Full Length Research Paper

ABSTRACT
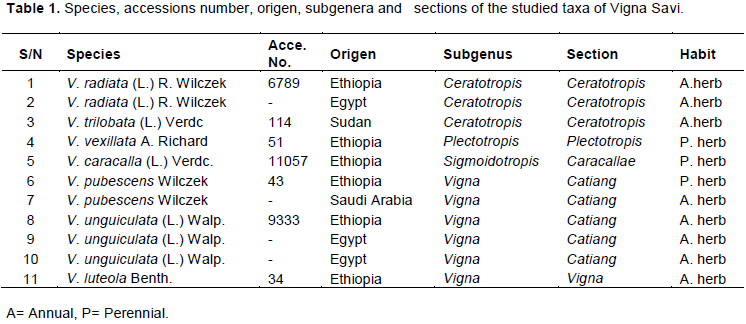
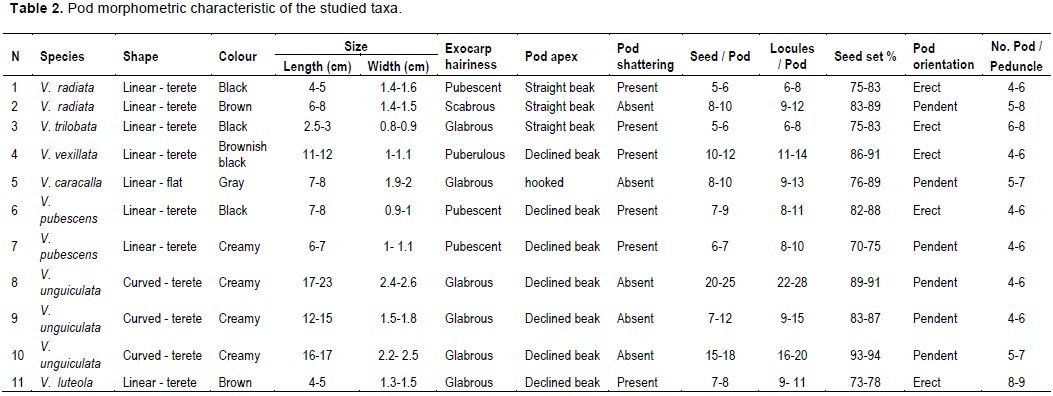
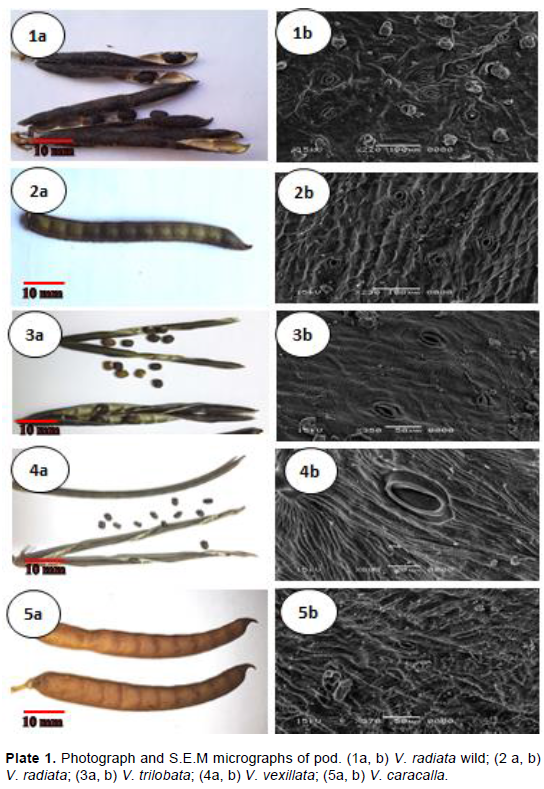
In this study of different taxa of genus Vigna the macro and micromorphological characters were investigated. These taxa collected from different countries (3 cultivated and 8 annual or perennial herbs) representing 7 species of this genus, namely: Vigna radiate, Vigna trilobata, Vigna vexillata, Vigna caracalla, Vigna pubescens, Vigna unguiculata, and Vigna luteola. Legume features such as shape, colour, size, surface texture, number of locules, number of legumes per peduncle seed set percentage, orientation. Moreover, by using scanning electron microscopy, additional details were obtained for stomatal shape, size, type, cuticular and wax ornamentation of the surface of the mature pod, including details of the seed such as hilum shape, size, position, ornamentation based on scanning electron microscope evidence. The usefulness of the macro and micromorphological features as criteria for taxonomic identification was emphasized.
