ABSTRACT
Breeding for resistant varieties has been shown to be the most suitable method to control bean anthracnose caused by Colletotrichum lindemuthianum though the method is challenged by the existence of many races of the pathogen. This work focused on characterizing races of C. lindemuthianum from potential bean agro ecological zones of Tanzania using a set of differential bean cultivars. From 144 anthracnose infected bean samples collected, 50 pure isolates were obtained and characterized whereby 42 races were identified. The most virulent race identified was race 3610 from the Southern Highland zone of Tanzania while the least virulent was characterized as race 0. Race 2 was the most widely spread (4.2 %) found in Northern, Southern highland and in Eastern zones of Tanzania. The work confirms that G2333 can still be used as a potential donor of resistant genes to varieties that are to be grown in Northern, Eastern and Lake zones but not for the varieties from Southern Highland and Western zones of Tanzania since isolates from these zones overcame resistant genes Co-42, Co-5, Co-7 in G2333. 95% of all races which were identified are new and were not specific to either Mesoamerican or Andean origin common bean.
Key words: Colletotrichum lindemuthianum, races, resistant genes, differential cultivars, Anthracnose.
Bean anthracnose is one of the most serious diseases of common bean caused by the fungal pathogen Colletotrichum lindemuthianum (Sacc. & Magnus) Briosi & Cavara in areas where common bean is grown. The disease has been reported to cause losses of up to 100% if not well controlled (Opio et al., 2001; Markell et al., 2012). In Tanzania, the work of Allen et al. (1998) in Northern Tanzania reported that, for each 1% increase in anthracnose incidence, seed yield decreases by 9 kg/ha while Shao and Teri (1985) and Mwalyego (1991) reported loss due to anthracnose to be up to 100% if the disease is not well controlled. According to Opio et al. (2001), the effective control measures to reduce losses caused by bean anthracnose is the use of resistant varieties. This has been proven to be cost effective and requires less skill for farmers to apply it (Mahuku et al.,
2002; Abraham and Mashilla, 2018).
Several genes from both Meso-Americans and Andean bean origin have been shown to offer resistance to anthracnose disease of common bean to anthracnose disease (Méndez-Vigo et al., 2005). The Mesoamerican genes include Co-2, Co-3 (and its alleles Co-32, Co-33, Co-34, and Co-35), Co-4 (and it alleles Co-42, Co-43), Co-5 (and its allele Co-52), Co-6, Co-11, Co-16, Co-17, Co-u, and Co-v while the Andean genes include: Co-1 (and its alleles Co-12, Co-13, Co-14, and Co-15), Co-12, Co-13, Co-14, Co-15, Co-x, Co-w, Co-y, and Co-z (Kelly and Vallejo, 2004; de Lima Castro et al., 2017). But the effective choice and introgression of resistant genes to the host (common bean) in order to develop a resistant variety depends on the knowledge of the variability of C. lindemuthianum.
Mahuku and Riascos (2004), Kelly and Vallejo (2004), Silva et al. (2007), and Vidigal et al. (2007) reported that C. lindemuthianum has a high rate of evolution resulting in emergence of the new race of the pathogen from time to time. This causes resistance breakdown for the already developed resistant varieties because the resistance of common bean to C. lindemuthianum is very specific, so that a resistance gene from a bean genotype confers protection against specific isolates or races of C. lindemuthianum.Therefore, before breeding for anthracnose resistant varieties, it is important to study and identify the available races of C. lindemuthianum which is helpful in planning suitable breeding strategies for varieties with durable resistance and to design effective gene pyramids and deployment of resistant cultivars over time and space.
In Tanzania, although some of the works to develop resistant varieties to bean anthracnose have been done, information on C. lindemuthianum variability and the race identification is still limited for effective breeding for the anthracnose disease. Therefore, the present work focused on studying the variability of C. lindemuthianum in major bean growing regions of Tanzania and identifying races of C. lindemuthianum available in such areas using differential bean cultivars as proposed by Pastor-Corrales (1991).
