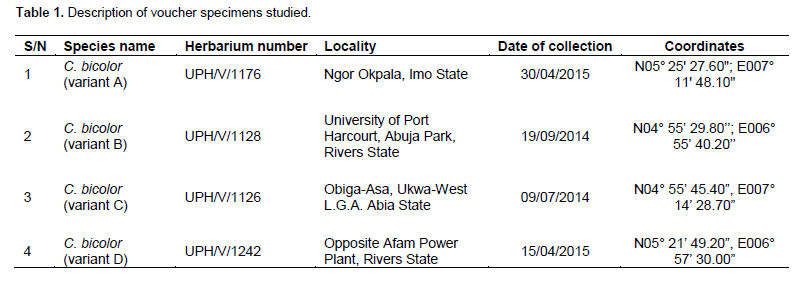
Full Length Research Paper

ABSTRACT
Four Caladium bicolor variants collected from different parts of Nigeria were subjected to anatomical comparison to enhance the taxonomic status of the species. Fresh samples (leaf and petiole) of these variants were fixed in formalin, acetic acid and alcohol (FAA), dehydrated in alcohol series, peeled or sectioned. Peeled specimens were stained with safranin, while sectioned ones were stained with Alcian blue and counterstained with safranin. Good preparations were mounted on slides, viewed and photographed with Optika B-1000 FL LED microscope. Epidermal cells from the variants are mainly pentagonal-hexagonal but rarely heptagonal while the anticlinal cell walls are mainly straight and partly arced/curved. Variants A, B and D are amphistomatic while variant C is hypostomatic. Isotricytic, anisocytic, tetracytic, anomocytic and contiguous stomata were observed among the taxa. The stomata index (SI) varied from 4.35 to 11.76 on the adaxial surface, and from 6.25 to 47.62 on the abaxial surface. Calcium oxalate crystals (druses, 8.18 to 19.09 µm and raphides, 21.82 to 68.18 µm) occur in all variants. Raphides are predominantly found in the petiole while druses and raphides are found in the midrib and petiole. The shapes of the adaxial surface of the midrib are relatively different from each other, and include curved (convex) and flat surfaces. The leaf lamina comprised one layer of palisade and spongy mesophylls each. The number of vascular bundles varied from 11 to 23 in the midrib. These characters can be used to distinguish these taxa especially when combined with the existing data on the species. The similarities among these variants point towards the same evolutionary origin; however may suggest that intraspecific or interspecific hybridizations may have produced the variants.
Key words: Amphistomatic, Caladium, druses, hypostomatic, midrib, petiole, raphides.
INTRODUCTION
Caladium bicolor (Aiton) Vent. belongs to the family Araceae and is characterized by various leaf colourations (Hutchinson and Dalziel, 1954). There are over 2000 cultivar names for Caladium, and over a hundred cultivars are grown today (Wilfret, 1993). The genus Caladium comprises 12 species (Mayo et al., 1997), of which C. bicolor is the major source of cultivars. C. picturatum and C. marmoratum are now considered synonyms of C. bicolor (Madison, 1981). It is a monotypic genus that is found in most West African countries (Hutchinson and Dalziel, 1954).
In South and Central America other species of Caladium like C. marmoratum Mathieu, C. picturatum C. Koch, and C. schomburgkii Schott including Caladium bicolor (Aiton) Vent. have been reported (Birdsey, 1951; Hayward, 1950; Wilfret, 1993). Breeding of this species of plant has led to many variants (Wilfret, 1993). Also, intraspecific or interspecific hybridizations among these American species may have produced the cultivated species C. hortulanum and other variants of economic values (Hayward, 1950; Birdsey, 1951; Wilfret, 1993; Deng and Harbaugh, 2006a). Members of this genus are used as ornamentals and other economic purposes (Evans et al., 1992; Deng et al., 2005).
The anatomy of vascular and support tissues in the leaf and petiole in Araceaea showed and inferred an interesting relationship among them (Keating, 2000, 2002, 2004; Goncalves et al., 2004). These could suggest correlations between changes in vascular bundle and collenchyma characters and the appearance and further evolution of the unisexual-flowered, aperigoniate aroids (Natalie et al., 2011). Morpho-anatomical character patterns seem to imply a major adaptive shift in the evolution of aroids (Hesse, 2006a, b), and could be used to distinguish closely related plants (Kemka-Evans et al., 2014; Osuji and Nwala, 2015).
