Full Length Research Paper

ABSTRACT
A greenhouse research was conducted at the research area of Department of Plant Breeding and Genetics, University of Agriculture, Faisalabad to evaluate 20 accessions for the identification of salt tolerant genotypes of sunflower (Helianthus annuus L.) as well as their characteristics. The experiment was conducted in completely randomized design with three repeats. Salinity was developed with NaCl to achieve the final levels of 3, 6 and 9 dsm-1 salinity, whereas control contained tap water. After 60 days of planting, 10 seedlings of each accession from each treatment and replication were uprooted and data was recorded. Sunflower genotypes G-36, G-61, A-23, A-61 and A-185 performed better in both controlled and saline conditions. These genotypes showed better shoot and root growth and biomass by least concentration of Na+ and higher concentration of Cl- in leaf sap.
Key words: Tap water, NaCl, replication, leaf sap.
INTRODUCTION
Sunflower (Helianthus annuus L.) is the second most important oilseed crop after soybean worldwide (Paniego et al., 2002). Sunflower is high yielding and non-conventional oilseed crop. It had desirable traits that is high oil contents (40-47%), protein (23%), high linoleic acid, toxic free elements and contain vitamins A, D, E, K. Salinity in soil or water is one of the major stresses and especially in arid and semi arid regions, can severely limit crop production. Salinity impairs seed germination, reduces nodule formation, retards plant development and reduces crop yield. High levels of soil salinity negatively affect productivity of most field crops (Munns, 1993). Saline soils remarkably reduce oil production potential and oil yield of sunflower (Szabolcs, 1994).The plants that grow in saline soils have diverse ionic compositions and a range in concentrations of dissolved salts. These concentrations fluctuate because of changes in water source, drainage, evapotranspiration, and solute availability. About 7% of arable lands of the world are under salinity pressure (Jumsoon et al., 1996). Soil salinity reduces water availability of plant roots via negative (low) osmosis potential, as well as decrease of germination dynamics of plant seeds by ionic toxicity of behavior and response of different accessions of sunflower to tolerate salt stress at seedling stage. The Na+ and Cl- (Munns et al., 1988).
The study was conducted to understand the genetic information so obtained will be useful in formulating criteria for salt stress tolerance and high yield. The objective was also the development of selection criteria through correlation and path analysis studies. The selected types could be used in hybridization programme aimed at breeding for sunflower yielding high under salt stress conditions. The availability of high yielding salt stress tolerant sunflower is perceived to attract farmer to use the land resources otherwise left fallow due to salt stress.
MATERIALS AND METHODS
The present study was carried out under the glass house of the department of Plant Breeding and Genetics, University of Agriculture, Faisalabad. The research material was comprised of 20 accessions of sunflower developed by the Oilseed Research Programme of the Department. These accessions (G-16, G-30, G-32, G-36, G-44, G-45, G-61, G-64, G-66, G-68, G-86, A-2, A-14, A-23, A-56, A-60, A-61, A-79, A-133 and A-185) were planted and evaluated for various traits in the salinity experiments.
Experimental layout
Experiment was conducted in a glass house with no control of humidity, temperature and light. The experiment was laid out following factorial complete randomized design in three replications. The sunflower seed were planted in iron trays. Each tray was filled with soil and sand in the ratio of 1:1. The seeds were sown at the depth of 1.5 cm by maintaining distance of 2.5 cm each for row to row and seed to seed.
Planting medium
Normal soil free from any salinity and sodicity hazards was collected from the research area of department of Plant Breeding and Genetics, University of Agriculture Faisalabad. The mixture of sand and soil was air dried, ground and passed through 2 mm sieve and analyzed for chemical characters (Table 1).

Treatments
Tap water was applied for irrigation for 15 days according to requirement. After germination, four salt (NaCl) levels of irrigation water were maintained: treatment 1 = normal water (tap water); treatment 2 = 3 dsm-1; treatment 3 = 6 dsm-1; Treatment 4 = 9 dsm-1. Composition of tap water is given in Table 2.

