Full Length Research Paper

ABSTRACT
The aim of the present study was to assess the correlation between the spatiotemporal variation of the gastropod Tricolia affinis and the macroalgal community on the sandstone reef at Piedade beach on the southern coast of Pernambuco State (PE) during the rainy and dry seasons in 2009 and 2010. Five square plots were randomly sampled on each date on the reef flat and pool to sample the gastropod population and measure the biomass of the dominant algae. Abiotic parameters were measured near the sampling sites. Three macroalgal genera, the Rhodophyta Gracilaria and Hypnea and Chlorophyta Bryopsis, showed representative biomasses at the sampling sites and were found throughout the sampling period. The abundance of T. affinis on the reef flat was significantly lower during the rainy season; however, no significant difference in this gastropod’s abundance was found between the reef and tide pool. The density of T. affinis was significantly correlated with the biomass of Gracilaria, which showed the highest biomass and structural complexity in the studied area.
Key words: Sandstone reefs, benthic algae, Gracilaria, mollusca, seasonality.
INTRODUCTION
Coastal regions and their associated reefs are among the zones that are most susceptible to environmental impacts because of the high human population densities and economic activities in these areas, including the existence of port complexes and tourism and leisure activities. These factors may cause changes in algal communities and the associated fauna, which are sensitive to human impacts (Sousa and Cocentino, 2004), such as trampling in the intertidal zone. Organic pollution is currently one of the main threats to coastal environments. This type of pollution can affect the development of benthic communities, causing a decrease in species diversity and potentially promoting the proliferation of opportunistic species, including ephemeral macroalgae (Breves-Ramos et al., 2005; Simões et al., 2009).
The sandstone reefs located on the southern coast of Pernambuco state are marine outcrops forming ridges parallel to the coast that often emerge during low tide. These reefs consist mainly of quartz and bioclasts, primarily originating from shellfish and algae (Sousa and Cocentino, 2004). Many of these reefs represent a key consolidated substratum for the attachment of benthic algae and a diverse associated fauna, therefore representing an ecosystem with significant biological interact-tions (Simões et al., 2009) that forms the base of the local food webs.
The Tricolia genus has a wide geographical distribution and is found in North Carolina (USA) to the coast of Brazil. The species Tricolia affinis is a prosobranchiate gastropod of the family Tricoliidae found at coastal locations in Florida, the Caribbean, Colombia, Venezuela and Brazil, exhibiting sizes varying between 0.2 and 4.0 mm (Rios, 1994). On the Brazilian coast, this genus is found from Santa Catarina State to the northeastern region, including the Fernando de Noronha archipelago (Nogueira et al., 2003). According to Braga (1983), T. affinis is an abundant species in the studied area (Piedade beach). This gastropod is commonly found in estuaries and on rocky shores, coral reefs, unconsolidated substrata and seagrass banks, often associated with algae.
T. affinis is an herbivorous species that is most frequently found in the subtidal zone, where there are higher nutrient concentrations. This species feeds on microscopic algae and bacteria (periphyton) on the surface of debris-covered macroalgae (Marcus and Marcus, 1960). According to Ourives et al. (2011), T. affinis is also an abundant component of the estuarine fauna of Rio Cachoeira, Bahia State, Brazil, where its distribution was found to be positively correlated with salinity. The aim of the present study was to assess the temporal and spatial variation of T. affinis, associated with a Gracilaria bank, in the intertidal zone of the reef at Piedade beach since, this alga has been commercially used on the Brazilian coast, for agar production, and its abundance was found to be reduced in several locations along the northeastern coast. While in other areas, it is considered as an invasive species (Fukunaga et al., 2014; Zi-Min and Lopez-Bautista, 2014).
Two areas were sampled: a tidal pool where the algae were always immersed and more likely subjected to water transport, the reef flat algae where they are better attached to the substrate and subjected to less water transport. Therefore, the present study was based on the hypotheses that a correlation exists between the dominant algal biomass, abiotic factors and the abundance of T. affinis at the sampling sites and that spatial and temporal differences exists in the mollusk and algae abundance.
