Full Length Research Paper

ABSTRACT
The expression of the UDP glucose: flavonoid-3-Oglucosyltransferase (UFGT), which encoded the last enzyme of the anthocyanin pathway, was under developmental control as well as affected by external stimuli such as ABA. Three fragments of the 1.38 kb upstream region of the UFGT gene from Litchi (Litchi chinesis Sonn.) were fused to the GUS-coding region, and the expression of these constructs was analyzed in onions. To characterize the cis-regulatory functions of the promoters for enzymes in anthocyanin biosynthesis, we examined onions carrying a series of nested UFGT promoter-β-glucuronidase (GUS) fusion for GUS activity by histochemical staining. The AE-box and ELI-box in anthocyanin biosynthesis of angiosperm were found in UFGT promoter. MRE which was MYB binding site involved in light responsiveness, control the transcription of genes in anthocyanin biosynthesis. The region of -910 to -344 in UFGT promoter showed high activities in the three parts. Although the expression characteristics were indistinguishable from those of the full-length promoter, we observed differences in UFGT promoter regulation for the different construct. The results suggested that region of 910 to 344 of UFGT promoter had multiple functions in the expression under the various developmental stages and stress conditions in litchi.
Key words: Flavonoid-3-Oglucosyltransferase (UFGT), promoter, litchi, transient expression.
INTRODUCTION
The litchi (Litchi chinesis Sonn.) is a tropical and subtropical fruit tree native to southern China, Taiwan, Bangladesh and Southeast Asia, and now cultivated in many parts of the world. The litchi has a history and cultivation going back as far as 2000 BC according to records in China. Cultivation began in the area of southern China, Malaysia and Vietnam. Wild trees still grow in parts of southern China and on Hainan Island. Litchi is extensively grown in China, and also elsewhere in Brazil, South-East Asia, India, Pakistan, Bangladesh, southern Japan and more recently in California, Jamaica and elsewhere in the Caribbean, Hawaii, Texas, Florida, the wetter areas of eastern Australia and sub-tropical regions of South Africa, Israel and also in the states of Sinaloa and San Luis Potosíin Mexico. A wide range of cultivars is available, with early and late maturing forms suited to warmer and cooler climates, respectively. They are also grown as an ornamental tree as well as for their fruit. Litchi was commonly sold fresh in Asian markets, and in recent years, also widely in supermarkets worldwide. The red rind turns dark brown when the fruit is refrigerated, but the taste is not affected. It is also sold canned year-round. Anthocyanin biosynthesis had been characterized in flowers of petunia, snapdragon, pericap of litchi, skins of grape and in kernels of maize, and the biosynthetic pathway is now one of the best known pathway in plants (Holton and Cornish, 1995). Anthocyanin was the predominant pigments of grape skins as well as other blue, red, or black fruits and flowers, and was biosynthesized through a flavonoid pathway. Regulation of the anthocyanin pathway had been shown to occur mainly at the transcriptional level in pigmented organs. Two classes of genes were required for anthocyanin biosynthesis. The structural genes encoded the enzymes that directly participate in the formation and storage of anthocyanins and other flavonoids. The UGFT enzyme of plants was the final gene in the anthocyanin pathway. The transfer of the glucosyl moiety from UDP-glucose to the 3-hydroxyl group of anthocyanidins by UFGT was shown to be the key for anthocyanidin stability and water solubility (Yoshihara et al., 2005). In grapevines, the main control point for anthocyanin quantitative variation was downstream in the pathway at theUFGT level, in agreement with the early observations (Boss et al., 1996a, b, c). In the grape berry, anthocyanin biosynthesis pathway, some cDNAs encoding structural genes had been cloned in Vitis vinifera (Sparvoli et al., 1994). It was showed that only the expression of the gene coding for UFGT was consistently associated with the berry color, depending on developmental stage and cultivar (Boss et al., 1996a, b, c). Expression analysis of UFGT genes in white and red-skinned cultivars revealed that the UFGT gene was present, but not expressed in the white cultivars (Boss et al., 1996a, b, c; Kobayashi et al., 2001). The UFGT sequences were identical in white and red-skinned sports including the promoter region, suggesting that the phenotypic change from white to red could be the result of a mutation in a regulatory gene controlling the expression of UFGT (Kobayashi et al., 2002). Commercial litchi cultivation had focused on fruit crops that develop red color within a month time on the tree. Therefore, this fruit could provide a model for investigating regulation of anthocyanin synthesis during fruit maturation. Here, we reported the isolation of the promoter of UFGT gene in the anthocyanin biosynthetic pathway. Expression of the GUS gene was examined in a series of nested UFGT promoter. The evidence presented revealed that GUS activity was the highest in one element of about 360 bp upstream of the transcription start site.
