Full Length Research Paper

ABSTRACT
Proteinase inhibitors are a group of defense related proteins, natural antagonists of proteinases, induced upon herbivory, play a defensive role against polyphagous insects and phytopathogens. Serine proteinase inhibitor isolated from Cocculus hirsutus (L.) Diels, Cocculus hirsutus trypsin inhibtor (ChTI) was found effective as antifungal, bactericidal and nematicidal agent. Tomato plants expressing ChTI have been developed by agro-infection with almost 27% transformation efficiency. Stable integration and expression of ChTI has been established by polymerase chain reaction (PCR), inhibitory assay and western blot assay. Transgenic plants showed increased fruit yield, antioxidants, phenolics, flavonoids and titratable acidity. Protein extracts of tomato plants inhibited Helicoverpa armigera (gut proteinases up to 40%. Transgenic plants MT2 and JT2 challenged with 2nd and 4thinstar H. armigera (Hubner) larvae, showed delayed larval growth with 100% mortality. The results put together suggest that ChTI is a potential candidate for developing transgenic plant with multiple biotic stress tolerance.
Key words: ChTI, Helicoverpa armigera, insect bioassay, transgenic tomato, trypsin inhibitory assay.
INTRODUCTION
MATERIALS AND METHODS
RESULTS
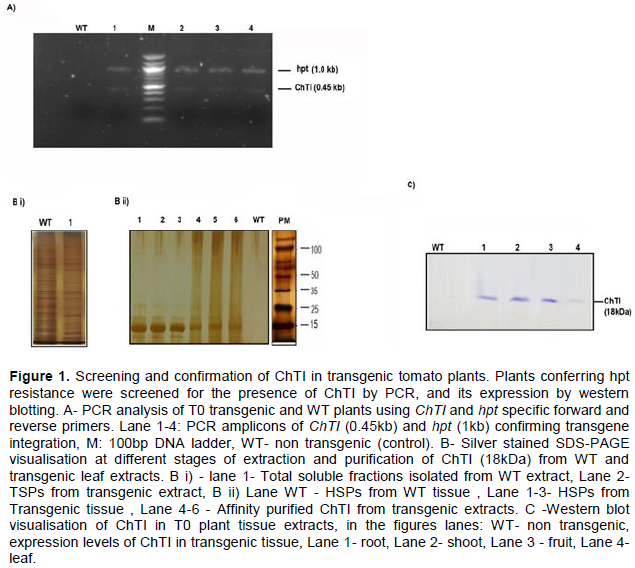
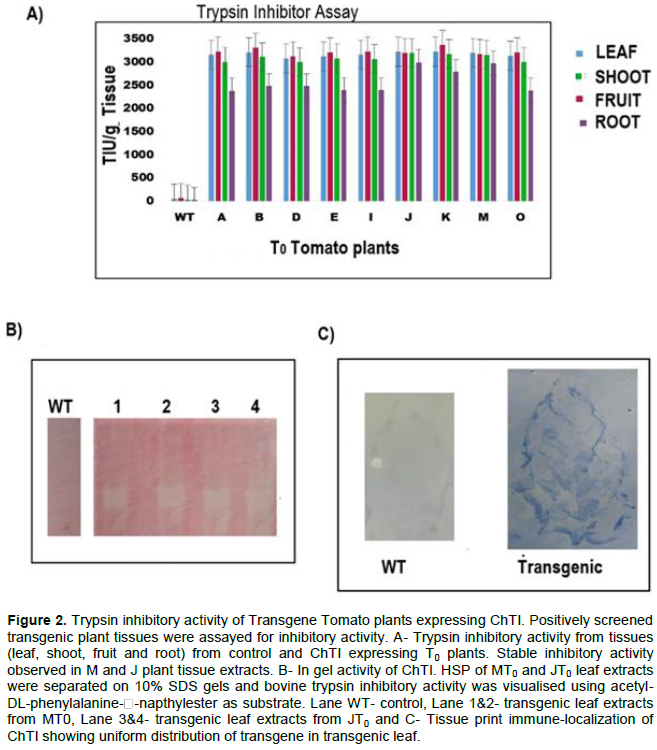
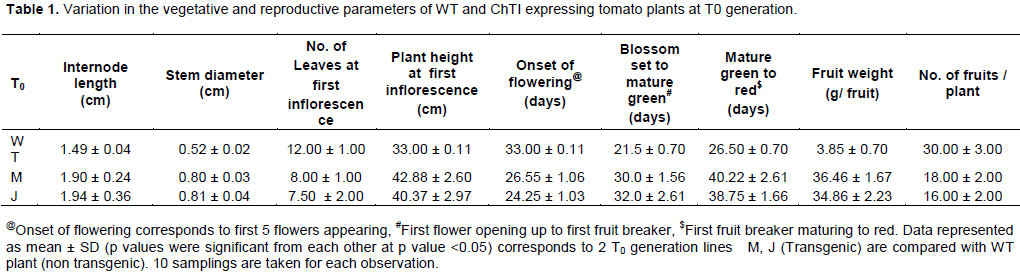
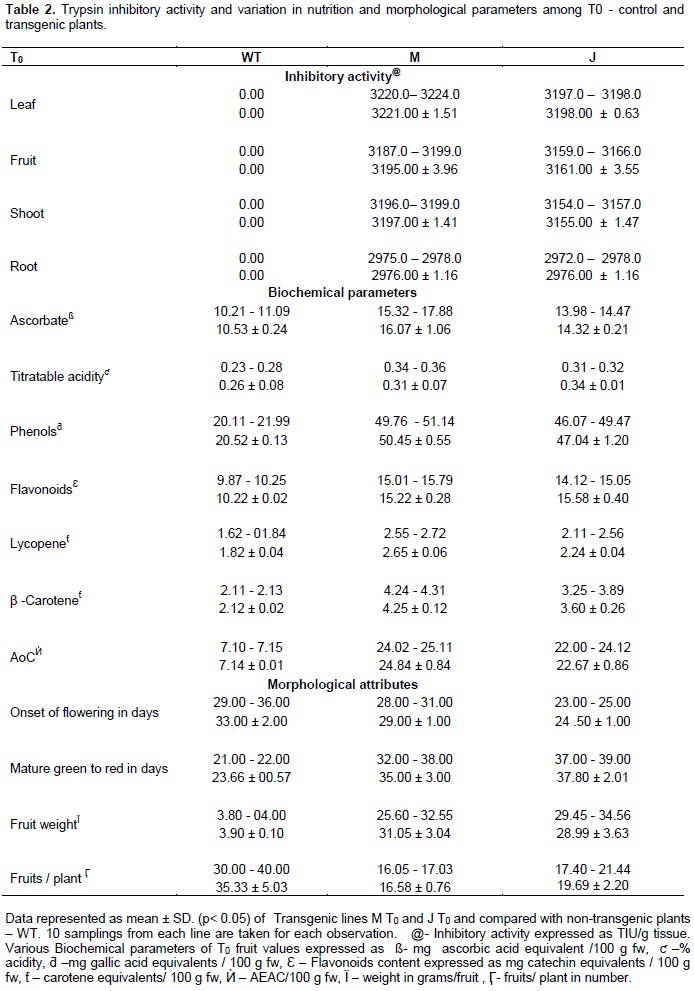
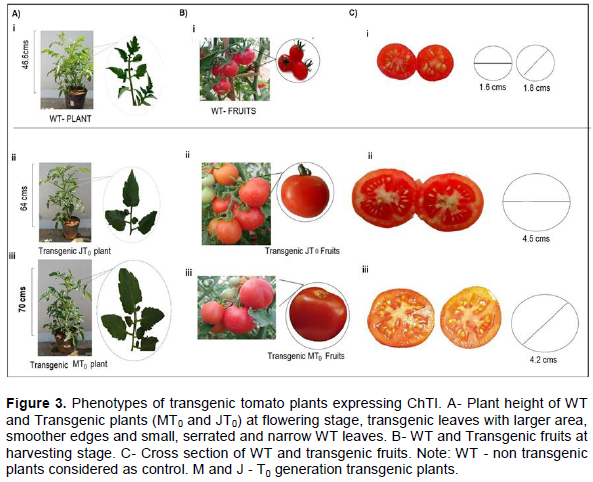

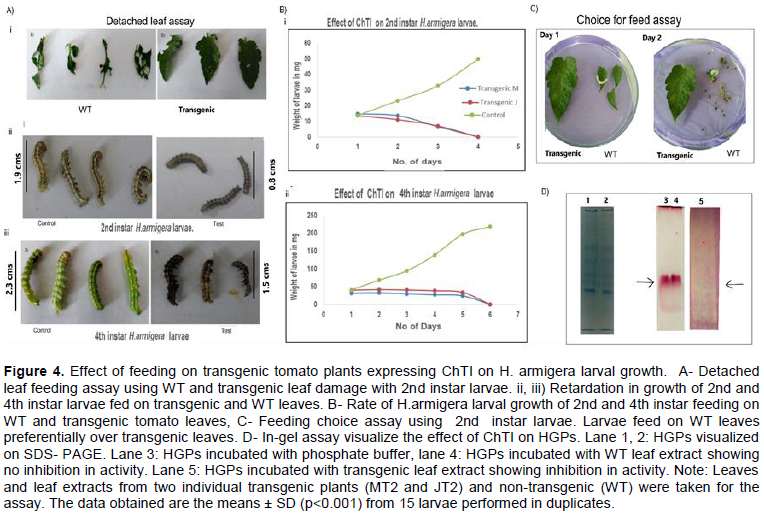
DISCUSSION
CONCLUSION
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ABBREVIATIONS
ACKNOWLEDGEMENTS
REFERENCES
|
Anderson P, Anton S (2014). Experience-based modulation of behavioral responses to plant volatiles and other sensory cues in insect herbivores. Plant Cell Environment 7:1820-1835. |
|
|
AOAC Ascorbic acid, in Official Methods of Analysis (2006). AOAC International, Gaithersburg, MD. 967.21. |
|
|
AOAC, Titratable acidity of fruit products, in Official Methods of Analysis. (2000). 17 edn- AOAC International, Gaithersburg MD. 942.15. |
|
|
Bacha AB, Jemel I, Moubayed NMS (2017). Purification and characterization of newly serine protease inhibitor from Rhamnus frangula with potential for use as therapeutic drug 3. Biotech 7:1-13. |
|
|
Benzie IF, Strain JJ (1996). The ferric reducing ability of plasma (FRAP) as a measure of 'antioxidant power': the FRAP assay. Annals of Biochemistry 23:70-76. |
|
|
Bhattacharjee C, Manjunath NH, Prasad DT (2010). Purification of a trypsin inhibitor from Cocculus hirsutus and identification of its biological activity. Journal of Crop Science and Biotechnology 12:253-260. |