MATERIALS AND METHODS
Study area
Piedade beach in the Jaboatão dos Guararapes municipality is located on the southern coast of Pernambuco state, between the coordinates 08° 11´ 08.48´´ S, 34° 55´ 04.66´´ W and 08° 11´ 03.45´´ S, 34° 55´ 03.24´´ W. Its reef area is approximately 9.8 km in length and encompasses approximately 234 km², with a formation running parallel to the coast. The reef is immersed at high tides, forming a lagoon between the beach and the reef at low tides. The region exhibits a tropical Atlantic climate and average air temperature near 26°C, and the distribution of rainfall defines two seasons: a dry season (September-February), with rainfall below 100 mm, and a rainy season (March-August), with rainfall greater than 100 mm (Cavalcanti and Kempf, 1969). During the study period, the precipitation in Piedade was highest in April 2009 (268.9 mm) and lowest in October 2009 (6.0 mm) according to data from the local meteorological station. The coast of Piedade consists of a coastal plain formed by fluvial and marine deposits, where there was once a strip of typical coastal vegetation referred to as restinga that is now occupied by quickly expanding real estate developments and is still influenced by the Jaboatão River, which receives pollution from industrial hubs and urban centers (Pereira et al., 2002).
Field work
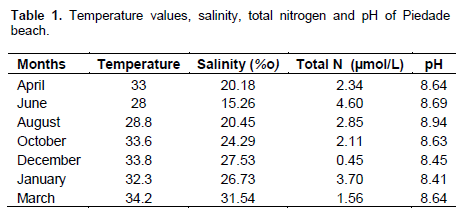
Sampling was conducted in the intertidal zone of the Piedade reef (PE) from April 2009 to March 2010. The first sampling site was a reef pool at the coordinates 8° 10´ 48.8´´ S and 34° 54´ 57.9´´ W. The second site was located on the reef flat in an area dominated by Gracilaria caudata (8° 10´ 49.4´´ S and 34° 54´ 58´´W) and a few meters away from the tide pool. Five 20 x 20 cm squares were randomly sampled using a spatula at each study site on each sampling date. Samples were collected in April, June, August, October and December 2009 and January and March 2010. The collected material was packed in plastic bags, labeled with the date, sampling site and square number and then immediately transported to the University of Pernambuco Marine Biology laboratory for freezing and subsequent triage. The water temperature and soluble oxygen were measured at the sampling sites near the square plots with an oximeter. The soluble oxygen values ??ranged between 9.2 and 18.1 mg/L at the sampling sites, and thus, oxygen was not a limiting factor for the T. affinis populations. Water samples were collected in an area near the sampling sites where there is water influx to the reef flat, through an opening on the edge of the reef, for subsequent analysis of salinity, pH and nitrogen concentrations (ammonia-N [NH3-+NH4+.N], nitrite-N [NO2-.N] and nitrate-N [NO3-.N]) at the Chemical Oceanography Laboratory of the Federal University of Pernambuco (Table 1). The salinity and nutrient analyses were conducted according to Strickland and Parsons (1972).
Laboratory and statistical analyses
The mollusks were separated on a plastic tray using a stereomicroscope, metallic tweezers and Petri dishes. Then, they were fixed in 70% ethanol for subsequent counting. Only T. affinis samples in good condition and with an intact operculum were included in the counts, with empty shells being excluded. The dominant algae were identified taxonomically at the genus and species level. Algae were transferred to laminated paper bags and dried at 60°C for 72 h to assess their biomass. The samples were then weighed on an analytical scale, and the algal biomass values were expressed per m². Bivariate and univariate analyses of variance (ANOVAs), in addition to Tukey’s tests, were used to assess the seasonal variation of algae at the reef flat and tide pool after logarithmic transformation of the biomass data.
Two-way ANOVAs were performed to assess the spatiotemporal variability of the T. affinis density in the samples, following confirmation of the normal distribution of the data and square root (x) + 0.5 transformation. Subsequently, one-way ANOVAs were performed for each sampling station individually, in addition to Tukey’s test. Canonical correspodence analyses were performed with the mean algal biomass and the mean density of T. affinis with the abiotic variables (water temperature, salinity, pH and total nitrogen) separately for the two sampling sites. The Pearson correlation coefficients between the dominant algal biomass and abundance of T. affinis were assessed. The statistical programs R (R Core Team, 2014) and Biostat 5.8.4 were used for these analyses.