MATERIALS AND METHODS
Plant material and bacterial strains
Litchi (L. chinesis Sonn.) cv. Nuomici was grown at College of Horticulture, South China Agricultural University in Guangzhou, China. Fresh leaves and fruit samples were taken to the laboratory and immediately frozen in liquid nitrogen and stored at -80°C. Escherichia coli strain DH5a was cultivated in LB medium for vector constructs and DNA manipulation. Agrobacterium tumefaciens strain EHA105 was cultivated in LB medium for onions transformation. White skin onion cultivar was used for transformation.
PCR cloning of the UFGT promoter region
Total genomic DNA was isolated from fresh leaves samples by CTAB method (Stewart and Via, 1993). The promoter region was cloned with high-efficiency thermal asymmetric interlaced PCR method described previously (Liu and Chen, 2007). The corresponding produce was subjected to pre-amplification reactions of TAIL-PCR with inner gene-specific primer (GSP1) (5'-TTC CTT CTT TGT TCT CTC AGC AGA CC-3') and the primary or secondary TAIL-PCRs with outer gene-specific primer (GSP2) (5'-ATT GTG TGTAGC AAC AGC AGG GC-3'). Major bands were isolated from using TaKaRa Agarose Gel DNA Purification kit (TaKaRa, Japan), and the isolated fragments were cloned into a pMD-19T vector (TaKaRa, Japan). The inserted sequence in recombinant plasmid DNA was analyzed using a BigDyeTM Terminator Cycle Sequencing Ready Reaction Kit (Perkin Elmer, USA) on an ABI PRISMTM 377 DNA Sequencer. In order to find out motifs regulation UFGT gene expression, functional motifs in UFGT gene promoter were predicted by PlantCARE (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/).
Construction of vectors
With the genomic as a template, a fragment upstream of the translational start codon of UFGT gene and its 5' deletion derivatives were generated by PCR with five primers: (Pf1:5'-CAAGCTTGGCCGCCCTGTTGGTTAAATAGTAT-3’; Pr1:5'-CGGATCCAGTGGTGGTGTTGTGGTGGTGGATA-3'; Pf2:5'-CAAGCTTTTAGTAAGGGCTGGATTAAGTGAAT-3'; Pr2:5'-CGGATCGAGGTTCTTAGTGCGTATCGGTCTTCG -3';Pf3:5'- CAAGCTTGACTGATATTTCGGTTTGAACTGCA-3'; Pr3: 5'-CGGATCCCCCCACGCTCACTACGAGACTACCC-3';), the introduced HindIII sites in the forward primers and the introduced BamHI site in the reverse primers. The amplified fragments were respectively inserted into the plasmid pBI121 (Clontech) as a HindIII-BamHI fragment at the corresponding restriction sites in place of the cauliflower mosaic virus (CaMV) 35S promoter region, resulting in a series of pBI-pUFGT::GUS vectors.
Onions transformation
Individual binary vectors, including pBI121 and a series of pBI-pUFGT::GUS were introduced into A. tumefaciens EHA105 by the freeze-thaw method (Walkerpeach and Velten, 1994).
The fresh onion outside 3 to 4 layers scales was removed , the bulb immersed in 75% ethanol with 10 min, washed three times with sterile water. Onions were cut with sterile scalpel bulbs, corms with fresh and hypertrophy of the internal scales including epidermal (concave) were cut an area of 1-2 cm of the small box. After the pre-incubation of onion tissue, they were placed in MS liquid medium re-suspended in broth for 20 min, slightly drained broth, spread on MS solid medium to photoperiod 16/8 h at 25°C, co-culture. After 16-24 h, the onion small piece of skin was removed with a clean wash liquid MS medium slightly shaken to remove the attached Agrobacterium. Transformed onion tissues were measured with histochemical staining and GUS activity.
