Full Length Research Paper

ABSTRACT
Morphological characteristics of the fruit, stem and leaf tissues of four species of Citrullus (L.) Schrad. were examined using standard histological methods. Plant materials included the cultivated watermelon (Citrullus lanatus (Thunb.) Matsum. & Nakai) and three of its related species (Citrullus colocynthis (L.) Schrad., Citrullus ecirrhosus Cogn. and Citrullus rehmii de Winter). Variation among the species was observed in the thickness of the subepidermal layer(s) of the fruit, this layer being thicker in fruits of C. ecirrhosus (~ 200 uM) than in fruits of C. lanatus (~100 uM), C. colocynthis (~100 uM) or C. rehmii.(~100 µM). Variation was also observed in the extent and organization of the subtending sclerenchymatous cells which were up to 10 layers thick in fruits of C. colocynthis, 1 or 2 cell layers thick in the fruits of C. lanatus., and an intermediate number of layers (3 to 6) in fruits of C. ecirrhosus and C. rehmii. A greater degree of lignifications was observed in the stem tissue of C. colocynthis and C. ecirrhosus, in comparison with that of C. lanatus and C. rehmii. Leaf thickness (~250 µM) was similar among three of the four species examined, but was reduced in C. rehmii (~150 µM). Further study is required to assess the possible contribution of these, and other, morphological attributes to drought tolerance in members the genus Citrullus.
Key words: Watermelon, Citrullus lantatus, Citrullus colocynthis, Citrullus rehmii, Citrullus ecirrhosus, histology, anatomy, structure, drought tolerance, caudex, pollen viability.
INTRODUCTION
The sweet-fleshed form of the cultivated watermelon (Citrullus lanatus) is an important crop in the USA and internationally (Levi et al., 2012). US PRODUCTION of watermelon ranked 5th globally at ~50 x 103 hectares with an estimated value of more than a half-billion dollars in 2012(http://www.agmrc.org/commodities__products/vegetables/watermelon/). In 2014, watermelon production in China and the USA exceeded 68.8 x 106 and 1.69 x 106 m/t, respectively (Food and Agriculture Organization of the United Nations, 2014). These data do not include the production of non-sweet citrons, egusi types and the bitter apple. In order to maintain (and improve) the current high levels of production of this crop, breeding objectives have typically included disease and insect tolerance/resistance, (Duke, 1978). in addition to fruit quality attributes. However, recent concerns associated with the increasing costs of irrigation (and water scarcity), and anticipated changes to the environment (including increased risk of environmental stress) have provided an additional area of study for the improvement of watermelon and other crops -drought tolerance (Condon et al., 2004; Liuigi et al., 2008). Fortunately, drought tolerance is a characteristic associated with members of this genus that are native to southern and western Africa (Meeuse, 1961, 1962; Jeffrey, 1975; Russell, 1985; Singh, 1990, Chomicki and Renner, 2014).
The Citrullus genepool is relatively small in comparison with that of other major vegetable crops (Whitaker and Davis, 1962; Bates and Robinson, 1995). Four diploid (2n=22) species are generally recognized; Citrullus lanatus (Thunb.) Matsum. & Nakai - the cultivated watermelon, Citrullus colocynthis (L.) Schrad., Citrullus ecirrhosus Cogn. and Citrullus rehmii De Winter. Recent taxonomic changes suggest that additional species be recognized (Chomicki and Renner, 2014). C. ecirrhosus is a desert perennial (Jeffrey, 1975; Russell, 1985). Fruits mature in February to March in the Namib where the leaves and stems die back each year and regrow from a caudex. This is a hardy species surviving on minimal ground water and morning fogs where it serves as an important source of water for desert fauna. In contrast to other members of the genus, C. ecirrhosus lacks tendrils. Its distribution appears to be restricted to Namibia and the Richtersveld of the Northern Cape (South Africa). C. colocynthis (a perennial in warmer climates) is also highly xerophytic and thrives on sandy loams, subdesert soils and along sandy seacoasts (Jeffrey, 1961; Duke, 1983). This species is distributed from northwestern Africa and the Mediterranean eastwards to Pakistan, India and Afghanistan (Sarafis, 1999; Renner and Pandey, 2013). In addition to being a source of seed-extractable oil, it is also used for medicinal purposes and has been cultivated in the Mediterranean region for centuries (Hussain et al., 2014). The annual C. rehmii is a more recently described species (De Winter, 1990) endemic to the desert regions of western and central Nambia. Possessing a central taproot, it inhabits gravelly to sandy-gravelly flats (De Winter 1990). C. lanatus, also an annual, is the most morphologically variable member of the group and has been cultivated in Central Africa for at least 5000 years (Whitaker and Davis, 1962). Though perhaps not generally considered to be as drought tolerant as its desert-dwelling related species, specific forms of C. lanatus [egusi types also referred to as C. lanatus var. (or subsp.) mucospermus] are common in the extremely arid regions of Northwestern Africa (National Research Council, 2006).