RESULTS AND DISCUSSION
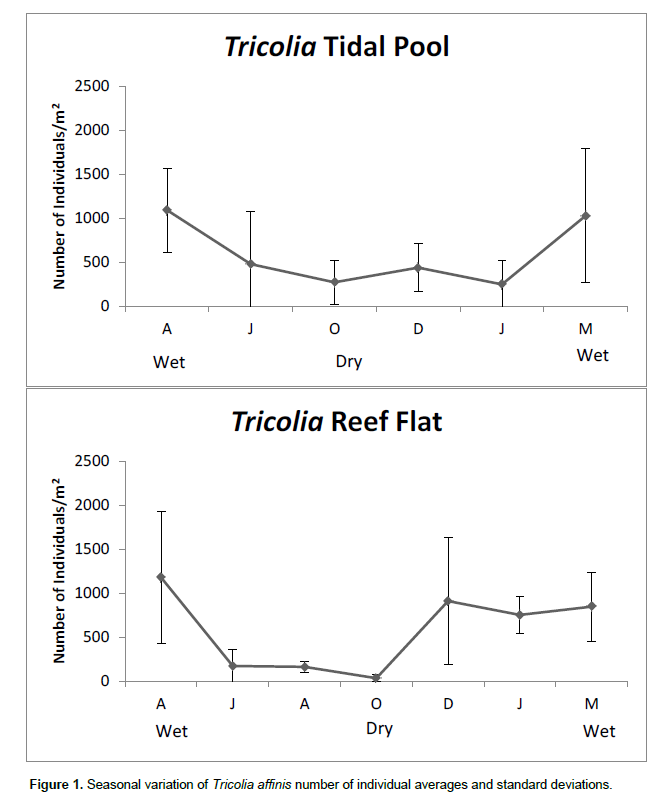
A two-way ANOVA showed that there was significant temporal variation in the T. affinis density, but no significant differences were found in gastropod abundance between the sampling sites. The temporal variation of Tricolia was significant in the reef flat samples, significant differences between the sampling months were found (p < 0.01) (Figure 1). The mean gastropod density on the reef flat differed between April 2009 and June, August and October 2009. A significant difference was also found between October 2009 and March 2010, when the mean density of Tricolia was approximately 850 individuals per m2, demonstrating a population recovery at the end of the dry season.
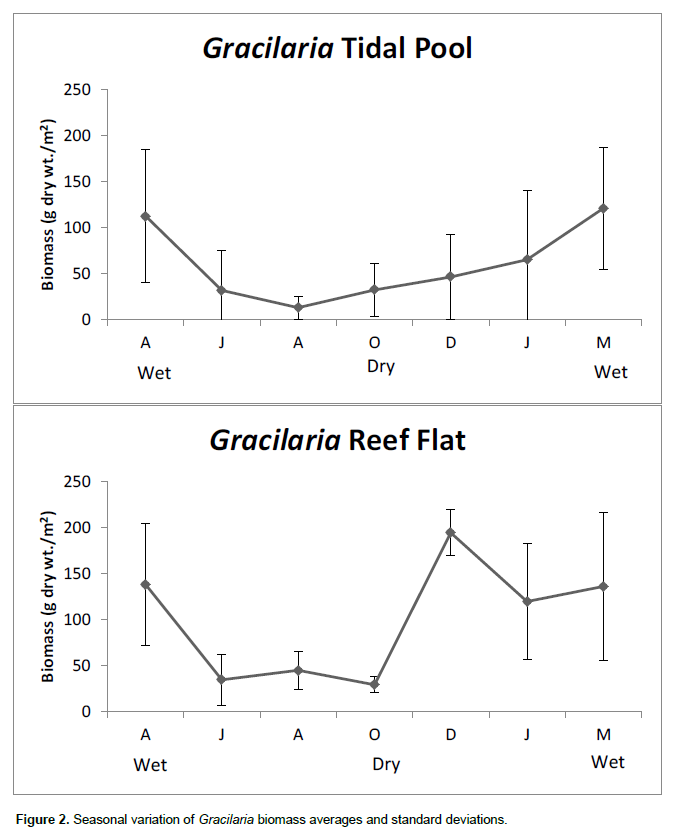
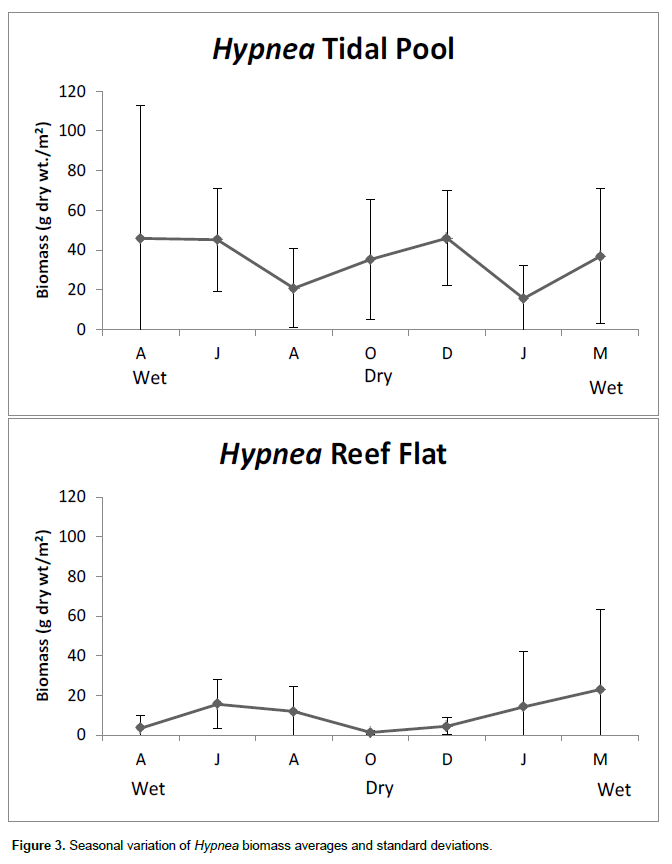
The Rhodophyta G. caudata, Hypnea (Hypnea musciformis, Hypnea spinella and Hypnea cenomice) and the Chlorophyta Bryopsis (Bryopsis plumosa and Bryopsis pennata) were the dominant algae in terms of biomass (Figures 2, 3 and 4). These algal genera were found on all of the sampling dates at both the tide pool and reef flat. Their mean dry weights were subjected to statistical analyses in combination with the density of T. affinis. Ulva spp. was also found during all of the sampling campaigns on Piedade reef, but at a lower biomass than Gracilaria, Hypnea and Bryopsis and was therefore not included in the statistical analyses. The