Fluorometric quantification of GUS activity and histochemical staining
Plant tissues were ground into a fine powder using liquid nitrogen with a mortar and pestle, and suspended in GUS extraction buffer (50 mM sodium phosphate, pH 7.0; 0.1% Triton X-100; 10 mM 2- mercaptoethanol; 10 mM 1,2-diaminocyclohexane-N,N,N,N-tetraacetic acid and 0.1% sodium lauryl sarcosine). The supernatant was collected after centrifugation at 12 000× g for 10 min at 4°C. Fluorometric quantification of GUS activity was performed using 4-methylumbelliferyl-b-D-glucuronide substrate (Jefferson et al., 1987). The content of total proteins was determined using the Bradford (1976) method. The GUS activity was expressed as pmol of 4-methylumbelliferone per mg protein per min. Histochemical localization of GUS activity was performed as follows: samples were fixed with 0.5% paraformaldehyde in 0.1 M sodium phosphate (pH 7.0) for 30 min, then various tissues of transgenic tobacco samples were incubated in 5-bromo-4-chloro-3-indolyl-D-glucuronic acid (X-gluc) solution at 37°C from 3 h overnight until the blue staining reached sufficient intensity (Grotewold et al., 1994). Photosynthetic tissues were cleared of chlorophyll by passaging through a 70-100% ethanol series. Photography was performed with a camera (Nikon 8700, Japan).
RESULTS AND DISCUSSION
Isolation and characterization of litchi UFGT promoter
With the genomic as a template, a 1487 bp fragment upstream of the translational start codon of UFGT gene was gotten (Figure 1). The three expression of vector were named P(-1487/-18), as the full-length promoter construct in this study, P1(-1487/-857), P2(-910/-344) and P3(-477/-18) (Figure 2).

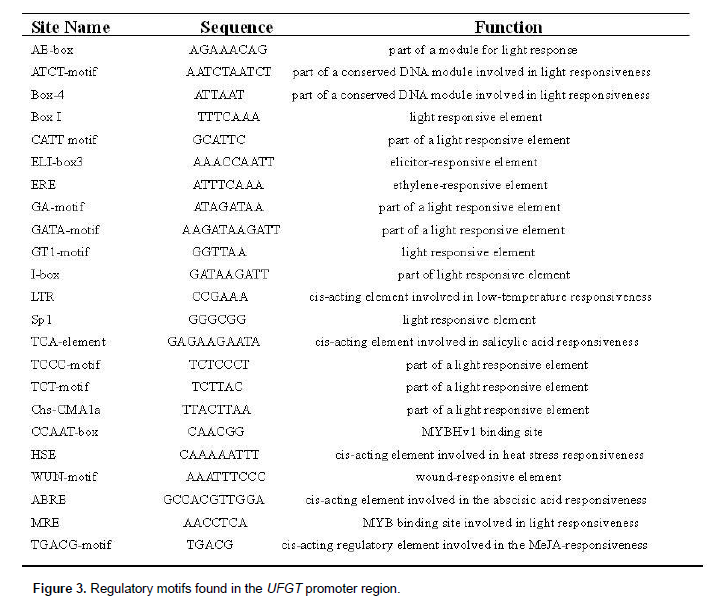
The transcriptional start sites of UFGT were determined and located 85 bp upstream from the ATG codon. The AE-box, and ELI-box in anthocyanin biosynthesis of angiosperm were found in UFGT promoter. MRE which was MYB binding site involved in light responsiveness, control the transcription of genes in anthocyanin biosynthesis (Grotewold et al., 1994; Jin et al., 2000; Moyano et al., 1996; Sablowski et al., 1994; Sainz et al., 1997; Sugimoto et al., 2000; Tamagnone et al., 1998; Yang, 2001). The UFGT gene promoter contained TCA elements and a HSE homologous sequence, UFGT promoter also contained WUN-motif that was wound-responsive element (Figure 3).
Expression pattern of the UFGT promoter in onions (Allium cepa Linn.)
In order to identify promoter regions that confer the spatial and developmental expression of litchi genes, the 1.48 kbp promoter fragment and three promoter deletions of UFGT were fused to a GUS reporter gene (Figure 4). All constructs were transferred into onions plants by Agrobacterium-mediated transformation system. Ten to 15 independent onions transformants were obtained for each construct. Figure 5 showed the GUS activities in onions transformants with the UFGT promoter-GUS construct. The GUS expression levels of the the P1promoter-GUS construct (-1487/-857) and the P3promoter (-477/-18) fragments were very low in onions plants. The highest level of GUS activity was detected in transformants carrying UFGT full length (-1,487 bp from the transcriptional start site). We then analyzed the histochemical localization for every individuals obtained from each UFGT promoter-GUS construct. In onions tissue, the expression levels of the P1promoter (-1487/-857) and the P3 promoter (-477/-18) deletions were very low (Figure 5) and CK no histochemical localization was detected. These results suggest that the region–910 to–344 of the UFGT promoter, in which reside HSE, ABRE, MRE and TCA-element (Table 1) responsible for the strong heat, light, and ABA expression in the transgenic onions. The UFGT promoter construct, P2 produced significant GUS activity in response to bending in onions (Figure 5), whereas other constructs did not show high activity. This indicates that the UFGT promoter region between -910 and -344 bp also contains possible bending stress responsive elements in litchi.