All species in this genus are regarded as xerophytes (Arnold and De Wet, 1993). However, relatively little information is available in the scientific literature regarding the mechanisms that affect their drought tolerance. Akashi et al. (2001) attributed elevated levels of citrulline in leaves to providing drought tolerance in Citrullus spp. Akashi et el. (2004) later demonstrated that drought-resistance in wild watermelon was correlated with the expression CLMT2, a gene sharing significant homology with a type-2 metallothionein (MT). Liu et al. (2008) noted that thicker leaves and stronger roots were associated with drought tolerance in mature C. lanatus plants, and that drought tolerance of C. lanatus geno-types at the seedling stage was not always correlated with tolerance at a more advanced stage of plant maturity. Zhang et al. (2011) screened 820 genebank accessions of C. lanatus for drought tolerance and assigned these to four groups, including tolerant, intermediate tolerant, moderately sensitive, and sensitive, respectively. The most drought-tolerant Citrullus germplasm, including 13 Citrullus lanatus var. lanatus and 12 C. lanatus var. citroides accessions, originated from Africa. It was suggested that these genetic materials could be used for rootstock breeding or for developing drought-tolerant watermelon cultivars, though no specific attribute conferring drought tolerance to the tolerant genotypes was identified.
The surface and other anatomical characteristics of stems, leaves and fruits are important factors capable of adaptations conferring drought tolerance (Baker and Procopiou, 2000). Thus, this study was undertaken to anatomically characterize stem, leaf and fruit pericarp tissue of the Citrullus spp. to attempt to identify unique anatomical features associated with individual taxa, and perhaps with drought tolerance.
MATERIALS AND METHODS
Plant materials for this study were obtained from the USDA/ARS genebank in Griffin, GA. Seeds of C. lanatus cv. Sugar Baby (PI 665007) and C. colocynthis (PI 652254) were treated with fungicide (Arasan) and planted in a commercial potting mix (Metro-Mix 360) in 2.5” x 2.5” x 3.5” peat pots. Previous efforts to germinate seed of C. ecirrhosus and C. rehmii in soil following the previously described procedure, proved unsuccessful. Seeds of C. ecirrhosus - (GRIF 15029) and C. rehmii GRIF 16946) were surface sterilized in 10% bleach and rinsed with tap water. The seed coats were then cracked with a pair of pliers and the seeds subsequently germinated in a germination chamber in darkness at a constant 30°C. Germinated seeds were transferred to peat pots containing the soil mix noted previously. When plants (3/spp.) reached the four to five-leaf stage, they were transplanted into plastic pots (~24 cm diameter x 18 cm depth) containing a 1:1:1 (v/v) mixture of the previously mentioned potting mix, pearlite and course sand. All plants received periodic fertilization with macro and micronutrients. Mature plants were established and maintained in the greenhouse from January 2012 through June 2013 without supplemental lighting at an average temperatures of 21 (days) and 16°C (nights). Fruit (1/plant; 3/spp.) were produced by manually pollinating (sib-mating) flowers. Efforts to establish plants of C. rehmii and C. ecirrhosus in the field in Griffin, GA were unsuccessful. Hence, only greenhouse-grown plants and fruits were included in the analyses.