The naming and identification of C. bicolor cultivars is difficult. This is due in part to the lack of up-to-date reference material which illustrates all cultivars in colour, as well as the sale of cultivars without reliable names (Jin et al., 1999). Among the Araceae family, calcium oxalate deposits can be diagnostic, including their presence, type, diversity, occurrence and distribution have been noted to enhance the delimitation of members of this family (Osuji and Nwala, 2015; Osuji, 2013; Nurul et al., 2013; Mais and Amal, 2012; Gary, 2009) and raphide idioblasts are known as storage facilities for Aroids (Okoli, 1988; Okoli and Green, 1987; Okoli and McEuen, 1986).
Except the morphological description by Metcalfe and Chalk (1968), there is a dearth of information on the biology and taxonomic status of Nigerian species of Caladium. The ornamental and economic values of these species and their variants underscore the need for their taxonomic characterization using different systematic lines of study. This study bridges the gap by providing information on the biology and comparative anatomical features (including calcium oxalate crystal types) of four C. bicolor variants in Nigeria. The results of the study shall enhance the establishment of the taxonomic status of the species.
MATERIALS AND METHODS
Source of materials
The materials for this study were collected from different parts of the country (Table 1), properly identified using Flora of West Tropical Africa (Hutchinson and Dalziel, 1954), and deposited at the University of Port Harcourt Herbarium. Live specimens are being maintained at the Ecological Center, University of Port Harcourt.
Petiole and midrib studies
Subsamples from the original materials (Table 1) were fixed in formalin, acetic acid and alcohol (FAA) for 12 h. Thereafter, the specimens were dehydrated in ethanol series of different concentration (30 and 50%) and stored in 70% ethanol until when needed. The leaf portion of the petiole, and the central portion of the midrib were sectioned following the method of Agbagwa et al. (2007). The sections were stained in Alcian blue and counter-stained with 1% Safranin red for two minutes, mounted on a slide, viewed and photographed with an Optika B-1000 FL LED microscope.
Epidermal studies
Foliar materials for epidermal studies were collected fresh from plants in the Ecological Centre. 0.5 to 1 cm square leaf cuttings were obtained from identical regions of each fresh leaf, generally from mid-way between the leaf base and apex of lamina including the mid-rib. The adaxial and abaxial epidermal peels were obtained using sharp pointed forceps. Peels were stained with 1% safranin or alcian blue, rinsed with distilled water to remove excess stain and mounted in a drop of pure glycerol on clean glass slides. A cover glass was placed over the drop and sealed with nail vanish to prevent dehydration (Okoli and Ndukwu, 1992). The epidermal features that were observed include: organization of the epidermis, arrangement of the epidermal cells, nature of trichomes, shape of epidermal cells and nature of the anticlinal cell wall of the leaf epidermis, stomatal types, density and index. The stomatal index (SI) was determined based on Metcalfe and Chalk (1979), while the terminology for the stomatal type is taken after Malvey (2004).
Statistical analysis
The average (mean), standard deviation (STD) and range were determined using IBM SPSS Statistics 20.
RESULTS AND DISCUSSION
Four variants of C. bicolor were identified. These specimens display a variety of colors including red, white, pink, yellow and green, and are tagged C. bicolor var. A, B, C and D (Figure 1). The coloration of the collections conformed to previous reports by Metcalfe and Chalk (1968). These authors reported that the genus Caladium in West Africa is monotypic with various colorations.
.png)
Epidermal characteristics
No variation was observed on the shapes of the leaf epidermal cell among the different variants of Caladium studied. For instance, the shape of the upper (adaxial) and the lower (abaxial) epidermal surfaces in variant A, C and D are mainly pentagonal-hexagonal but rarely heptagonal, while the anticlinal cell wall are mainly straight and partly arced/curved. However, in variant B the shape of the adaxial and the abaxial epidermal surfaces are pentagonal-hexagonal (Figures 2 and 3, Table 2).
Variants A, B and D are amphistomatic (with stomata on both surface of the leaf) while variant C is hypostomatic (stomata on only one surface). This character makes this variant different from the other variants studied. The stomata types observed among the different Caladium variants are isotricytic, anisocytic, tetracytic, anomocytic and contiguous (Figures 2 and 3, Table 2). Among these stomata types, tetracytic and anomocytic stomata were dominant (Figures 2 and 3) and were observed on both leaf surfaces of variants A, B and D while in variant C they were recorded only on the lower surface (Table 2).