macroalgae showed seasonal variation, reflected by a lower biomass of G.caudata in rainy season samples (p < 0.05). However, the biomass of Hypnea showed no significant differences between the sampling months at either the tide pool or reef flat (p > 0.05) but was different between sites. No significant differences in Bryopsis biomass were found on the reef flat between the sampling dates (p > 0.05). In turn, Bryopsis showed a peak biomass in December 2009, which was significantly higher (p < 0.01) than the mean biomasses detected in the other sampling months at the tide pool, although no differences in biomass were found between sampling sites. A two-way ANOVA demonstrated a higher abundance of G. caudata at the reef flat than in the pool.

The T. affinis density was positively correlated with the Gracilaria biomass for the samples collected at both the reef flat (r = 0.91, p < 0.01) and tide pool (r = 0.88, p < 0.05). The salinity in the study area was low (15.26 – 20.45‰) during the rainy season, which may have been a limitation for T. affinis populations. The canonical correspondence analysis for the reef pool showed the correlation between total nitrogen and Hypnea biomass and Tricolia density close to Gracilaria biomass (Figure 5) where the mollusk was less variable, in the same site Gracilaria is opposite to Bryopsis. In the reef flat, salinity is close to Gracilaria which is opposite of Hypnea and Bryopsis (Figure 6).