|
|
Burge J, Mickelsen O, Nicklow C, Marsh LG (1975). Vitamin C in tomatoes: Comparison of tomatoes developed for mechanical or hand harvesting. Ecology of Food Nutrition 4:27-31. |
|
|
Chikate YR, Tamhane VA, Joshi RS, Gupta VS, Giri AP (2013). Differential protease activity augments polyphagy in Helicoverpa armigera. Insect Molecular Biology 22:258-272. |
|
|
Clemente M, Corigliano MG, Pariani SA, Sánchez-López EF, Sander VA, Ramos-Duarte VA (2019). Plant serine protease inhibitors: Biotechnology application in agriculture and molecular farming. International Journal of Molecular Sciences 20:1345-1366. |
|
|
Cloutier C, Jean C, Fournier M, Yelle S, Michaud D (2000). Adult Colorado Potato Beetles, Leptinotarsa decemlineata compensate for nutritional stress on Oryzacystatin I transgenic potato plants by hypertrophic behavior and over-production of insensitive proteases. Archives of Insect Biochemistry 44:69-81. |
|
|
De Leo F, Volpicella M, Licciulli F, Liuni S, Gallerani R, Ceci LR (2002). PLANT-PIs: A database for plant protease inhibitors and their genes. Nucleic Acids Research 30:347-348. |
|
|
Doyle JJ, Doyle JL (1987). A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochemical Bulletin 19:11-15. |
|
|
Dunse KM, Stevens JA, Lay FT, Gaspar YM, Heath RL, Anderson MA (2010). Co-expression of potato type I and II proteinase inhibitors gives cotton plants protection against insect damage in the field. Proceedings of the National Academy of Sciences, USA 107:15011-15015. |
|
|
Filho JX, Moreira RDA (1978). Visualization of proteinase inhibitors in SDS-polyacrylamide gels. Annals of Biochemistry 84:296-299. |
|
|
Fluhr R, Lampl N, Roberts TH (2012). Serpin protease inhibitors in plant biology. Physiologia Plantarum 145:95-102. |
|
|
Gatehouse JA (2011). Prospects for using proteinase inhibitors to protect transgenic plants against attack by herbivorous insects. Current Protein and Peptide Science 12: 409-416. |
|
|
Ghorbani S, Hoogewijs K, PeÄenková T, Fernandez A, Inzé A, Eeckhout D, Kawa D, De Jaeger G, Beeckman T, Madder A, Breusegem FV, Hilson P (2016). The SBT6.1 subtilase processes the GOLVEN1 peptide controlling cell elongation. Journal of Experimental Botany 67:4877-4887. |
|
|
Giri AP, Harsulkar AM, Deshpande VV, Sainani MN, Gupta VS, Ranjekar PK (2002). Chickpea defensive proteinase inhibitors can be inactivated by pod borer gut proteinases. Plant Physiology 116:393- 401. |
|
|
Golla SK, Rajasekhar P, Akbar SMD, Sharma HC (2018). Proteolytic activity in the midgut of Helicoverpa armigera (Noctuidae: Lepidoptera) larvae fed on wild relatives of chickpea, Cicer arietinum. Journal of Economic Entomology 111:2409-2415. |