.png)
.png)
.png)
Isotricytic and anisocytic occurred only on the lower leaf surface of variant D. The occurrence of the same stomata type among these variants of Caladium indicates very close phylogeny as shown in Sphenostylis stenocarpa (Hochst ex A. Rich) Harms by (Nyananyo and Osuji, 2007). The presence of isotricytic, anisocytic and contiguous stomata on variant D distinguishes it from the other variants. In the same vein, the presence of brachyparacytic stomata in the abaxial leaf epidermis of an accession of Xanthosoma ‘Ede Uhie’ and its absence in other cultivars indicate divergent advancement in its evolution (Osuji and Nwala, 2015).
This suggests evolutionary divergence between Caladium variant D and the other variants. In a similar study among the Araceae, different stomata types namely anomocytic, actinocytic, paracytic, cyclocytic and transitionary types between paracytic and cyclocytic have been noted (Wang and Zhao, 2002). They observed that the epidermal cells are nearly isodiametric in outline with straight, arched and undulate anticlinal walls. Striate ornamentation occurs on periclinal walls of epidermal cells in some species. They noted that though the stomatal apparatus types in Araceae are of little taxonomical significance at intra-family level of Araceae, the combined characters of stomatal apparatuses, the shape of anticlinal wall and ornamentation of cuticules in guard cells may be useful for species identification (Wang and Zhao, 2002; Osuji and Nwala, 2015).
The stomata index (SI) varied from one variant to another. For instance, the stomatal index on the abaxial surface include- 25.93 to 37.5 (31.26±4.05), 22.22 to 37.50 (30.92±5.93), 6.25 to 36.36 (19.93±10.15) and 26.67 to 47.62 (36.64±7.68) for variants A, B, C and D respectively (Table 3). On the other hand, the stomatal index on the adaxial surface of the variants varied from 4.55 to 8.82 (6.31±2.24) in variant D to 6.25 to 8.11 (7.08±0.94) in variant A, and 4.35 to 11.76 (6.94±3.01) in B. The differences in stomata types, their distribution and indices on the leaf surfaces of these Caladium variants are diagnostic and could be used to distinguish among variants.
.png)
In this study, variants A and B have the same stomata types but the indices varied slightly from each other. Also, variant C has the same stomata types with A and B but it is hypostomatic while variant D has the same stomata types with these variants in addition to isotricytic, anisocytic and contiguous stomata. This finding however conforms with Osuji and Nwala (2015) who noted the presence of stomata on both upper and lower epidermis of both Colocasia and Xanthosoma spp. They further noted that stomata were more on the lower epidermis than the upper epidermis. Also, they observed epidermal variations and differences in stomatal indexes within the cultivars of Xanthosoma and Colocasia and suggested that this could account for their ecological adaptation to variation in the degree of wetness of the environment. Also, stomata types and nature of the epidermal cells have been of diagnostic importance in other members of angiosperm namely Emilia (Ndukwu and Agbagwa, 2006), Abrus (Agbagwa and Okoli, 2006), Vernonia (Kemka-Evans et al., 2014) and Ixora (Essiett and Umoh, 2014).
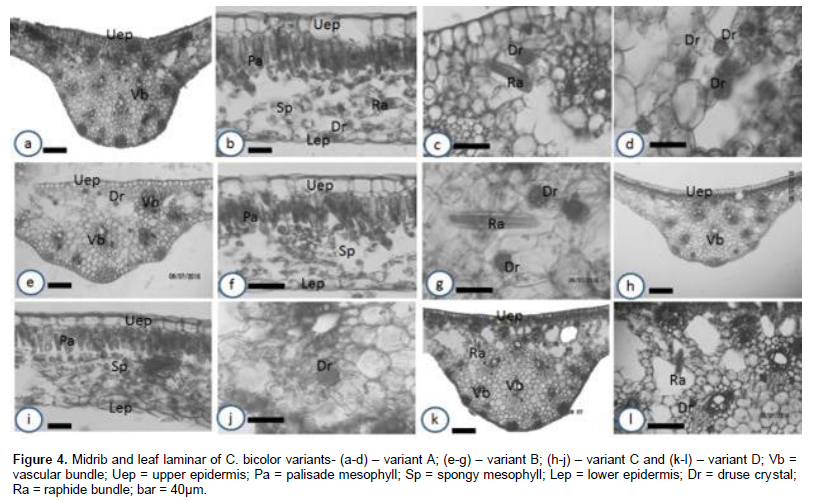
Shape of midrib and petiole
The shapes of the adaxial surface of the midrib are relatively different from each other. In variants A (Figure 4a) and C (Figure 4h), the shapes are curved (convex); in variant B (Figure 4e), it is flat while in variant D it is relatively flat (Figure 4k). This character is fairly diagnostic among the different variants. The leaf lamina comprised one layer of palisade and spongy mesophylls each (Figures 4b, f and i). The upper epidermal cells in the lamina of all the variants are isodiametric or oval. They elongate periclinally (Figures 4b, f and i). Also, the shapes of the petiole in all the variants studied are oval (Figures 5a, c, e and g). These similarities confirm that the variants have the same evolutionary origin and are of the same monotypic taxon (Metcalfe and Chalk, 1968); however intraspecific hybridizations may have produced the variants (Hayward, 1950; Birdsey, 1951; Wilfret, 1993; Deng and Harbaugh, 2006b).