Fixing, embedding and thin-sectioning of plant tissues
Procedures for the fixing, paraffin embedding and staining of tissues generally followed the procedures of Sass (1958) as described here. Sections of mature fruit pericarp (~1.5 cm in depth), mature leaf tissue and stem tissue (from the terminal 10 cm of rapidly growing vines) were harvested and placed in glass vials. FAA (50% EtOH, 5% acetic acid, 10% (37-40%) formaldehyde, and 35% ddH2O, v/v) was added to completely cover the tissue samples which were then vacuum infiltrated for 15 min. The vials were subsequently drained, fresh FAA added, and the tissues allowed to fix for 48 h at RT. Tissues were then transferred to 70% EtOH for a maximum of 2 weeks before dehydration and embedding.
Tissues were dehydrated in an alcohol series (60 min in each) of 70, 80, 95 and 100%, and then cleared in ethylene:xylene (30 min each in 3:1 ethanol:xylene, 1:1 ethanol:xylene, 1:3 ethanol:xylene and 100% xylene) (v/v). Fully dehydrated tissues were placed in molten Tissue Prep II (Fisher Scientific) at 60°C. The Tissue Prep was replaced 3 times at 12 h intervals followed by a final immersion for 48 h. Thin-sections (10 µ) were cut on a Ventana model 100 rotary microtome (Ventana Medical Systems, Tucson, AZ) and placed on glass slides.
Staining of plant tissues and microscopy
Paraffin was removed and tissues prepared for staining by successive immersions of 2 min each in 100% xylene, 100% EtOH, 90% EtOH, 70% EtOH, 50% EtOH, 30% EtOH and distilled H2O. Rehydrated tissues were stained in an acquoeous solution (1%, w/v) of safranin (Sigma Chemical) for 8 h. Excess safranin was removed by immersion in ddH2O (several water changes) and then dehydrated in an alcohol series (5 min each) of 30, 50, 70 and 95% EtOH. Tissues were counter-stained for ~5 s in 0.1% fast green (in 95% EtOH). Stained tissues were briefly immersed in 100% EtOH (3X) - 5 min each, 100% xylene (3X) - 5 min each, drained and the cover slip mounted with Permount (Fisher Scientific). Pollen grains were collected at ~8AM and immediately stained with 3.3. diaminobenzidine (Sigma Cat. # D-4168) as described by Dafni et al. (2005).
Microscopy was accomplished on an Olympus Model CX31 bright field microscope equipped with a digital camera.
RESULTS AND DISCUSSION
Cucurbitaceaous fruits possess a rind which serves to confine the fleshy tissues, and that is typified by the presence of a uniserate epidermis with a cuticle (con-taining stomata), and a subepidermal tissue that varies in width among species. This subepidernal tissue may consist of parenchyma (colored or colorless) and some-time collenchymatic (non-lignified) cells. At a greater depth, a layer of sclerids (sclerenchyma) occurs beneath which parenchmous tissue extends to the center of the fruits (Esau, 1977).
Variation among Citrullus species was observed in the thickness of the subepidermal tissue layer (Figure 1A - 1D) being less than 100 µM thick in fruit of C. ecirrhosus and approximately 175 to 225 µM in fruit of C. colocynthis and C. rehmii. Fruit of C. lanatus were intermediate to these values for this characteristic (125 to 150 µM). Variation among species was also seen in the extent and degree of organization of the subtending sclerenchymous cells being most extensive (up to 10 cell layers thick) in C. colocynthis, only a single or perhaps two cell layers thick in C. lanatus, and an intermediate layer of cells (3 to 6) in C. rehmii and C. ecirrhosus. Observations of this sclerenchymous layer suggested that is more highly organized in the fruits of C. rehmii than in the other species examined (Figure 1A to D).