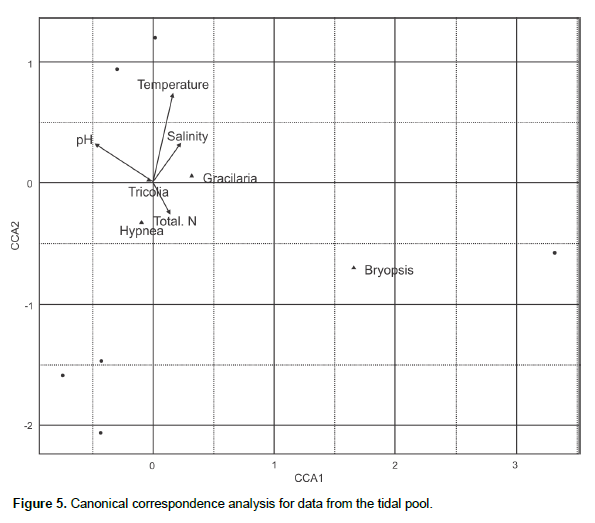
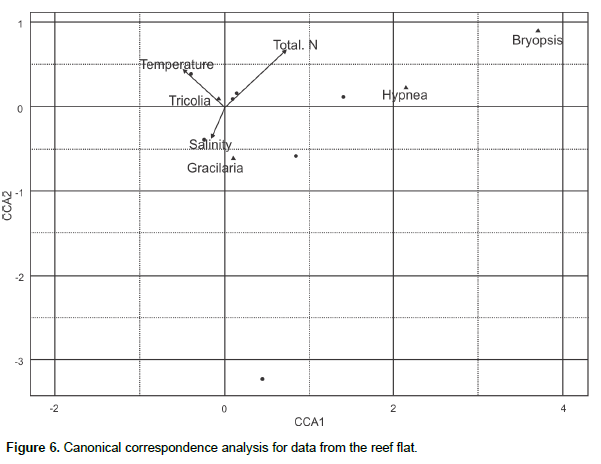
The opposite canonical correspondence analyses ordering of algal genera indicates competition between functional groups (Gao et al., 2014), evidenced also by the analyses of variance since Hypnea spp. was more abundant at the tidal pool whereas G.caudata biomass was greater at the reef flat. A two-way ANOVA showed that there was temporal variation in the T. affinis density; however, there was no significant difference in the gastropod’s abundance between the two sampling sites. We can therefore conclude that the spatial distribution of T. affinis was proportional at the two sampling sites. Therefore, a correlation between T. affinis density and G. caudata biomass was found, at the reef flat and pool, possibly related to the alga’s higher structural complexity or the provision of greater protection for recruitment and from predation and ocean currents, in addition to being a reliable source of food. Gracilaria exhibited a lower biomass in the samples from the rainy period, directly affecting the number of sampled Tricolia. Gracilaria’s stem complexity confirms the correlation of T. affinis with its structural composition, which may be related to the retention of sediments that provide a greater supply of nutrients and protection against predation and hydro-dynamic counterflows for the mollusk (Chemello and Milazzo, 2002). Pereira and Jacobucci (2008) suggested that although T.affinis has been registered as a diet staple for fishes and crabs and an important link in the food chain of aquatic environments, it may not play a key role in the diets of marine species due to its low nutritional quality and calcareous shell (Dubiski-Silva and Setuko, 2008). The study area is a highly impacted region, constant trampling by bathers, which may be one reason for the success of the gastropod and relationship with Gracilaria. Invasive Gracilaria salicornia in the coast of Hawaii improved the density of epifauna and detritivores in comparison with Padina thivyi, while the invasive Acanthophora spicifera did not cause any epifauna improvement (Fukunaga et al., 2014).
The density of this gastropod was positively correlated with the biomass of G. caudata which showed the highest dry weight values at the sampling sites and presented a significantly higher biomass at the reef flat than the pool, where the temporal variation in Tricolia was less evident. Along with Tricolia, G. caudata showed seasonal varia-tion between the dry and rainy periods, with a higher biomass found in dry periods, when there was also a higher temperature and salinity (Silva et al. 1987; Marinho-Soriano et al., 2001). Pereira et al. (2010) found no seasonal variations in T. affinis density or a positive correlation with Sargassum spp. biomass; however, these authors observed a constant presence of juvenile T. affinis throughout the year in the phytal fauna of Sargassum, indicating that factors such as habitat availability may affect this gastropod’s abundance and recruitment. On the Piedade reef, climatic and hydrodynamic factors should be considered, because they may affect the dispersal of organisms in the reef pool, thereby causing a greater variability in the population distribution of T. affinis in the tide pool, in addition to the trampling of the reef flat.
CONFLICT OF INTERESTS
The authors declare that there is no conflict of interests.
ACKNOWLEDGEMENT
This study was partially funded by the National Council for Scientific and Technological Development [CNPq] Universal Announcement with process number 470424/2007-9.