.png)
.png)
.png)
.png)
.png)
.png)
.png)
.png)
.png)
.png)
.png)
.png)
Number of vascular bundle in the midrib and petiole
Variations in number and arrangement of vascular bundles in petiole and midrib have been used to differentiate species of the same genus or family (Ekeke and Mensah, 2015; Agbagwa and Ndukwu, 2004; Metcalfe and Chalk, 1979). For instance, in their studies, Ekeke and Mensah (2015) noted that the number of vascular traces in the midrib of members of Asteraceae varied from one species to another and is diagnostic. Also, Agbagwa and Ndukwu (2004) reported that the number of vascular bundles in the petioles of members of Cucurbita could be used to distinguish them (C. moschata, 10; C. maxima, 16 and C. pepo, 14).
In this study, the number of vascular bundles in the midribs and petioles varied from one variant to another (Table 3). The number of vascular bundles in the petiole of the different variants are variant A (19), variant B (27), variant C (22) and variant D (22). In the midrib (Table 3), variant A has 19 vascular bundles, variant B (11), variant C (13) and variant D (23). This variation in the number of vascular bundles is diagnostic and could be used to distinguish the different Caladium variants. Though variants C and D had the same number of petiolar vascular bundle, the number of vascular bundle in their midrib differed and could be used to differentiate them. This finding therefore supports that the variation in number of vascular bundle in the petiole and midrib are diagnostic (Ekeke and Mensah, 2015; Agbagwa and Ndukwu, 2004; Metcalfe and Chalk, 1979) and however may suggest that intraspecific or interspecific hybridizations may have produced the variants.
.png)
Calcium oxalate types and sizes
Two main calcium oxalate crystal types were observed among the taxa studied. These are druses and raphides (Table 4, Figures 4 and 5). The druses are ubiquitous, raphides are predominantly found in the petiole while druses and raphides are found in the midrib. The occurrence of some of these crystals is specific or restricted to some tissues. For instance, among the taxa studied, the occurrence of the raphides is specific. They are found in the midrib and petiole but not in the lamina. This observation has been made in Dieffenbachia seguine (Araceae) (Gray, 2009). He noted that druses appear to be nearly ubiquitous throughout this plant occurring in most of plant parts except in the ovaries.
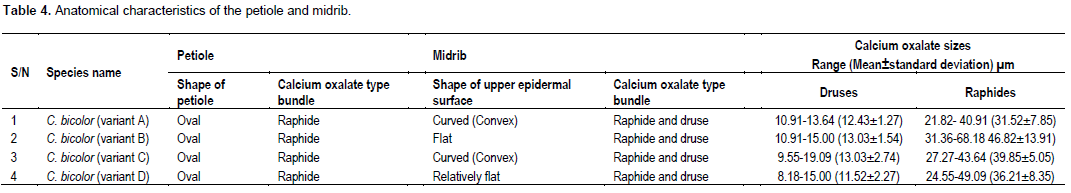
Furthermore, he observed that different portions of the same organ may have different crystal types such as the leaf margins which have a greater density of druses than the lamina, as well as overlapping raphide bundles not present in the lamina (Gray, 2009).
In the lamina, the calcium oxalates (druses) are embedded in the palisade and spongy mesophylls. They are identified by their dark and spiny nature. The accumulation of the calcium oxalate crystals in the leaf suggests that the taxa are poisonous. This explains why the leaves are not normally consumed by animals and humans except after being properly processed. These crystals could therefore be a source of defense to the plant against herbivores. This observation is in line with previous studies on Xanthosoma and Colocasia (Osuji, 2013; Uno et al., 2001).