Tissue sap analysis for ion uptake
Sample collections
The 10 randomly selected plants per replication and per treatment for each genotype were uprooted. Two lower leaves (form the basal node) and two upper leaves (from the top node) were collected, washed with tap water to remove the soil residues and then dipped instantly in distilled water for a short period of time. The samples were blotted dry with the help of a sheet of blotting paper, placed in polyethylene bags, marked with the spirit marker and stored in the deep freezer for tissue sap extraction.
Extraction of leaf sap
Frozen leaf samples were thawed, after washing with distilled water, the tissue sap was extracted by using metal rod .The tissue sap oozing out from the samples was collected in epindroph tubes and immediately stored back in the deep freezer.
Centrifugation of tissue sap
The epindroph tubes were taken out from the freezer and placed at the room temperature to thaw. Then the thawed tissue sap was centrifuged at the 6500 rpm for 5 min. The supernatant tissue sap samples from epindroph tubes were analyzed for chloride, sodium and potassium ions.
Determination of chloride ions
Chloride ions in the tissue sap was determined by chloride analyzer (Sherwood chloride analyzer 926).
Determination of sodium ions
The tissue sap was diluted as required with distilled water. Sodium ions were determined using Flame Photometer (Sherwood flame photometer 410).
Data recording
After 60 days of planting, ten seedlings of each accession from each treatment and replication were uprooted. Data were recorded from the experiment on following parameters viz. Germination percentage (G %), emergence index (EI), emergence rate index (ERI), chlorophyll content, root: shoot ratio, mortality (M %).
RESULTS AND DISUCSSION
The experimental results were obtained, presented and discussed separately.
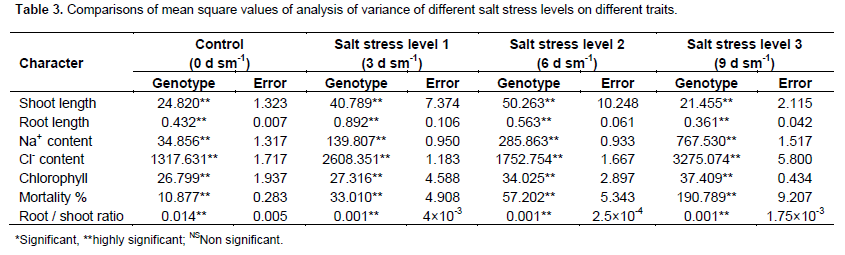
Analysis of variance under four treatments
Under salt stress condition level 2 (6 dS m-1) and level 3 (9 dS m-1), significant differences and marked variation among accessions for all the traits were detected under normal and all salt stress levels, furthermore, the genotypes behaved differently to the stress. The comparison of treatment expression of various plant traits under normal and salt stress level 3 (9 dS m-1) conditions suggested that salt stress adversely affected for the characters (Table 3).
Germination percentage (G%)
The analysis of variance of germination percentage of sunflower population under the study (Table 3). Non significant differences existed among of genotypes of the sunflower.
Emergence index (EI)
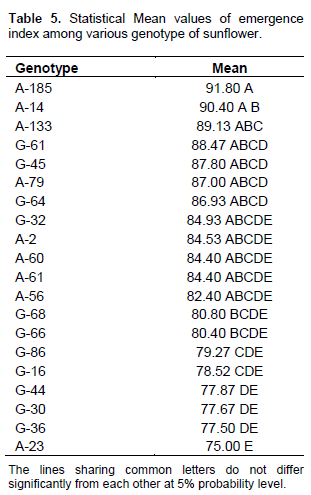
The analysis of variance of sunflower genotypes for emergence index is shown in Table 4. The results indicate that sunflower genotypes had significant differences and ranged from 91.80 to 75.00 (Table 5). The maximum value 91.80 was observed in the genotype A-85 followed by the genotype A-14 (90.40) and genotype A-133 (89.13).