Collection of bean sample infected with anthracnose
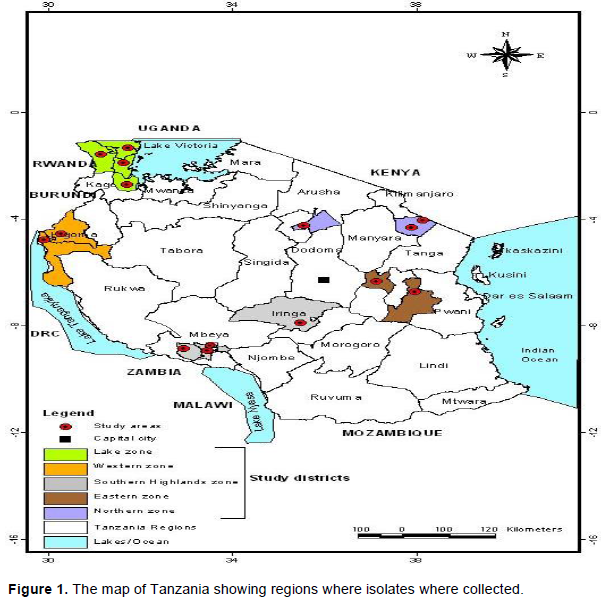
Collection of common bean samples infected with anthracnose disease was done in five bean growing zones in Tanzania (Figure 1) during 2016/2017 bean growing season. Selection of zone was based on potential common bean production. In each zone, the distance between sampling fields was separated by a distance of 2 to 5 km. A minimum of five fields were sampled per zone. In the field, quadrants of 3 m x 3 m were used to randomly collect anthracnose symptomatic bean plants. A total number of four subplots identified by quadrants per field were established and, in each subplot four to nine plants were sampled. The collected infected plant parts were pods, leaves and stems. The pod samples collected were pressed well and stored in the herbarium, while other bean pods of the same plant were preserved in silica gel. Leaves and stems were pressed well in the herbarium before they were taken to the plant pathology laboratory at Sokoine University of Agriculture, Morogoro for isolation and characterization of C. lindemuthianum.
Pathogen isolation and inoculum preparation
Pathogen isolation was performed from the well-developed anthracnose diseased bean pods and leaves. Selected samples were sterilized for two minutes using 2% sodium hypochlorite (NaOCl) solution and rinsed well using sterile distilled water. Small cuts of 5cm long were made on the symptomatic areas and the cut placed on the prepared V8 growth media (17 g micro agar, 3 g of calcium carbonate, 200 ml of V8 juice and 800 ml of distilled water mixed together and autoclaved at 120°C and 2 bars of pressure for 15 min). The culture media were then incubated in the dark room at 22 to 24°C for seven days. From well grown isolates, mono-conidial isolates were obtained by picking pin-point colonies from the media using a sterile needle, and then sub-cultured on fresh V8 medium for purification of cultures using the single spore technique; and then single spore isolates were incubated aseptically. The standard spore suspensions for each obtained fungal isolate were prepared following the procedure of (Pastor-Corrales et al., 1995) and were stored at -4°C before inoculation to the differential bean cultivar.