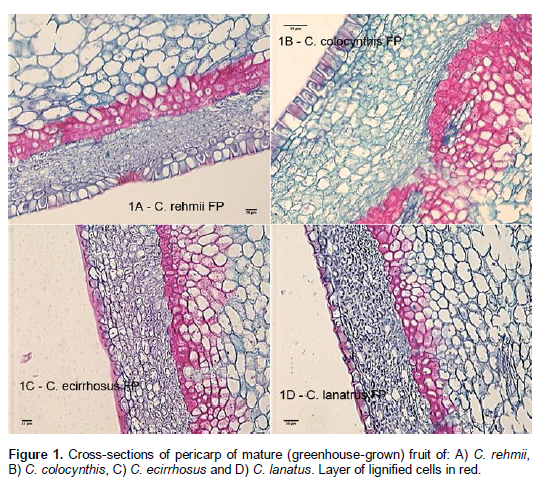
The occurrence of a greater degree of lignification within the exocarp of fruits of C. colocynthis is in keeping with the noticeably firmer fruit of this species when compared with C. ecirrhosus, C. lanatus or C. rehmii. Fruits of C. colocynthis were globose, 6 to 10 cm in diameter with green striping and intensely bitter flesh. In contrast to the other members of this genus that were examined in this study, the fruits of C. rehmii had neither a smooth surface nor were noticeably firm. The fruits of C. rehmii had a semi-flexible rind with a rugose-surface resembling, in that respect, fruit of certain Cucumis spp. The surface of mature C. rehmii fruits was mottled in appearance being generally dark green to tan in color with salmon-pink to orange-pink splotches as described by De Winter (1990). The fruit flesh of C. rehmii is known to be bitter and contain cucurbitacin E, B and I (Enslin and Rehm, 1958). Mature fruit of C. lanatus and C. ecirrhosus remained green at maturity, whereas mature fruit of C. colocyhthis developed a yellowish overcast as noted by Sarafis (1999).
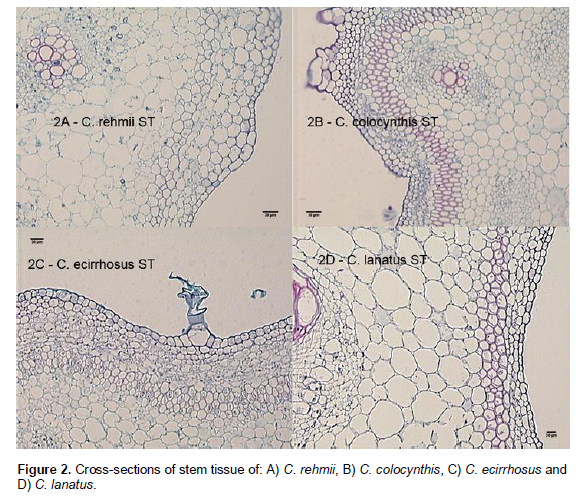
Cross-sections of stem (vine) tissue revealed a somewhat greater degree of lignification in the sections of C. colocynthis and C. ecirrhosus as illustrated in Figure 2B and C, and a reduced and more organized cell layer in C. colocynthis, as compared to other species. In contrast to C. lanatus and C. rehmii, both C. colocynthis and C. ecirrhosus develop swollen stems (caudices) from which multiple shoots arise. A greater degree of caudex development was observed in C. ecirrhosus as compared to C. colocynthis. Caudices averaged 2.5 cm in diameter in C. ecirrhosus and 1 cm or less in C. colocynthis - over the time span of these experiments. Histological examination of the caudices of these two species was not undertaken. A determination of the extent to which, if any, a caudex contributes to drought tolerance may prove useful.
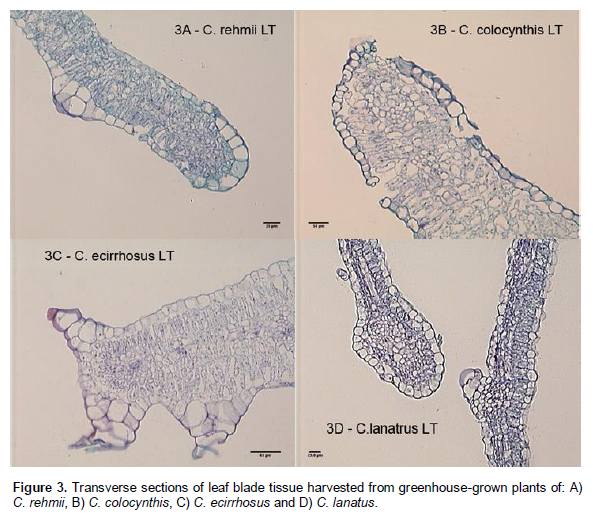
Adaptation to arid conditions is partially determined by the ability of the plant to regulate water loss from the aerial tissues (Ali and Grace, 1986; Baker and Procopiou, 2000; Benesova et al., 2012). Increased leaf (Karaba et al., 2007) and cuticle thickness (Martin and Juniper, 1974) have been shown to be associated with drought tolerance and leaf thickness has been suggested as a characteristic to be used in the selection of drought tolerant breeding lines (Hameed et al., 2002, 2012). We found leaf thickness to be variable among the four Citrullus species: C. lanatus - 80 to 100 µM; C. colcynthis - 200 to 250 µM; C. rehmii 100 to 150 µM; C. ecirrhosus - 250 to 350 µM (Figure 3A to D). General leaf morphology of C. rehmii was most similar to C. lanatus (broadly cordate and lobed) though its leaf size is reduced as compared to C. lanatus. The leaf surface area of C. colocynthis is also reduced as compared to both C. lanatus and C. rehmii, its leaves being triangular in shape and very deeply lobed. The leaves of C. ecirrhosus have the most distinctive morphology of the 4 species. The lamina of its leaves is curved over the mid-rib and the lateral veins so that when viewed from above the top surface is only visible in the vein regions and the leaves have a greenish white appearance due to the lower epidermis being reflected up as the upper surface of the leaf. This curving of the lamina is less evident when plants are grown in the greenhouse. The lower epidermis is covered with warts and hairs which account for the whitening effect. Both lower and upper epidermis contain similar amounts of stomata (Sarafis, 1999).