|
|
Goulet MC, Dallaire C, Vaillancourt LP, Khalf M, Badri MA, Preradov A, Duceppe MO, Goulet C, Cloutier C, Michaud D (2008). Tailoring the specificity of a plant cystatin toward herbivorous insect digestive cysteine proteases by single mutations at positively selected amino acid sites. Plant Physiology 146:1010-1019. |
|
|
Gutiérrez-Campos R, Torres-Acosta JA, Pérez-Martínez JDJ (2001). Pleiotropic Effects in Transgenic Tobacco Plants Expressing the Oryzacystatin I Gene. Horticulture Sciences 36:118-119. |
|
|
Habib H, Fazili KM (2007). Plant protease inhibitors: A defense strategy in plants. Biotechnology and Molecular Biology 2:68-85. |
|
|
Harsulkar AM, Giri AP, Patankar AG, Gupta VS, Sainani MN, Ranjekar PK, Deshpande VV (2002). Successive use of non-host plant proteinase inhibitors required for effective inhibition of Helicoverpa armigera gut proteinases and larval growth. Plant Physiology 121:497-506. |
|
|
Haruta M, Major IT, Christopher ME, Patton JJ, Constabel CP (2001). A Kunitz trypsin inhibitor gene family from trembling aspen (Populus tremuloides Michx.) : cloning, functional expression, and induction by wounding and herbivory. Plant Molecular Biology 46:347-359. |
|
|
Jamal F, Pandey PK, Singh D, Khan MY (2013). Serine protease inhibitors in plants: Nature's arsenal crafted for insect predators. Phytochemistry Review 12:1-34. |
|
|
Johnston KA, Lee ML, Gatehouse JA, Anstee JH (1991). The partial purification and characterisation of serine protease activity in midgut of larval Helicoverpa armigera. Insect Biochemistry 21:389-397. |
|
|
Jyothishwaran G, Kotresha D, Selvaraj T, Srideshikan S, Rajvanshi P, Jayabaskaran C (2007). A modified freeze thaw method for efficient transformation of Agrobacterium tumefaciens. Current Sciences 93:770-772. |
|
|
Kakade ML, Simons N, Liener IE (1969). An evaluation of natural and synthetic substrates for the antitryptic activity of soybean samples. Cereal Chemistry 46:518-522. |
|
|
Kansal R, Kumar M, Kuhar K, Gupta RN, Subrahmanyam B, Koundal KR, Gupta VK (2009). Purification and characterization of trypsin inhibitor from Cicerarietinum L. and its efficacy against Helicoverpa armigera. Brazilian Journal of Plant Physiology 20:313-322. |
|
|
Kim JY, Park SC, Hwang I (2009). Protease inhibitors from plants with antimicrobial activity. International Journal of Molecular Sciences 10:2860-2872. |
|
|
Kuwar SS, Pauchet Y, Vogel H, Heckel DG (2015). Adaptive regulation of digestive serine proteases in the larval midgut of Helicoverpa armigera in response to a plant protease inhibitor. Insect Biochemistry and Molecular Biology 59:18-29. |