Raising differential cultivars and inoculation
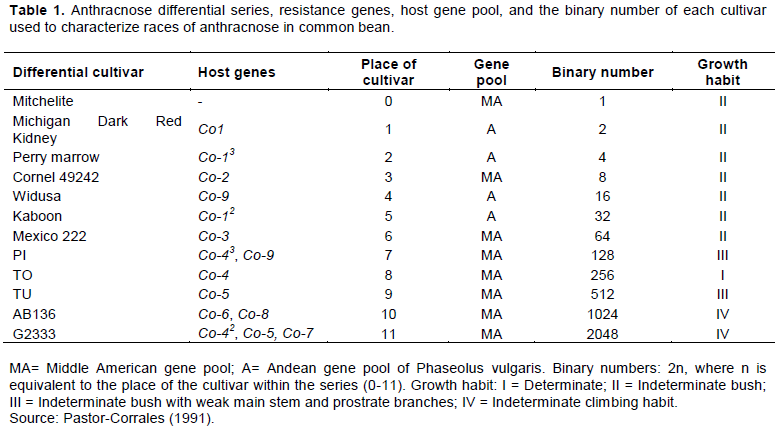
A set of differential cultivars as described by Pastor-Corrales (1991) (Table 1) obtained from the Bean Improvement Program at Sokoine University of Agriculture (SUA) were planted in pots in the screen house at the Department of Crop Science and Horticulture, SUA. At fourteenth days after sowing, each differential cultivar seedling was inoculated separately by standard C. lindemuthianum inoculum prepared from the obtained isolate. The spore concentration used were quantified and adjusted to spore concentrations of 1.2×106 spores/ml (Pastor-Corrales et al., 1995) using the hemocytometer (Neubauer-Preciss Chamber) and the light microscope. The inoculation was done using the hand sprayer and the experiment was replicated three times.
In order to maintain the relative humidity of approximately 95% for the infection to take place, each plant was covered by the transparent polythene sheets for four days after inoculation followed by regular management such as watering and fertilizer application.
Data collection and data analysis
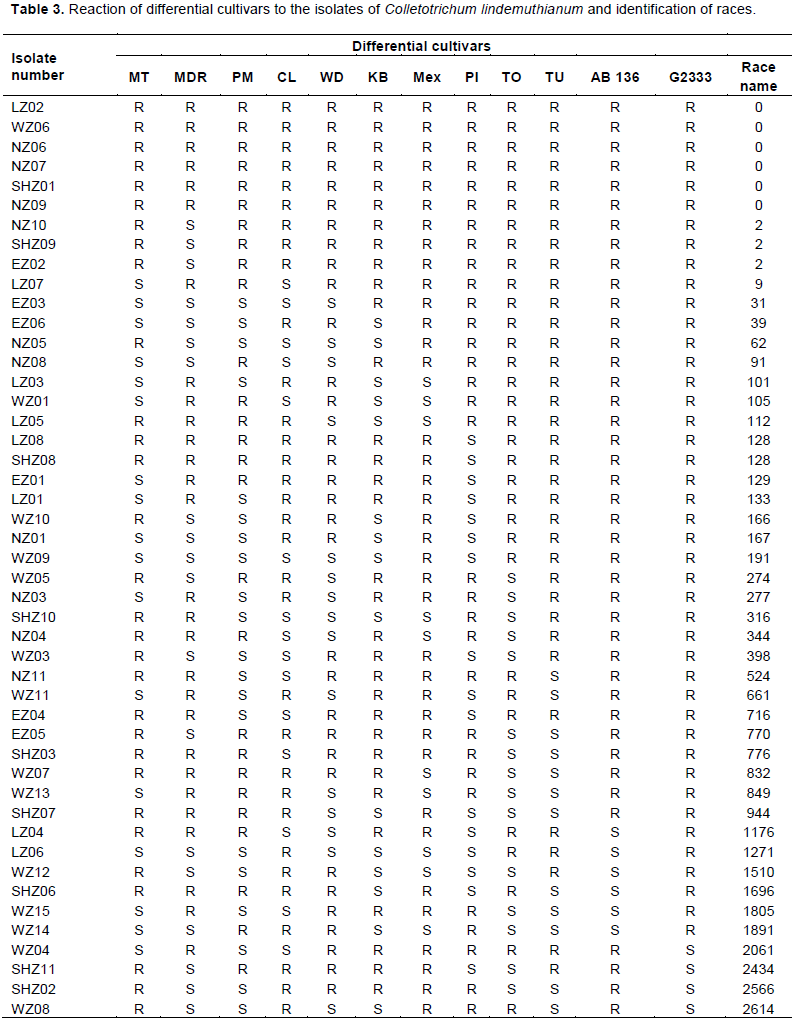
Symptom evaluation and scoring of anthracnose disease severity on inoculated bean plants was done eight days after inoculation using a scale of 1-9 previously used by Balardin et al. (1990). The average disease scores were obtained and the disease reaction type categories were determined. Cultivars with scores of 1-3 were regarded as resistant while cultivars with scores of 4-9 were regarded as susceptible (Barladin et al., 1990). Race identification was done as the sum of binary numbers of all differential cultivars showing susceptible reaction to the particular isolate.