Emergence rate index (ERI)
Table 4 reveals that the differences for emergence rate index among genotypes were non-significant.
Effect of salt stress (NaCl) on shoot length (cm)
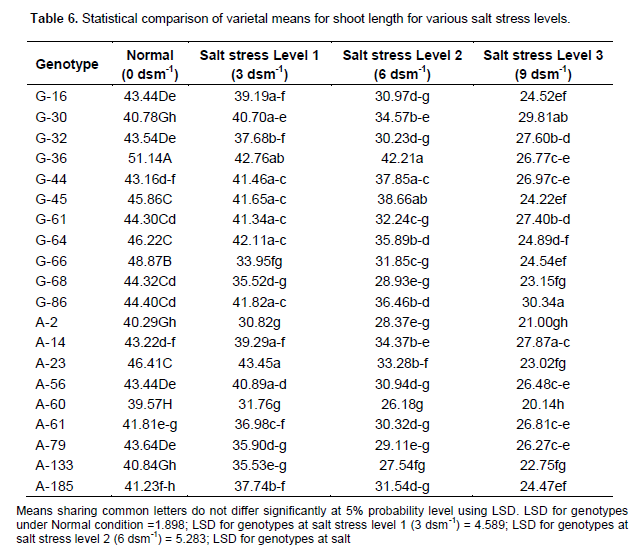
Table 6 indicates that the shoot length was decreased significantly with increasing salinity levels. Interaction of all the genotypes and treatments was found significant.
Table 6 shows that accession G-36 closely followed by G-66 and A-23 had maximum shoot length and accession A-60 followed by A-2 and G-30 had minimum shoot length under normal condition. Accession A-23 closely followed by G-36 and G-64 had maximum shoot length under salt stress level 1 (3 dsm-1). Accessions G-36 closely followed by G-45 and G-44 had maximum shoot length under salt stress level 2 (6 dsm -1). Accession G-86 closely followed by G-30 and A-14 had maximum shoot length under salt stress level 3 (9 dsm-1). Ramoliya and Panday (2003); Mer et al. (2000) and Ramdiya and Panday (2003) also demonstrated that salinity in nutrient solution reduced the growth of black spot (Diospyros digvna Jacq).
Effect of salt stress (NaCl) on root length (cm)
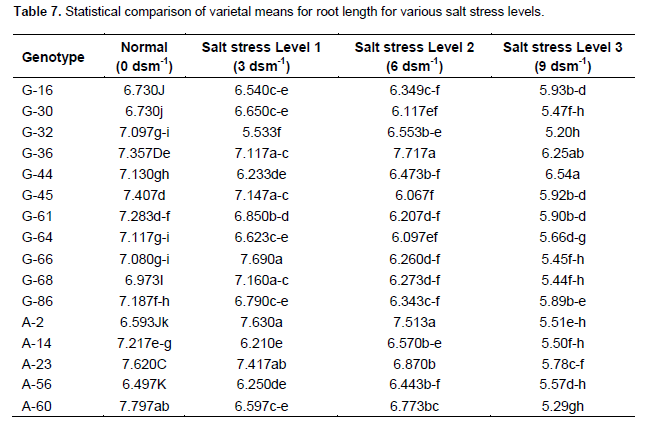
Table 7 shows that accession A-133 closely followed by A-60 and A-185 had maximum root length under normal condition. Accession G-66 closely followed by A-2 and A-79 had maximum root length under salt stress level 1 (3 dsm-1). Accessions G-36 closely followed by A-2 and A-23 had maximum root length under salt stress level 2 (6 dsm -1). Accession G-44 closely followed by G-36 and A-185 had maximum root length under salt stress level 3 (9 dsm-1). Qureshi et al. (1998), Hussain and Rehman (1995) and Ghumman (2000) also conducted experiments on sunflower and found that root length and relative root length decreased with increase in salinity.
