Midrib (Plate 12C and D): Epidermis uniseriate, cells are irregular in shape, thin cuticle. Vascular bundle, 1 amphicribal. No trichome present.

DISCUSSION
A survey of literature of leaf anatomy shows that data obtained from it can be used amply for the clarification of taxonomic and phylogenetic relationships. The commonly used characters like palisade and spongy features, types of vascular bundles, trichome types were largely employed in this study. In all the species studied, the major or primary veins are pinnate. Venation types were actinodromous, craspedodromous or camptodromous, the presence of rectangular areoles were predominant in the species studied and this show family characteristics. However, the presence of crystal druses in the areoles of C. odorata is of diagnostic importance. The veinlet terminating end per areole also varies from one species to another, ranges from 0-2 in A. conyzoides, A. africana, C. crepidioides, E. praetermiissa L. taraxacifolia, S. nodiflora and V. amygdalina. For these taxa, number of veinlet endings is classificatory though there is an overlap. The veinlet endings divided singly or bifurcated. The general significant difference observed in the quantitative characters may be used to separate the species. C. odorata has a higher areolar area than the other species while V. cinerea has a higher number of veinlet endings than others (Plate 13A, B, C and D).
Leaf anatomy according to Carlquist (1961) provides a variety of features that could be used for taxonomic purposes. Many researchers have utilized leaf anatomical features for taxonomic consideration in many species of plants. These include the works of Naik and Nigrude (1981) on Chlorophytum, Ogundipe and Olatunji (1991) on Cochlospermum, Illoh (1995) on Celosia, Adedeji (2004) on Emilia and Adedeji and Illoh (2004) on Hibiscus. Though there is uniformity in the anatomy of the leaf in the species studied, characters like the shape of palisade and spongy mesophyll cells, non-prominence and non-striation of the cuticles and types of vascular bundles could be used to classify members of the family. The type of vascular bundle in the leaf midribs is classificatory as it divided the taxa studied into two groups; vascular bundles are amphicribal in B. pilosa, C. odorata, L. taraxacifolia, C. crepidiodes, T. procumbens and V. cinerea and bicollateral in A. conyzoides, A. africana, E. praetermissa, S. nodiflora, T. diversifolia and V. amygdalina. Similarly, the shape of vascular bundle is classificatory in those with dissected xylem, that is, A. africana and C. crepidiodes and those without dissected xylem, this delimit the two from other species studied. Among all the taxa studied, S. nodiflora is quite distinct in the surface view of the epidermis by having deep groove and numerous number of bundles.
CONFLICT OF INTERESTS
The author(s) have not declared any conflict of interests.
REFERENCES
| Adedeji O (2004). Leaf epidermal studies of the species of Emilia Cass. (Senecioneae, Asteraceae) in Nigeria. Botanical Lithuanica 10(2):121-133. | ||||
| Adedeji O (2005). Pollen morphology of the three species of the genus Emilia Cass. (Asteraceae) from Nigeria. Thaiszia - J. Bot. 15: 1-9. | ||||
| Adedeji O, Illoh HC (2004). Comparative foliar anatomy of ten species in the genus Hibiscus Linn. in Nigeria. New Botanists 31:147-180. | ||||
| Akinloye AJ, Illoh HC, Olagoke OA (2012) Significance of wood anatomical features to the Taxonomy of five Cola species. Sustain. Agric. Res. 1(2):21-26. | ||||
|
Barthlott W (1981). Epidermal and seed surface characters of plants: Systematic applicability and some evolutionary aspects. Nordic J. Bot. 1:345-355. Crossref |
||||
| Carlquist (1961). Comparative Plant anatomy. Holt, Rinehart and Wintson, New York. | ||||
| Illoh HC (1995). Foliar epidermis and petiole anatomy of four species of Celosia L. in Nigeria. Feddes Repertorium 106(1-2):15-23. | ||||
| Metcalfe CR, Chalk L (1950). Anatomy of the Dicotyledons Vol. I and II, Oxford, Clanredon Press. | ||||
| Metcalfe CR, Chalk L (1979). Anatomy of Dicotyledons 2nd ed. Vol. 1 Clarendon Press, Oxford. | ||||
| Ming LC (1999). Ageratum conyzoides, A tropical source of medicinal and agricultural Products. In: J. Janick (Ed.) Perspectives on New Uses. ASHS Press, Alexandria V.A. pp. 469-473. | ||||
| Naik VN, Nigurde SM (1981). Anatomy in relation to taxonomy of Chlorophytum (Liliaceae). Indian J. Bot. 4(2):48-60. | ||||
| Ogunkunle ATJ, Oladele FA (2000). Diagnostic value of trichomes in some Nigerian species of Ocimum Hyptis Jazq and Tinnea kotschy and Peys (Lamiaceae). J. Appl. Sci. 3:1163-1180. | ||||
| Olatunji OA (1983). Practical manual for Plant Anatomy. A laboratory practical manual Obafemi Awolowo University, Ile Ife, Nigeria (manuscript). | ||||
| Olorode O (1984). Taxonomy of West Africa Flowering Plants. Longman Group Ltd., London and New York. pp. 98-100. | ||||
| Palmer G, Tucker AE (1981). A Scanning Electron Microscope Survey of the Epidermis of East Africa Grasses I. Smithsonian Contribution to Botany: U.S.A. No 43. | ||||
|
Pilbeam DJ, Bell EA (1979). A reappraisal of the free amino acids in seeds of Crotalariajuncea (Leguminosae). Phytochem. 18(2):320-321. Crossref |
||||
|
Polhill RM (1968). Miscellaneous notes on African species of Crotalaria L. II. Kew bulletin 22:169-348. Crossref |
||||
|
Stace CA (1969). The significance of the leaf epidermis in the taxonomy of the Combretaceae II, the genus Combretum, sub-genus Combretum in Africa. Bot. J. Linn. Soc. 62:131-168. Crossref |
||||
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