The extent to which the drought tolerance inherent in the Citrullus genepool can be characterized and harnessed to substantially improve the drought tolerance of the crop, remains to be determined. Drought tolerance is a complex trait involving multiple mechanisms that typically co-exist and that together contribute to whole-plant drought resistance (Xiong et al., 2012). The potential contribution of specific anatomical, morpholo-gical, physiological and other unique characteristics present in this genus towards improving drought tolerance in watermelon (and perhaps other crops as well) requires additional study. C. rehmii was successfully hybridized with C. lanatus by Rehm and Neethling (Unpublished Progress Report, 1967 as cited by De Winter, 1990). C. rehmii was also successfully hybridized with C. lanatus, C. ecirrhosus and C. colocynthis and the F1 hybrids found to be fertile. The F2 progeny of the crosses C. lanatus x C. colocynthis and C. lanatus x C. ecirrhosus showed a high degree of sterility whereas progeny of the cross C. colocynthis x C. ecirrhosus proved to be fertile (R.P. Elis as noted by De Winter, 1990). Navot and Zamir (1987) suggested the use of C. ecirrhosus as a source of drought tolerance genes for watermelon improvement and successfully hybridized C. ecirrhosus and C. lanatus (Navot et al., 1990). This has been viewed as demonstrating the potential for breeding C. lanatus containing genes from C. ecirrhosus (Sarafis, 1999).
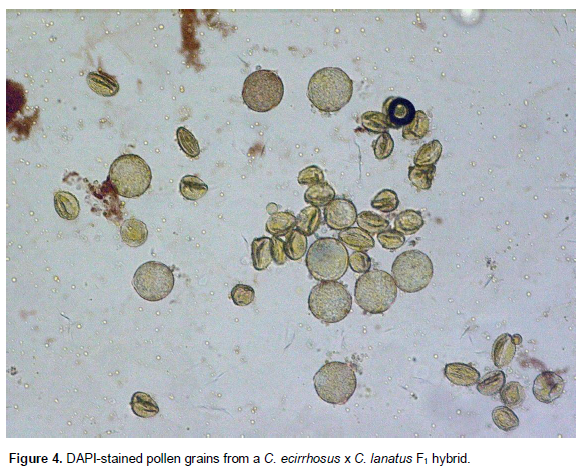
We have found (Jarret - unpublished) that F1 hybrids of C. ecirrhosus x C. lanatus, C. rehmii x C. lanatus and C. colocynthis x C. lanatus (and their reciprocal hybrids) have significantly reduced pollen viability (Figure 4). Although hybrid F1 seed may be readily produced, F2 seed produced from a selfed F1 C. lanatus x C. ecirrhosus had a very low rate of germination (< 5%). Fertility of the F2 plants is yet to be examined. Reduced pollen viability in hybrids does not preclude the use of the wild Citrullus species for the improvement of C. lanatus.