|
|
Laluk K, Mengiste T (2011). The Arabidopsis extracellular unusual serine protease inhibitor functions in resistance to necrotrophic fungi and insect herbivory. The Plant Journal 68:480-494. |
|
|
Lichtenthaler HK (1987). Chlorophylls and carotenoids: pigments of photosynthetic bio-membranes. Methods in Enzymology 148:350-382. |
|
|
Lomate PR, Dewangan V, Mahajan NS, Yashwant K, Kulkarni A, Wang L, Saxena LS, Gupta VS, Giri AP (2018). Integrated transcriptomic and proteomic analyses suggest the participation of endogenous protease inhibitors in the regulation of protease gene expression in Helicoverpa armigera. Molecular and Cellular Proteomics 17:1324-1336. |
|
|
Manamohan M, Prakash MN, Chandra GS, Asokan R, Nagesha SN (2011). An improved protocol for rapid and efficient Agrobacterium mediated transformation of tomato (Solanumlycopersicum L.). Journal of Applied Horticulture 13:3-7. |
|
|
Munger A, Coenen K, Cantin L, Goulet C, Vaillancourt LP, Goulet MC, Tweddell R, Sainsbury F, Michaud D (2012). Beneficial 'unintended effects' of a cereal cystatin in transgenic lines of potato (Solanum tuberosum). BMC Plant Biology 12:1-12. |
|
|
Ntagkas N, Woltering E, Bouras S, De Vos RCH, Dieleman JA, Nicole CCS, Labrie C, Marcelis, LFM (2019). Light-induced vitamin C accumulation in tomato fruits is independent of carbohydrate availability. Plants 8:86. |
|
|
Papolu PK, Dutta TK, Tyagi N, Urwin PE, Lilley CJ, Rao U (2016). Expression of a cystatin transgene in eggplant provides resistance to root-knot nematode, Meloidogyne incognita. Frontiers in Plant Sciences 1122:1-9. |
|
|
Parde VD, Sharma HC, Kachole MS (2010). In vivo inhibition of Helicoverpa armigera gut pro-proteinase activation by non-host plant protease inhibitors. Journal of Insect Physiology 56:1315-1324. |
|
|
Pariani S, Contreras M, Rossi FR, Sander V, Corigliano MG, Simón F, Busi MV, Gomez-Casati DF, Pieckenstain FL, Duschak VG (2016). Characterization of a novel Kazal-type serine proteinase inhibitor of Arabidopsis thaliana. Biochimie 123:85-94. |
|
|
Quilis J, Meynard D, Vila L, Aviles FX, Guiderdoni E, Segundo BS (2007). A potato carboxypeptidase inhibitor gene provides pathogen resistance in transgenic rice. Plant Biotechnology Journal 5:537-553. |
|
|
Rustgi S, Boex-Fontvieille E, Reinbothe C, Wettstein DV, Reinbothe S (2017). Serpin1 and WSCP differentially regulate the activity of the cysteine protease RD21 during plant development in Arabidopsis thaliana. Proceedings of National Academy of Sciences. USA 114:2212-2217. |
|
|
Sambrook J, Fritish EF, Maniatis T (1989). Molecular cloning. A laboratory manual. 2nd Ed. Cold Spring Harbour Laboratory Press, USA. |