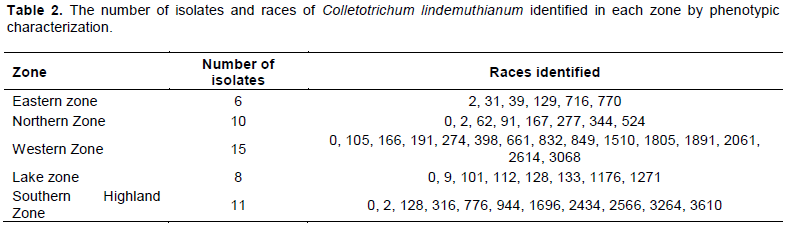
Phenotypic characterization of C. lindemuthianum done by using set of differential cultivars as proposed by Pastor-Corrales (1991) was able to identify 50 isolates from 144 bean samples collected. Of these 50 isolates obtained, 15 were from Western zone, 8 from Lake zone, 10 from Northern zone, 11 from Southern highland zone and 6 isolates were from Eastern zone (Table 2). Further it was possible to identify 42 races out of 50 isolates obtained (Tables 2 and 3). The most virulent race was race 3610 which was collected from the Southern Highland zone while the least virulent isolate was characterized as race 0, found in the Northern, Lake, Western and Southern Highland zones (Tables 2 and 3). From the Eastern zone, six isolates of C. lindemuthianum were identified out of all samples which were collected (Table 2). No isolate from the Eastern zone was able to infect the differential cultivar G2333 while differential cultivar Michigan Dark Red Kidney was the most affected differential cultivar with the four isolates. From the Eastern zone, the least virulent isolate was race 2 while the most virulent was characterized as race 770 which affected the differential cultivars Michigan Dark Red, TO, TU and AB136 (Table 3). From the Northern bean growing zone of Tanzania, among a total of 10 isolates collected, 8 races of C. lindemuthianum isolates were characterized (Table 2). These isolates affected almost all the differential cultivars except AB136 and G2333. Differential cultivars Michigan dark red kidney, Perry Marrow, Cornell and Widusa were highly infected with four isolates from the northern zone. The most virulent isolate was characterized as race 524 while the least was characterized as race 0 (Table 3). From the western zone, 15 isolates were obtained (Table 2). All differential cultivars were affected by these isolates including the most resistant differential cultivar G2333, which was affected by two isolates while Perry Marrow was the most affected differential cultivar. From this zone, the most virulent race was race 3068 while the least virulent race was race 0 (Table 3).
A total of eight isolates of C. lindemuthianum were obtained from the Lake zone (Table 2). These isolates could not affect differential cultivars G2333, TO and TU while Michelite was the most affected differential cultivar by four isolates (Table 3). The most virulent isolate from the Lake zone was characterized as race 1271 while the least virulent isolate from this zone was characterized as race 05 (Tables 2 and 3). From the Southern highland zone, 11 isolates were obtained in which (Table 2). Michelite was the only differential cultivar not affected by any isolate from this zone; while TU was the most affected differential cultivar affected by four isolates (Table 3). Isolates from this zone were able to infect the differential bean cultivar G2333 (Table 3). The most virulent isolate from the Southern Highland zone was characterized as race 3610 which infected differential bean cultivars Widusa, TU, AB136 and G2333; while the least virulent isolate was characterized as race 0 (Table 3).