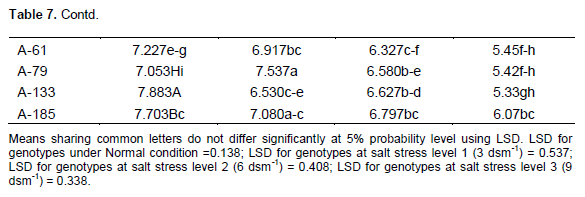
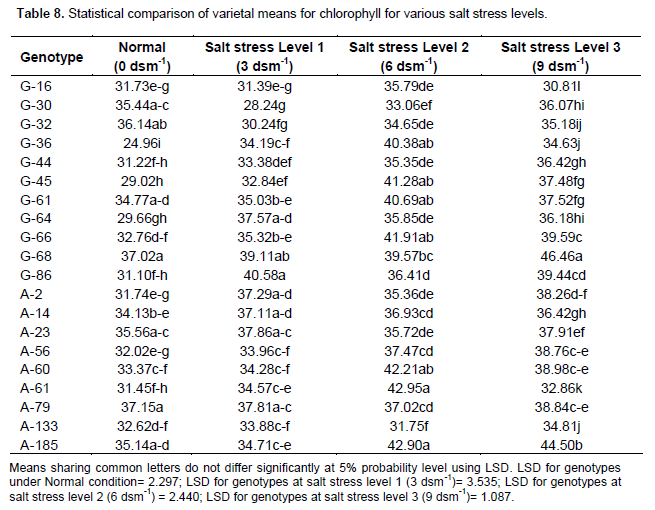
Effect of salt stress (NaCl) on chlorophyll content
Table 8 shows that accession A-79 closely followed by G-68 and G-32 had maximum chlorophyll under normal condition. Accession G-86 closely followed by G-68 and A-23 had maximum chlorophyll under salt stress level 1 (3 dsm-1). Accessions A-61 closely followed by A-185 and A-60 had maximum chlorophyll under salt stress level 2 (6 dsm -1). Genotypes G-68 closely followed by A-185 and G-66 had maximum chlorophyll under salt stress level 3 (9dsm-1).
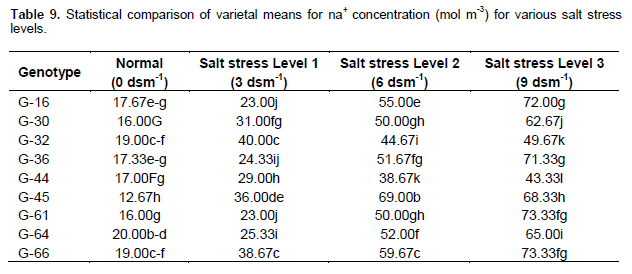
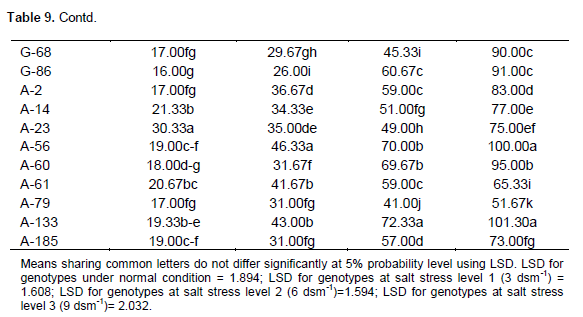
Effect of salt stress (NaCl) on sodium concentration (mol m-3) in extracted leaf sap
Table 9 shows that accession A-23 closely followed by A- 14 and A-61 had maximum Na+ content under normal condition. Accession A-56 closely followed by A-133 and A-61 had maximum Na+ content under salt stress level 1 (3 dsm-1). Genotype A-133 closely followed by A-56 and A-60 had maximum Na+ content under salt stress level 2 (6 dsm -1). The line A-133 closely followed by A-56 and A-60 had maximum Na+ content under salt stress level 3 (9 dsm-1 ). Nawaz et al. (2002) in sunflower also reported that the increase in sodium contents in leaves with increasing salinity was attributed to the increased amount of sodium ion in rooting medium, passive Na+ diffusion through damaged membranes, decreased efficiency of exclusion mechanism.