Key words: Vigna, Fabaceae, morphological characters, pod, hilum, S.E.M., taxonomy.
INTRODUCTION
MATERIALS AND METHODS
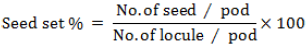
RESULTS
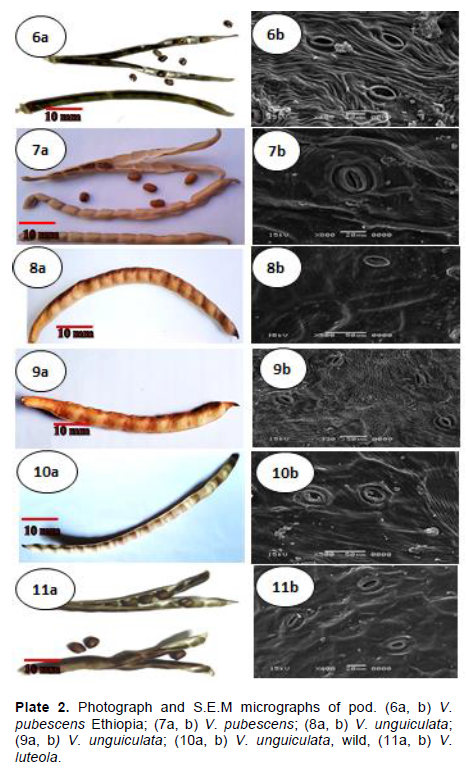
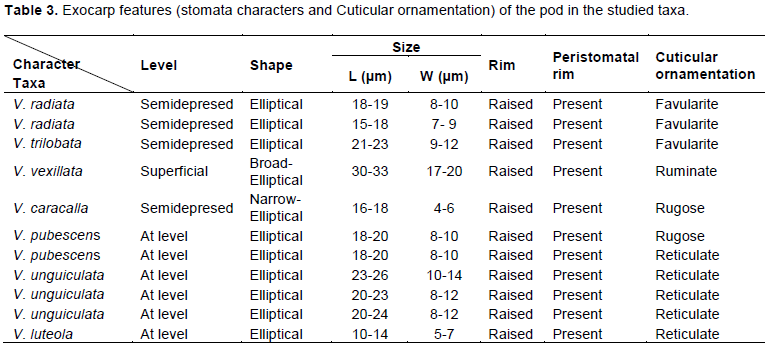