Intraspecific variation for the characteristics examined in this study is to be expected. However, only a limited amount of plant material of each species was available for examination. No commercial source of seed of either C. rehmii or C. ecirrhosus was identified. In as much as these two species could not be established in the field in Griffin, GA, an evaluation of the extent to which environ-mental variables affect the characteristics examined, was not attempted. Nevertheless, a further investment in support of efforts to identify and fully characterize the mechanisms endowing Citrullus spp. with drought tolerance could result in the identification of a trait or traits or gene(s) capable of enhancing drought tolerance in the cultivated crop. Such information might also be expected to increase our understanding of the fundamental processes involved in stress/drought tolerance in other cucurbitaceous crops.
CONFLICT OF INTERESTS
The author(s) have declared that there is no conflict of interests.
ACKNOWLEDGEMENTS
The author wish to acknowledge Dr. Ynes Ortega (University of Georgia, Department of Food Safety, Griffin, GA) for the generous use of equipment and facilities, and Mr. Chris Tatum (University of Georgia, Department of Plant Genetic Resources, Griffin, GA) for expert technical assistance.
REFERENCES
|
Akashi K, Miyake C, Yakota A (2001). Citrulline, a novel compatible solute in drought-tolerant wild watermelon leaves is an efficient hydroxyl radical scavenger. FEBS Lett. 508:438-442. Crossref |
||||
|
Akashi K, Nishimura N, Ishida Y, Yokota A (2004). Potent hydroxyl radical-scavenging activity of drought-induced type-2 metallothionein in wild watermelon. Biochem. Biophys. Res. Commun. 323:72-78. Crossref |
||||
|
Ali MA, Grace J (1986). Water use by the desert cucurbit Citrullus colocynthis (L.) Schrad. Oecologia. 70:475-480. Crossref |
||||
| Arnold TH, De Wet BC (1993). Plants of Southern Africa: Names and Distribution. Mem. Bot. Surv. S. Africa no. 62. | ||||
| Baker EA, Procopiou J (2000). The leaf and fruit cuticles of selected drought tolerant plant. In: Janoudi A (Ed.) Proceedings of the International Symposium on Growth and Development of Fruit Crops. Acta Hort. 527:85-93. | ||||
| Bates DM, Robinson RW (1995). Cucumbers, melons and water-melons In: Smartt J, Simmonds NW (Eds.) Evolution of Crop Plants, Longmans, London, UK. pp. 89-96. | ||||
| Benesova M, Hola D, Fischer L, Jedelsky PL, Hnilicka F, Wilhelmova N, Rothova O, Kocova M, Prochazkova D, Honnerova J, Fridrichova L, Hnilickova H (2012). The physiology and proteomics of drought tolerance in maize: Early stomatal closure as a cause of lower tolerance to short-term dehydration. PLos ONE 7(6):e38017. | ||||
|
Chomicki G, Renner S (2014). Watermelon origin solved with molecular phylogenetics including Linnaean material: another example of museomics. New Phytologist. Crossref |
||||
|
Condon AG, Richards RA, Rebetzke GJ, Farquhar GD (2004). Breeding for high water-use efficiency. J. Expt. Bot. 55:2447-2460. Crossref |
||||
| Dafni A, Pacini E, Nepi M (2005). Pollen and stigma biology In: Dafni A, Kevan PG and Husband BC (Eds.), Practical Pollination Biology. Enviroquest, Ltd., Cambridge, Ontario, Canada. pp. 83-129. | ||||
| De Winter B (1990). Notes on African plants. Cucurbitaceae. A new species from the Namib Desert, Namibia. Bothalia 20:209-211. | ||||
| Duke JA (1978) not cited. The quest for tolerant germplasm. In: Jung, GA (Ed.) Crop Tolerance to Suboptimal Land Conditions. American Society of Agronomy Special Symposium 32, Madison, WI. pp. 1-61. | ||||
| Duke JA (1983). Citrullus colocynthis In: Handbook of Energy Crops. (http://www.hort.purdue.edu/newcrop/duke_energy/dukeindex.html). | ||||