Effect of salt stress (NaCl) on potassium concentration (mol m-3) in extracted leaf sap of sunflower genotypes
Table 7 shows that accession A-60 closely followed by G-68 and G-32 had maximum K+ content under normal condition. Accession G-32 closely followed by G-44 and G-45 had maximum K+ content under salt stress level 1 (3 dsm-1). The line A-56 closely followed by G-61 and G-44 had maximum K+ content under salt stress level 2 (6 dsm -1). Accession G-68 closely followed by G-66 and A-56 had maximum K+ content under salt stress level 3 (9 dsm-1). Decrease in K+ concentration with increasing salinity was also reported a significant reduction of potassium in sorghum with increasing salinity. There is a debate that K+ influx could be used as an index to salinity tolerance Shainberg and Levy (1992).
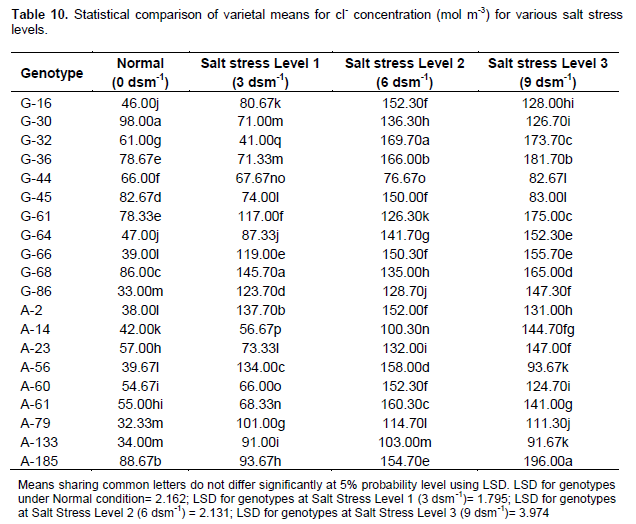
Effect of salt stress (NaCl) on chloride concentration (mol m-3) in extracted leaf sap
Table 10 shows that genotype G-30 closely followed by G-185 and G-68 had maximum Cl- content under normal condition. Accession G-68 closely followed by A-2 and A-56 had maximum Cl- content under salt stress level 1 (3 d Sm-1). The line G-32 closely followed by G-36 and A-61 had maximum Cl- content content under salt stress level 2 (6 d Sm-1). Accession A-185 closely followed by G-36 and G-61 had maximum Cl- content under salt stress level 3 (9 dsm-1).
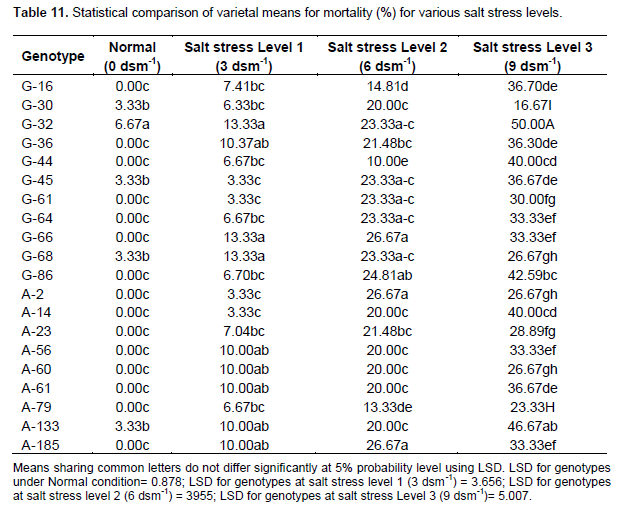
Effect of salt stress (NaCl) on mortality % in sunflower genotypes
Table 11 shows that accession A-185 followed by A-66 and A-79 had minimum mortality % under normal condition. The genotype G -14 followed by G-61 and G-45 had minimum mortality % under salt stress level 1 (3 dsm-1). The line G-44 followed by A-79 and G-16 had minimum mortality % under salt stress level 2 (6 dsm -1). Accession G-30 followed by A-79 and G-68 had minimum mortality % under salt stress level 3 (9 dsm-1). Several researchers have also documented that higher concentration of salt in the rooting medium cause mortality of many plant species (Donahave et al., 1983).