The size of druses varied from 8.18 µm in variant D to 19.09 µm in variant C which corresponds to smallest and largest sizes of druse found in Caladium variants studied (Table 3). However, the sizes of the raphides ranged from 21.28 µm in variant A to 68.18µm in variant B (Table 4). In similar study in Araceae, Gray (2009) recorded different sizes of crystals in Dieffenbachia seguine. In this present study, the sizes of the calcium oxalate observed among the different variants include; druses (8.18 µm to 19.09 µm) and raphides (21.28 µm to 68.18 µm). These ranges fall within 1.0 µm to 250 µm recorded in Amorphophallus mueleri (Nurul et al., 2013).
Though there is difference between this value and that recorded in Caladium variants, this result supports the placement of both genera in the same family as having the same evolutionary origin. They are genetically controlled (Mais and Amal, 2012) and are useful in germplasm characterization and classification. Furthermore, the result conforms to the previous work in Araceae in the sense that druses and raphides were observed but in contrast because prismatic and styloid were not observed (Osuji, 2013; Nurul et al., 2013; Mais and Amal, 2012; Gray, 2009).
CONCLUSION
This study has revealed the different calcium oxalate crystals, their sizes and occurrence including the epidermal and anatomical characteristics of leaf, petiole and midrib among the different variants of Caladium bicolor in Nigeria. These are useful taxonomic characters for delimiting the variants especially when combined with the existing data on the species.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGMENT
The authors are most grateful to the Department of Plant Science and Biotechnology, University of Port Harcourt for providing laboratory facilities used in this study.
REFERENCES
|
Agbagwa IO, Ndukwu BC (2004). The value of morpho-anatomical features in the systematics of Cucurbita L. (Cucurbitaceae) species in Nigeria. Afr. J. Biotechnol. 3(10): 541-546. |
|
|
Agbagwa IO, Okoli BE (2006). Leaf Epidermal Micromorphology in the Systematics of Abrus Adanson (Papilionaceae) in Parts of Tropical West Africa. Asian J. Plant Sci. 5:41-49. |
|
|
Agbagwa IO, Okoli BE, Ndukwu BC (2007). Comparative Anatomy of Abrus Adanson species in Parts of Tropical West Africa. Asian J. Plant Sci. 6:732-740. |
|
|
Birdsey MR (1951). The cultivated aroids. Gillick Press, Berkeley, CA. |
|
|
Deng Z, Harbaugh BK (2006a). 'Garden White'-A large white fancy-leaved Caladium for sunny landscapes and large containers. HortScience 41:840-842 |
|
|
Deng Z, Harbaugh BK (2006b). Independent inheritance of leaf shape and main vein color in caladium. J. Am. Soc. Hortic. Sci. 131:53-58. |
|
|
Deng Z, Harbaugh BK, Kelly RO, Seijo T, McGovern RJ (2005). Pythium root rot resistance in commercial caladium cultivars. HortScience 40:549-552. |
|
|
Ekeke C, Mensah SI (2015). Comparative Anatomy of Midrib and its Significance in the Taxonomy of the Family Asteraceae from Nigeria. J. Plant Sci. 10(5):200-205. |
|
|
Essiett UA, Umoh NU (2014). Studies of the leaf and floral anatomy of two species of Ixora. Int. J. Med. Plants Altern. Med. 2(2):13-20. |
|
|
Evans MR, Wilfret GJ, Harbaugh BK (1992). Caladiums as potted and landscape plants. Florida Coop. Ext. Services, Univ. Florida, Inst. Food Agr. Sci. 30 Apr. 2007. |
|
|
Gary GC (2009). Diversity and distribution of idioblasts producing calcium oxalate crystals in Dieffenbachia seguine (Araceae). Am. J. Bot. 96(7):1245-1254. |
|
|
Goncalves EG, Paiva EAS, Nadruz Coelho MA (2004). A preliminary survey of petiolar collenchyma in the Araceae. Ann. Mo. Bot. Gard. 91:473-484. |
|
|
Hayward W (1950). Fancy-leaved caladiums. Plant Life 6:131-142. |
|
|
Hesse M (2006a). Pollen wall ultrastructure of Araceae and Lemnaceae in relation to molecular classifications. Aliso 22:204-208. |