Among the differential bean cultivars used, Perry Marrow was the most susceptible differential cultivar and it was infected by 42% of all C. lindemuthianum isolates collected while differential bean cultivar G2333 was the most resistant cultivar to most of the collected isolates; it was affected by only 14 % of isolates collected, and these were from the Western and Southern Highland zones (Tables 2 and 3).
Of all the races identified in bean growing zones of Tanzania, race 2 was the most common with high frequency of 4.8 % and widely distributed within bean growing zones; and it was distributed in Northern, Southern Highland and in Eastern zones, while race 128 was only distributed in Lake and Southern Highland zones. Race 3610 collected from the Southern Highland zone was the most virulent isolate compared to all isolates collected from bean growing zones while the least virulent race identified was characterized as race 0.
The phenotypic characterization of C. lindemuthianum in major bean growing areas in Tanzania revealed the existence of races and the variability of C. lindemuthianum. This study reveals new races of C. lindemuthianum which were not reported before. Earlier studies by Ansari et al. (2004) identified five races (0, 2, 12, 38 and 192) of C. lindemuthianumin Tanzania whereby only races 0 and 2 were also reported by this work distributed in all five zones.
This study confirms the evolution of C. lindemuthianum with time in Tanzania. The work of Mwalyego (1991) identified races 6, 28, 60, 63, 98, 155, 182, 287, 618, 958, 1478, 1515 and 1678 whereby out of them no races were reported in this study.
In this study 96% of the races identified from the collected isolates have not been reported before in Tanzania. The previous work done by Ansari et al. (2004) to study the variability of C. lindemuthianum of isolates from Tanzania who also identified five races (0, 2, 12, 38 and 192) out of 8 isolates which were characterized and race 0 and 2 were common races which were also identified from this work. This could be due to the extensive collection of isolates which was done during this study; it covers large area growing common bean. Some new races could have evolved from when the last study was conducted in the country till now when this study was conducted. Most of the virulent isolates were found in areas where the weather condition favors the growth and development of the pathogen (Mohamed, 2013). For example, the most virulent isolate characterized as race 3610 was found in the Southern Highland zone. This zone was characterized by very cool and humid conditions to most of its areas during the bean growing season (Tanzania Meteorological Agency, 2017). Such conditions favor growth and developments of C. lindemuthianum which may influence its variability as it have been reported by Tu (1983) and Mohammed (2013).
This study shows that C. lindemuthianum in Tanzania is not specialized to either Meso-American or Andean common bean gene pool only. The collected isolates were able to affect differential bean cultivars of both Meso-American and Andean gene pool. For example isolate SHZ05 from the Southern Highland zone was able to overcome both Andean resistance gene Co-9 in Widusa and Meso-American resistance genes (Co-42, Co-5, Co-7) in G2333, Co-6 and Co-8 in AB136 and Co-5 in TU. Similar work of Balardin et al. (1990) and Balardin et al. (1997) indicated that some races contain virulence factors to both Meso-American and Andean gene pools and they infect both Andean and Meso-American bean gene pools. In the areas with large host diversity of both Mesoamerican and Andean at the same time it highly influences the pathogen diversity such as isolates collected from Western zone. Therefore, in order to develop the durable resistance variety, it is important to incorporate more than one resistant gene by gene pyramiding technique as it was also proposed by Balardin and Kelly (1998), Kelly and Vallejo (2004), Pastor-Corrales et al. (1995) from both Meso-American and Andean sources so as to provide the broader range and durable resistance variety.