CONCLUSION
The research concluded that the accession G-86 closely followed by G-30 and A-14 had maximum shoot length under salt stress level 3 (9 dsm-1) whereas genotype G- 44 closely followed by G-36 and A-185 had maximum root length under salt stress level 3 (9 dsm-1). Genotypes G-68 closely followed by A-185 and G-66 had maximum chlorophyll under salt stress level 3 (9 dsm-1) whereas line G-30 followed by A-79 and G-68 had minimum mortality % under salt stress level 3 (9 dsm-1).
CONFLICT OF INTEREST
The authors have not declared any conflict of interest.
REFERENCES
| Donahue RL, Miller RW, Shickluna JC (1983). Soils, an introduction to soil and plant growth, Prentice-Hall, Englewood Cliffs, NJ. | ||||
| Ghumman MI (2000). Evaluation of sunflower (Helianthus annuus L.) genotypes for salinity tolerance. M.Sc. Thesis. Dept. PBG, Uni. Agri., Faisalabad. | ||||
| Hussain MK, Rehman OU (1995). Breeding sunflower for salt tolerance. Association of seedling growth and mature plant traits for salt tolerance in cultivated sunflower (Helianthus annuus L.). Helia. 18:69-76 | ||||
| Jumsoon K, Jeuonlai C, Ywonok J (1996). Effect of seed priming on the germinability of tomato (Lycopercicon esculentum Mill.) seeds under water and saline stress. J. Korean Soc. Hortic. Sci. 37:516-521. | ||||
|
Mer RK, Prajith PK, Pandya DH, Pandey AN (2000). Effect of salts on germination of seeds and growth of young plants of Hordeum vulgare, Triticum aestivum. Cick arietinum and Brassica Juncea. J. Agron. Crop Sci. 185:209-217. Crossref |
||||
|
Munns R (1993). Physiological processes limiting plant growth in saline soil: Some dogmas and hypotheses. Plant Cell Environ. 16:1-24. Crossref |
||||
|
Munns RA, Gardrer ML, Rawson HM (1988). Growth and development in NaCl treated plants. II. Do Na+ or Cl- concentrations in dividing or expanding tissue determine growth in barley. Aust. J. Plant. Physiol. 15:529-540. Crossref |
||||
|
Paniego N, Echaide M, Munoz M, Fernandez L, Torales S, Faccio P, Fuxan I, Carrera M, Zandomeni R, Suarez EY, Hopp HE (2002). Microsatellite isolation and characterization in sunflower (Helianthus annuus L.). Genome 4:34-43. Crossref |
||||
| Qureshi RH, Barret-Lannard EG (1998). Saline agriculture for irrigated land in Pakistan. A. Hand Book. ACIAR, Australia. p. 42. | ||||
|
Ramoliya PJ, Panday AN (2003). Effect of salinization of soil an emergence, growth and survival of seedlings of cordial rothii. For. Ecol. Manage. 176:185-194. Crossref |
||||
| Shainberg I, Levy GJ (1992). Physicocemical effects of salts upon infiltration and water movement in soils. P. 37-93. and water movement in soils. pp. 37-93. In: R. J. Wagenet, P. Baveye, and B. A. stewart (eds.). Interacting Processes in soil sci. Advances in Soil Sciences Series, Lewis Publishers, Chelsea, Mich. | ||||
| Szabolcs I (1994). Soils and salination. In Pessarakli M (ed.). Handbook of Plant and Crop Stress. Marcel Dekker, New York, p. 311. | ||||
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