|
|
Santamaria ME, Diaz-Mendoza M, Diaz I, Martinez M (2014). Plant protein peptidase inhibitors: an evolutionary overview based on comparative genomics. BMC Genomics 15:812-826. |
|
|
Schluter U, Benchabane M, Munger A, Kiggundu A, Vorster J, Goulet MC, Cloutier C, Michaud D (2010). Recombinant protease inhibitors for herbivore pest control: a multitrophic perspective. Journal of Experimental Botany 61:4169-4183. |
|
|
Singleton VL, Rossi JA (1965). A colorimetry of total phenolics with phosphomolybdic -phosphotungstic acid reagents. American Journal of Enology and Viticulture 16:144-158. |
|
|
Somayaji P, Prasad DT, Shivanna MB (2014). Agrobacterium-mediated transformation for development of transgenic tomato with ySAMdc gene. Indian Journal of Biotech 13:19-25. |
|
|
Song SC, Yu JH, Hester PY, Kim KI (1985). Antibodies to the alpha subunit of insulin receptor from eggs of immunized hens. The Journal of Immunology 135:3354-3359. |
|
|
Srinivasan A, Giri AP, Harsulkar AM, Gatehouse JA, Gupta VS (2005). A Kunitz trypsin inhibitor from chickpea (Cicerarietinum L.) that exerts anti-metabolic effect on podborer (Helicoverpa armigera) larvae. Plant Molecular Biology 57:359-374. |
|
|
Stout MJ, Fidantsef AL, Duffey SS, Bostock RM (1999). Signal interactions in pathogen and insect attack: systemic plant-mediated interactions between pathogens and herbivores of the tomato (Lycopersicon esculentum). Physiology and Molecular Plant Pathology 54:115-130. |
|
|
Tamayo MC, Rufat M, Brava JM, Segundo BS (2000). Accumulation of a maize proteinase inhibitor in response to wounding and insect feeding, and characterization of its activity toward digestive proteinases of Spodoptera littoralis larvae. Planta 211:62-71. |
|
|
Tamhane VA, Chougule NP, Giri AP, Dixit AR, Sainani MN, Gupta VS (2005). In vivo and in vitro effect of Capsicum annum proteinase inhibitors on Helicoverpa armigera gut proteinases. Biochimica et Biophysica Acta 1722:156-167. |
|
|
Tang QB, Jiang JW, Yan YH, Van Loon JJA, Wang CZ (2006). Genetic analysis of larval host-plant preference in two sibling species of Helicoverpa. Entomologia Experimentalis et Applicata 118:221-228. |
|
|
Tanpure RS, Barbole RS, Dawkar VV, Waichal YA, Joshi RS, Giri AP, Gupta VS (2017). Improved tolerance against Helicoverpa armigera in transgenic tomato over-expressing multi-domain proteinase inhibitor gene from Capsicum annuum. Physiology and Molecular Plant pathology 23:597-604. |
|
|
Thomas JC, Wasmann CC, Echt C, Dunn RL, Bohnert HJ, Mccoy TJ (1994). Introduction and expression of an insect proteinase inhibitor in alfalfa Medicagosativa L. Plant Cell Reports, pp. 31-36. |