REFERENCES
| Braga RAP (1983). Participation of Tricolia affinis C. B, Adams, 1850 (Mollusca, Gastropoda) in the community of vagil animals associated to macroalgae in Piedade (PE) reefs. An. Soc. Nord. Ecol. 1:80-81. | ||||
|
Breves-Ramos A, Lavrado HP, Junqueira AO, Silva SHG (2005). Succession in rocky intertidal benthic communities in areas with different pollution levels at Guanabara Bay (RJ-Brazil). Braz. Arch. Biol.Tech. 48:951-965. Crossref |
||||
| Cavalcanti LB, Kempf M (1969). Study of the continental plataform in the area of Recife (Brasil). II. Meteorologia e Hidrologia. Trab. Oceanogr. 9:149-158. | ||||
|
Chemello R, Milazzo M (2002). Effect of algal architecture on associated fauna: some evidence from phytal molluscs. Mar. Biol. 140:981-990. Crossref |
||||
| Dubiski-Silva J, Setuko M (2008). Natural diet of fish and crabs associated with the phytal community of Sargassum cymosum C. Agardh, 1820 (Phaeophyta, Fucales) at Ponta das Garoupas, Bombinhas, Santa Catarina State, Brazil. J. Nat. Hist. 42:27-28. | ||||
|
Fukunaga A, Peyton, KA, Thomas FIM (2014). Epifaunal community structure and ammonium uptake compared for the invasive algae, Gracilaria salicornia and Acanthophora specifera and the native alga, Padina Thivyi. J. Exp. Mar. Biol. Ecol. 456:78-86. Crossref |
||||
|
Gao Z, Xu D, Meng C, Zhang X, Wang Y, Li D, Zou J, Zhuang Z, Ye N (2014). The green tide-forming macroalga Ulva linza outcompetes the red macroalga Gracilaria lemaneiformis via allelopathy and fast nutrientes uptake. Aquat. Ecol. 48:53-62. Crossref |
||||
|
Marinho-Soriano E, Silva TSF, Moreira WSC (2001). Seasonal variation in the biomass and agar yield from Gracilaria cervicornis and Hydropuntia cornea from Brazil. Biores. Techn. 77:115-201. Crossref |
||||
| Marcus E, Marcus E (1960). On Tricolia affinis cruenta. Bol. Facul. Fil. Ciênc. Let. 23:171-198. | ||||
| Nogueira EMS, Freitas L M, Jesus IML, Vieira DM (2003). Tricolia Risso, 1826 (Mollusca – Gastropoda – Phasianellidae) in the Paripueira beach, north litoral from Alagoas. Bol. Lab. Hidrobiol. 16:1-4. | ||||
|
Ourives TM, Rizzo AE, Boehs G (2011). Composition and distribution of the benthic macrofauna in the Cachoeira River estuary, Ilhéus, Bahia, Brazil. Rev. Biol. Mar. Oceanogr. 46:17-25. Crossref |
||||
| Pereira PHC, Jacobucci GB (2008). Diet and feeding behaviour of Malacoctenus delalandii (Perciformes: Labrisomidae). Biot. Neot. 8:1-9. | ||||
| Pereira PHC, Biasi PC, Jacobucci GB (2010). Seasonal population dynamics and spatial distribution of Tricolia affinis (Mollusca: Gastropoda) associated to Sargassum spp. in the north litoral from São Paulo. Ver. Bras. Zoociec. 12:7-16. | ||||
| Pereira SMB, Oliveira-Carvalho, MF, Angeiras JAP, Bandeira-Pedrosa ME, Oliveira N MB, Torres J, Gestinari LMS, Cocentino ALM, Santos MD, Nascimento PRF, Cavalcanti RR (2002). Diagnosis of the Biodiversity of Pernambuco. Recife: Massangana, pp. 97-124. | ||||
| Rios EC (1994). Seashells of Brazil. Museu oceanográfico: Rio Grande: Ed. FURG, 368p. | ||||
|
Silva RL, Pereira SMB, Oliveira EC, Eston VR (1987). Structure of a bed of Gracilaria spp. (Rhodophyta) in northeastern Brazil. Bot. Mar. 30:517-523. Crossref |
||||
|
Simões IP, Guimaraens MA, Oliveira-Carvalho MF, Valdevino J, Pereira SMB (2009). Floristic and ecological succession of the macroalga in reefs Piedade – PE beach. Neot. Biol. Conserv. 4:49-56. Crossref |
||||
| Sousa GS, Cocentino ALM (2004). Macroalgae as environmental quality indicators. Trop. Oceanogr. 32:1-22. | ||||
| Strickland JDH, Parsons TR (1972). A practical handbook of seawater analysis. Ottawa: Bulletin 167 Fisheries Research Board of Canada, 310p. | ||||
| Zi-Min H, Lopez-Bautista J (2014). Adaptation mechanisms and ecological consequences of seaweed invasions: a review case of the agarophyte Gracilaria vermiculophilla. Biol. Inv. 16:967-976. | ||||
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