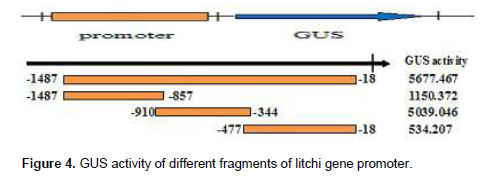

Functional motifs of the UFGT gene promoter was predicted by PlantCARE. Promoter deletion analysis of UFGT showed that P2 (-910/-344) promoter-driven GUS activity was detected in response to ABA stress in onions. The expression of UFGT is strongly associated with the anthocyanin accumulation in “Feizixiao” litchi by ABA treatment. The expression of UFGT and anthocyanin were affected by bagging in “Feizixiao”litchi. The induction of UFGT-910 activity under bending stress suggested that the activation of promoter under bending stress might be one of important regulatory mechanisms to response to the stress in onions. The UFGT promoter –910 to –344 contained ABRE, MRE and TCA-element, also have multiple functions in controlling the expression of UFGT under various conditions in litchi. The results suggested that the region of -910 and -344 in the UFGT promoter contains a putative positive regulatory element in the expression of UFGT gene. R2R3-MYB family has important roles in the expression of genes encoding the enzymes in the anthocyanin metabolism via interaction of AC elements (Stracke et al., 2001). Previous study showed that the ectopic expression of PtMYB4 in transgenic tobacco enhanced the transcription of several genes for lignin biosynthetic enzymes (Patzlaff et al., 2003). Recently, several MYBs expressed in differentiating xylem has been isolated from Picea glauca; PgMYB1, PgMYB2, PgMYB3, PgMYB4 and PgMYB8 from Picea glauca (Moench) Voss (Bedon et al., 2007). Further studies will be needed to identify conclusively that the transcription factors control the UFGT gene transcription in litchi. In conclusion, our study showed that UFGT promoter from litchi directs the GUS reporter gene expression in onion. Deletion analysis revealed that the promoter region between -910 and -344 was necessary for its function. This study provides a candidate of cis-regulatory elements that control anthocyanin biosynthsis gene expression from litchi and would contribute to the molecular mechanism of litchi coloring.
CONFLICT OF INTEREST
The authors have not declared any conflict of interest.
ACKNOWLEDGEMENTS
The authors thank Dr Wang and Miao of the College of Horticulture, South China Agricultural University. This work was supported by the “China Litchi Industry Technology Research System” (nycytx-32-01) and “National Natural Science Fund of China” (30971985, 31471850?948 (2011-G13).
REFERENCES
|
Bedon F, Grima-Pettenati J (2007). Mackay, J., Conifer R2R3-MYB transcription factors: sequence analyses and gene expression in wood-forming tissues of white spruce (Picea glauca), BMC Plant Biol. 7:17. Crossref |
|||||||
|
Boss PK, Davies C, Robinson SP (1996a). Analysis and the expression of anthocyanin pathway genes in developing Vitis vinifera L. cv Shiraz grape berries and the implications for pathway regulation, Plant Physiol. 111:1059-1066. Pubmed |
|||||||
|
Boss PK, Davies C, Robinson SP (1996b). Anthocyanin composition and anthocyanin pathway gene expression in grapevine sports differing in berry skin colour. Aust. J. Grape Wine Res. 2:163–170. Crossref |
|||||||
|
Boss PK, Davies C, Robinson SP (1996c). Expression of anthocyanin biosynthesis pathway genes in red and white grapes, Plant Mol. Biol. 32:565–569. Crossref |
|||||||
|
Bradford MM (1976). A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding, Anal Biochem. 72:248-254. Crossref |
|||||||
|
Grotewold E, Drummond BJ, Bowen B, Peterson T (1994). The mybhomologous P gene controls phlobaphene pigmentation in maize floral organs by directly activating a flavonid biosynthetic gene subset, Cell. 76:543-553. Crossref |