|
|
Tittonell P, Degrazia J, Chiesa A (2001). Effect of nitrogen fertilization and plant population during growth on lettuce (Lactucasativa L.) postharvest quality. Proceedings of the Fourth International Conference on Postharvest Science, Acta Horticulturae 553:67-68. |
|
|
Toor RK, Savage GP (2005). Antioxidant activity in different fractions of tomatoes. Food Research International 38:487-494. |
|
|
Turra D, Bellin D, Lorito M, Gebhardt C (2009). Genotype-dependent expression of specific members of potato protease inhibitor gene families in different tissues and in response to wounding and nematode infection. Journal of Plant Physiology 166:762-774. |
|
|
Vieira P, Wantoch S, Lilley CJ, Chitwood DJ, Atkinson HJ, Kamo K (2015). Expression of a cystatin transgene can confer resistance to root lesion nematodes in Lilium longiflorum cv. 'Nellie White'. Transgenic Research 24:421- 432. |
|
|
Wang Y, Ma Y, Zhou DS, Gao SX, Zhao XC, Tang QB, Wang CZ, Van Loon JJA (2017). Higher plasticity in feeding preference of a generalist than a specialist: Experiments with two closely related Helicoverpa species. Scientific Reports 7:1-12. |
|
|
War AR, Paulraj MG, Ahmad T, Buhroo AA, Hussain B, Ignacimuthu S, Sharma HC (2012). Mechanism of plant defense against insect herbivores. Plant Signaling and Behavior 7:1306- 1320. |
|
|
Wingate VPM, Franceschi VR, Ryan CA (2008). Tissue and Cellular Localization of Proteinase Inhibitors I and II in the Fruit of the Wild Tomato, Lycopersicon peruvianum (L.) Mill. Plant Physiology 97:490- 495. |
|
|
Winterer J, Bergelson J (2001). Diamondback moth compensatory consumption of protease inhibitor-transformed plants. Molecular Ecology 10:1069-1074. |
|
|
Wu J, Baldwin IT (2010). New Insights into Plant Responses to the Attack from Insect Herbivores. Annual Review of Genetics 44:1- 24. |
|
|
Wu Y, Llewellyn D, Mathews A, Elizabeth S (1997). Adaptation of Helicoverpa armigera (Lepidoptera: Noctuidae) to a proteinase inhibitor expressed in transgenic tobacco. Molecular Breeding, pp. 371-380. |
|
|
Yang F, Zhang Y, Huang Q, Yin G, Pennerman KK, Yu J, Liu Z, Li D, Guo A (2015). Analysis of key genes of jasmonic acid mediated signal pathway for defense against insect damages by comparative transcriptome sequencing. Scientific Reports 5:1-12. |
|
|
Zhu-Salzman K, Koiwa H, Salzman RA, Shade RE, Ahn JE (2003). Cowpea bruchid Callosobruchus maculatus uses a three-component strategy to overcome a plant defensive cysteine protease inhibitor. Insect Molecular Biology 12:135-145. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