Out of 12 differential cultivars used, G2333 was the most resistant differential bean cultivar affected by only 7 isolates from the Western and Southern Highland zones only. Previous work reported that the presence of three resistant genes (Co-42, Co-5 and Co-7) in G2333 offer broader resistance to a wide range of C. lindemuthianum races (Young et al., 1998; Mahuku et al., 2002). Therefore based on races identified in Northern, Eastern and the Lake zones, G2333 can be used as a potential donor for resistance genes in bean breeding program for the varieties to be grown in those areas, according to Mwalyego (1992a) who reported 15 separate isolates and none of them was pathogenic to G2333 bean cultivar. But varieties that are to be grown in the Southern Highland and Western zones should be incorporated with genes for resistance from other sources. de Lima Castro et al. (2017) reported Paloma cultivar as a new source of resistance gene to common bean anthracnose. This cultivar has a new dominant gene conferring resistance to anthracnose, which is independent of Co-1, Co-2, Co-34, Co-4, Co-42, Co-43, Co-5, Co-6, Co-12, Co-13, Co-14, Co-15 and Co-16 resistance genes identified previously; this gene is named Co-Pa. Therefore, in the future Paloma cultivar can be included in the list of differential cultivars before being recommended as the donor source of resistant genes to the cultivars grown in the Western and Southern Highland zone. Also more characterization work should be done on (Co-Pa) resistant genes in Paloma.
This study indicates that the pathogen, C. lindemuthianum which causes anthracnose disease, C. lindemuthianum shows variability in major bean growing areas of Tanzania. The pathogen seems to be widely distributed in all bean growing areas with the most virulent isolate from the Southern Highland zone. The best control method for the disease in these potential bean growing areas is using the host resistance. It is recommended to use Co-42, Co-5 and Co-7 from G2333 to develop resistant varieties to be grown in the all zones except in Southern and Western zone. The research should also focus on characterizing other genes that can offer resistance to races from Western and Southern Highland zones that were able to overcome resistance genes in G2333 such as Co-Pa in cultivar Paloma.
The authors have not declared any conflict of interests.
REFERENCES
|
Abraham N, Mashilla D (2018). Effect of integrating variety, seed treatment, and foliar fungicide spray timing on managing common bean Anthracnose at Bako, western Ethiopia. East Africa Journal of Science 12(2).
|
|
|
|
Allen DJ, Burachara RA, Smithson JB (1998). Diseases of common bean. In. Allen DJ, Lenne JM (Eds.), The Pathology of Food and Pasture Legumes. CAB International: Wallingford, UK, pp. 179-265.
|
|
|
|
|
Ansari KI, Palacios N, Araya C, Langin T, Egan D, Doohan FM (2004). Pathogenic and genetic variability among Colletotrichum lindemuthianum isolates of different geographic origins. Journal of Plant Pathology 53:635-642.
Crossref
|
|
|
|
|
Balardin RS, Kelly JD (1998). Interaction between Colletotrichum lindemuthianumraces and gene pool diversity in Phaseolus vulgaris. Journal of American Society 123:1038-1047.
Crossref
|
|
|
|
|
Balardin RS, Jarosz AM, Kelly JD (1997). Virulence and molecular diversity in Colletotrichum lindemuthianum from South, Central and North America. Phytopathology 87:1184-1191
Crossref
|
|
|
|
|
Balardin RS, Pastor-Corrales MA, Otoya MM (1990). Variabilida de patogênica de Colletotrichum lindemuthianum no Estado de Santa Catarina. Fitopatologia. Brasilia 15:243-245.
|
|
|
|
|
de Lima Castro SA, Gonçalves-Vidigal MC, Gilio TA, Lacanallo GF, Valentini G, Martins VD, Song Q, Galván MZ, Hurtado-Gonzales OP, Pastor-Corrales MA (2017). Genetics and mapping of a new anthracnose resistance locus in Andean common bean Paloma. BioMed Central Genomics 18(306):1-13.
Crossref
|
|
|
|
|
Kelly JD, Vallejo VA (2004). A comprehensive review of the major genes conditioning resistance to anthracnose in common bean: Crop and soil sciences. Journal of Horticultural Science 39(6):1196-1207.