|||||||
|
Holton TA, Cornish EC (1995). Genetics and biochemistry of anthocyanin biosynthesis, Plant Cell. 7:1071-1083. Crossref |
|||||||
|
Jefferson RA, Kavanagh TA, Bevan MW (1987). GUS fusions: b-glucuronidase as a sensitive and versatile gene fusion marker in higher plants, EMBO J. 6:3901-3907. Pubmed |
|||||||
|
|||||||
|
Kobayashi S, Ishimaru M, Ding CK, Yakushiji H, Goto N (2001). Comparison of UDP-glucose: flavonoid 3-O-glucosyltransferase (UFGT) gene sequences between white grapes (Vitis vinifera) and their sports with red skin, Plant Sci. 160:543-550. Crossref |
|||||||
|
Kobayashi S, Ishimaru M, Hiraoka K, Honda C (2002). Mybrelated genes of the Kyoho grape (Vitis labruscana) regulate anthocyanin biosynthesis, Planta. 215:924–933. Crossref |
|||||||
|
Liu YG, Chen YL (2007). High-efficiency thermal asymmetric interlaced PCR for amplification of unknown flanking sequences, BioTechniques, 431:649-656. Crossref |
|||||||
|
Moyano E, Martinez-Garcia JF, Martin C (1996). Apparent redundancy in myb gene function provides gearing for the control of flavonoid biosynthesis in antirrhinum flowers, Plant Cell. 8:1519–1532. Crossref |
|||||||
|
Patzlaff A, Newman LJ, Dubos C, Whetten RW, Smith C, McInnis S, Bevan MW, Sederoff RR, Campbell MM (2003). Characterisation of PtMYB1, an R2R3-MYB from pine xylem, Plant Mol Biol. 53:597–608. Crossref |
|||||||
|
Sablowski RW, Moyano E, Culianez-Macia FA, Schuch W, Martin C, Bevan M (1994). A flower-specific Myb protein activates transcription of phenylpropanoid biosynthetic genes, EMBO J. 13:128–137. Pubmed |
|||||||
|
Sainz MB, Grotewold E, Chandler VL (1997). Evidence for direct activation of an anthocyanin promoter by the maize C1 protein and comparison of DNA binding by related Myb domain proteins, Plant Cell. 9:611–625. Crossref |
|||||||
|
Sparvoli F, Martin C, Scienza A, Gavazzi G, Tonelli C (1994). Cloning and molecular analysis of structural genes involved in flavonoid and stilbene biosynthesis in grape (Vitis vinifera L.), Plant Mol. Biol. 24:743-755. Crossref |
|||||||
| Stewart CN, Via LE (1993). A rapid CTAB DNA isolation technique useful for RAPD fingerprinting and other PCR applications, Biotechniques, 5:748-750. | |||||||
|
Stracke R, Werber M, Weisshaar B (2001). The R2R3-MYB gene family in Arabidopsis thaliana, Curr. Opin. Plant Biol. 4:447–456. Crossref |
|||||||
|
Sugimoto K, Takeda S, Hirochika H (2000). MYB-related transcription factor NtMYB2 induced by wounding and elicitors is a regulator of the tobacco retrotransposon Tto1 and defenserelated genes, Plant Cell. 12:2511–2528. Crossref |
|||||||
|
Tamagnone L, Merida A, Parr A, Mackay S, Culianez-Macia FA, Roberts K, Martin C (1998). The AmMYB308 and AmMYB330 transcription factors from antirrhinum regulate phenylpropanoid and lignin biosynthesis in transgenic tobacco, Plant Cell. 10:135–154. Crossref |
|||||||
|
Walkerpeach CR, Velten J (1994). Agrobacterium-mediated gene transfer to plant cells: co-integrate and binary vector systems, In: Gelvin SB, Schilperoort RA, eds. Plant molecular biology manual. Dordrecht: Kluwer, pp. 1-19. Crossref |
|||||||
|
Yang S, Sweetman JP, Amirsadeghi S, Barghchi M, Huttly AK, Chung WI, Twell D (2001). Novel anther-specific myb genes from tobacco as putative regulators of phenylalanine ammonialyase expression, Plant Physiol. 126:1738–1753. Crossref |
|||||||
|
Yoshihara N, Imayama T, Fukuchi-Mizutani M, Okuhara H, Tanaka Y, Yabuya T (2005). cDNA cloning and characterization of UDP-glucose: anthocyanidin 3-O-glucosyltransferase in Iris hollandica. Plant Sci. 169:496-501. Crossref |
|||||||
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