| Esau K (1977). Anatomy of Seed Plants, 2nd ed. John Wiley & Sons, NY. | ||||
|
Enslin PR, Rehm S (1958). The distribution and biogenesis of the cucurbitacins in relation to the taxonomy of the Cucurbitaceae. Proc. Linn. Soc. London. 169(3):230-238. Crossref |
||||
|
Food and Agriculture Organization of the United Nations (2014). FAOSTAT. |
||||
| Hameed M, Mansoor U, Ashraf M, Rao AUR (2002). Variation in leaf anatomy in wheat germplasm from varying drought-hit habitats. Int. J. Agric. Biol. 4:12-16. | ||||
|
Hameed M, Batool S, Naz N, Narwaz T, Ashraf M (2012). Leaf structural modifications for drought tolerance in some differentially adapted ecotypes of blue panic (Panicum antidotale Retz.). Acta Physiol. Plant 34:1479-1491. Crossref |
||||
|
Hussain AI, Rathore HA, Sattar MZA, Chatha SAS, Sarker SD, Gilani AH (2014). Citrullus colocynthis (L.) Schrad (bitter apple fruit): A review of its phytochemistry, pharmacology, traditional uses and nutritional potential. J. Ethnopharm. 155:54-66. Crossref |
||||
|
Jeffrey C (1961). Colocynthis and Citrullus. Taxon. 10:195-196. Crossref |
||||
|
Jeffrey C (1975). Further notes on Cucurbitaceae: III: Some southern African taxa. Kew Bull. 30:475-493. Crossref |
||||
|
Karaba A, Dixit S, Greco R, Aharoni A, Trijatmiko KR, Marsch-Martinez N, Krishnan A, Nataraja KN, Udayakumar M, Pereira A (2007). Improvement of water use efficiency in rice by expression of HARDY, an Arabidopsis drought and salt resristance gene. Proc. Natl. Acad. Sci. 25:15270-15275). Crossref |
||||
| Levi A, Wechter WP, Thies JA, Ling KS, Reddy UK, Xu Y, Guo S, Zhang X (2012). Watermelon. In: Wang YH, Behera TK, Cole C (Eds.) Genetics, Genomics and Breeding of Cucurbits. CRC Press, Enfield, NH. pp. 309-334. | ||||
| Liu DS, Yang WB, Zhao XQ (2008). Studies on watermelon drought tolerance identification indices and method in gravel-mulched land of Northwest China. China Vegetables 7:17-21 (in Chinese with English summary). | ||||
|
Liuigi C, Rizza F, Badeck FW, Mazzucotelli E, Mastrangelo AM, Francia E, Mare C, Tondelli A, Stanca AM (2008). Drought tolerance improvement in crop plants: An integrated view from breeding to genomics. Field Crops Res. 105:1-14. Crossref |
||||
| Martin JJ, Juniper DE (1974). The Cuticles of Plants. Edward Arnold, London. | ||||
|
Meeuse AD (1961). The Conservation of Citrullus. Taxon. 10:29-30. Crossref |
||||
| Meeuse AD (1962). The Cucurbitaceae of southern Africa. Bothalia 8:58. | ||||
| National Research Council (2006). Egusi In: Lost Crops of Africa, vol. II. National Academies Press, Washington, DC. | ||||
|
Navot N, Zamir D (1987). Isozyme and seed protein phylogeny of the genus Citrullus (Cucurbitaceae). Plant Syst. Evol. 156:61-67. Crossref |
||||
|
Renner SS, Pandey AK (2013). The Cucurbitaceae of India: Accepted names, synonyms, geographc distribution, and information on images and DNA sequences. PhytoKeys 20:53-118. Crossref |
||||
| Sarafis V (1999). Cucurbit resources in Namibia. In: Janick J (Ed.), Perspectives on new crops and new uses. ASHS Press, Alexandria, VA. pp. 400-402. | ||||
| Sass JE (1958). Botanical Microtechnique. 3rd ed. Iowa State College, Constable. 228pp. | ||||
| Singh AK (1990). Cytogenetics and evolution in the Cucurbitaceae In: Bates DM, Robinson RW, Jeffrey C (Eds.) Biology and Utilization of the Cucurbitaceae. Comstock Publishing, Ithaca, NY USA. pp. 10-28. | ||||
| Whitaker TW, Davis GN (1962). The cucurbits: Botany, cultivation and utilization. InterScience Publisher, NY. | ||||
| Zhang H, Gong G, Guo S, Ren Y, Xu Y (2011). Screening the USDA watermelon germplasm collection for drought tolerance at the seedling stage. HortSci. 46:1245-1248. | ||||
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