|
|
Hesse M (2006b). Reasons and consequences of the lack of a sporopollenin ektexine in Aroideae (Araceae). Flora 201:421-428. |
|
|
Hutchinson J, Dalziel JM (1954). Flora of West Tropical Africa. Crown Agents, London UK. |
|
|
Jin PL, Ruth K, Andrea K, Leong HG, Yik-Yuen G (1999). Amplified Fragment Length Polymorphism (AFLP) Provides Molecular Markers for the Identification of Caladium bicolor Cultivars. Ann. Bot. 84:155-161. |
|
|
Keating RC (2000). Collenchyma in Araceae: Trends and relation to classification. Bot. J. Linn. Soc. 134:203-214. |
|
|
Keating RC (2002). Acoraceae and Araceae. In. Gregory M. and Cutler D.F. [eds.], Anatomy of the monocotyledons, vol. 9. Oxford University Press, Oxford, U.K. |
|
|
Keating RC (2004). Systematic occurrence of raphide crystals in Araceae. Ann. Mo. Bot. Gard. 91:495-504. |
|
|
Kemka-Evans CI, Okoli BE, Nwachukwu C (2014). Epidermal studies of three species of Vernonia Schreb. in Southern Nigeria. Biodiversitas 15:137-141. |
|
|
Madison M (1981). Notes on Caladium (Araceae) and its allies. Selbyana 5:342-377. |
|
|
Malvey P (2004). Structure, nomenclature and classification of stomata. Acta Bot. Sin. 44(2):242-252. |
|
|
Mayo SJ, Bogner J, Boyce PC (1997). The genera of Araceae. The Trustees, Royal Botanic Gardens, Kew, U.K. |
|
|
Metcalfe CR, Chalk L (1968). Current development in Systematic Plant Anatomy, In. Modern methods in Plant Taxonomy (V.H. Heywood.). Academy press London, New York. pp. 45-47. |
|
|
Metcalfe CR, Chalk L (1979). Anatomy of the dicotyledon, vol. 1: systematic anatomy of the leaf and stem. Oxford University Press, New York. |
|
|
Natalie C, Josef B, Simon JM, Peter CB, Sin YW, Michael H, Wilbert LAH, Richard CK, Jim CF (2011). Relationships within the Araceae : Comparison of Morphological patterns with Molecular phylogenies. Am. J. Bot. 98(4):654-668. |
|
|
Ndukwu BC, Agbagwa IO (2006). The value of leaf micromorphological characters in the taxonomic delimitation of Emilia cass. (Asteraceae) species. Glob. J. Pure Appl. Sci. 12(2):183-187 |
|
|
Nurul C, Nunung H, Retno M (2013). Variation of Calcium Oxalate (CaOx) Crystals in Porang (Amorphophallus muelleri Blume). Am. J. Plant Sci. 4:1765-1773. |
|
|
Nyananyo BL, Osuji JO (2007). Biosystematic investigation into Sphenostylis stenocarpa (Hochst ex A. Rich) Harms (Fabaceae) in Nigeria. Niger. J. Bot. 20(2):411-419. |
|
|
Okoli BE (1988). On the probable function and taxonomic value of calcium oxalate crystals in Cucurbitaceae. Feddes Repert. 99:139-142. |
|
|
Okoli BE, Green BO (1987). Histochemical localization of calcium oxalate crystals in starch grains of yams (Dioscorea). Ann. Bot. 60:391-394. |
|
|
Okoli BE, McEuen AR (1986). Calcium-containing crystals in Telfairia Hooker (Cucurbitaceae). New Phytol. 102:199-207. |
|
|
Okoli BE, Ndukwu BC (1992). Studies on Nigerian Curcurbita moschata. Niger. J. Bot. 5:18-26. |
|
|
Osuji JO, Nwala PC (2015). Epidermal and Cytological Studies on Cultivars of Xanthosoma (L.) Schott. and Colocasia (L.) Schott. (Araceae). Int. J. Plant Soil Sci. 4(2):149-155. |
|
|
Osuji JO (2013). Probable functions of calcium oxalate crystals in different tissues of the edible aroids (Xanthosoma and Colocasia spp.) in Nigeria. Afr. J. Biotechnol. 12(25):3952-3956. |
|
|
Mais Sk, Amal KM (2012). Studies on the calcium oxalate crystals (Raphides) and idioblast of some selected members of Araceae in Eastern India. Afr. J. Plant Sci. 6(9):256-269. |
|
|
Uno G, Storey R, Moore R (2001). Principles of Botany. McGraw Hill Companies Inc. Boston Burr Ridge, New York, London. P 552. |
|
|
Wang W, Zhao N (2002). Epidermal Characters of Leaves in Araceae. Plant Sci. J. 20(5):343-349. |
|
|
Wilfret GJ (1993). Caladium. In: de Hertogh A, le Nard M, eds. The physiology of flower bulbs. New York: Elsevier, pp. 239-247. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