Crossref
|
|
|
|
|
Mahuku GS, Jara CE, Cajiao C, Beebe S (2002). Sources of resistance to Colletotrichum lindemuthianum in the secondary gene pool of Phaseolus vulgaris and in crosses of primary and secondary gene pools. Plant Disease 86:1383-1387.
Crossref
|
|
|
|
|
Mahuku SG, Riascos JJ (2004). Virulence and molecular diversity within Colletotrichum lindemuthianum isolates from Andean and Mesoamerican bean varieties and regions. European Journal of Plant Pathology 110:253-263.
Crossref
|
|
|
|
|
Markell S, Wunsch M, Del Río L (2012). Anthracnose of Dry Beans. State University Extension Publication, Fargo. 33p.
|
|
|
|
|
Méndez-Vigo B, Rodriguez-Suarez C, Pañeda A, Ferreira J, Giraldez R (2005). Molecular markers and allelic relationships of anthracnose resistance gene cluster B4 in common bean. Euphytica 141:237-245.
Crossref
|
|
|
|
|
Mohammed A (2013). An Overview of Distribution, Biology and the Management of Common Bean Anthracnose. Journal of Plant Pathology and Microbiology 4:193-199.
Crossref
|
|
|
|
|
Mwalyego FM (1991). Progress in bean anthracnose research in Tanzania. In. Buruchara R (eds.), Proceedings of the First Pan-African Working Group Meeting on Anthracnose of Beans, Ambo, Ethiopia African Workshop. Centro Internacional de Agricultura Tropical, Cali Colombia. pp. 61-65.
|
|
|
|
|
Mwalyego F (1992a) Progress report on studies of the bean anthracnose pathogen (Colletotnchum lmdemuthianum) in Tanzania. Unpublished report Uyole Agncultural Centre Mbeya Tanzania Cited in Allen DJ, CIAT African Workshop Senes No 27 211 236.
|
|
|
|
|
Opio F, Ugen MA, Kyamanywa S, David S, Mugisa-Mutetika M (2001). Beans. In. Mukiibi K (ed.), Agriculture in Uganda Crop. (National Agricultural Research Organization, Uganda, pp. 56- 72.
|
|
|
|
|
Pastor-Corrales MA (1991). Standardization of differential varieties and designation of races of Colletotrichum lindemuthianum. Phytopatholoy 81(6):694.
|
|
|
|
|
Pastor-Corrales MA, Otoya MM, Molina A, Singh SP (1995). Resistance to Colletotrichum lindemuthianum isolates from Middle America and Andean South America in different common bean races. Plant Disease 79:63-67.
Crossref
|
|
|
|
|
Shao FM, Teri JM (1985). Yield losses in Phaseolus beans induced by anthracnose. International Journal of Pest Management 31(1):60-62.
Crossref
|
|
|
|
|
Silva KJD, Souza EA, Ishikawa FH (2007). Characterization of Colletotrichum lindemuthianum isolates from the state of Minas Gerais, Brazil. Journal of Phytopathology 155:241-247.
Crossref
|
|
|
|
|
Tanzania Meteorological Agency (2017). Tanzania weather condition.
View site visited on 4/9/2018.
|
|
|
|
|
Tu JC (1983). Epidemiology of anthracnose caused by Colletotrichum lindemuthianum on white bean (Phaseolus vulgaris) in Southern Ontario: Survival of the pathogen. Plant Disease 67:402-404.
Crossref
|
|
|
|
|
Vidigal FPS, Goncalves-Vidigal MC, Kelly JD, Kirk WW (2007). Sources of resistance to anthracnose in traditional common bean cultivars from Parana, Brazil. Journal of Phytopathology 155:108-113
Crossref
|
|
|
|
|
Young R, Melotto A, Nodari MRO, Kelly JD (1998). Marker assisted dissection of oligogenic anthracnose resistance in the common bean cultivar. Theoretical and Applied Genetics 96:87-89.
Crossref
|
|