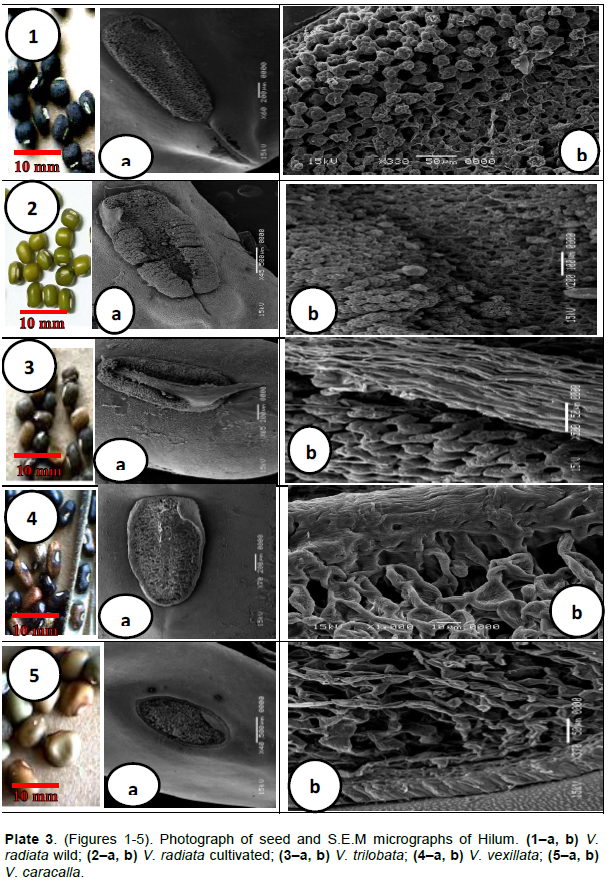
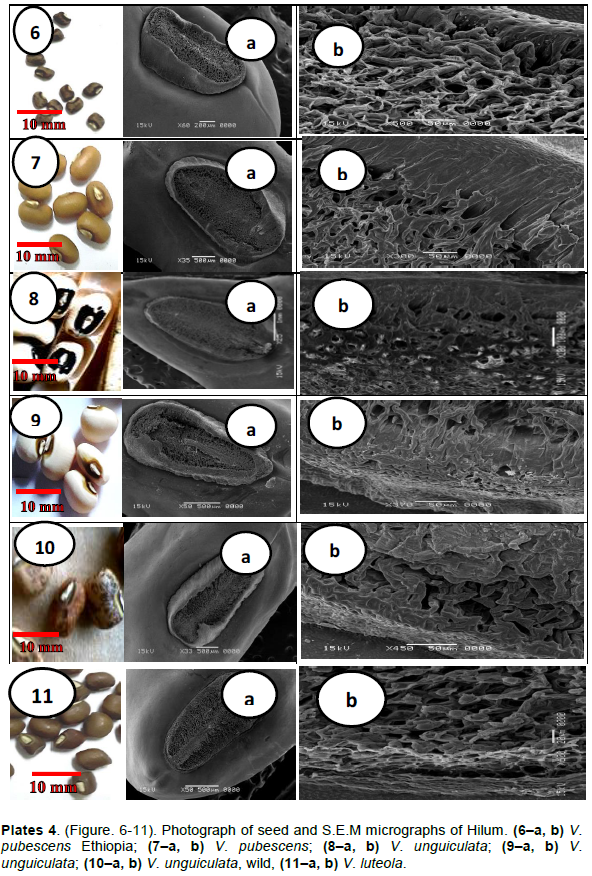
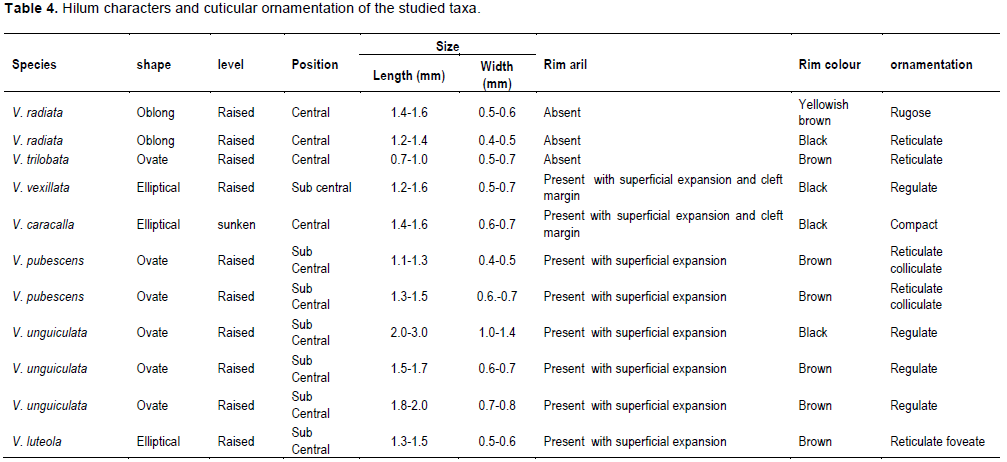
DISCUSSION
CONCLUSION
CONFLICT OF INTERESTS
ACKNOWLEDGEMENT
REFERENCES
|
Akçin ÖE (2008). Seed coat and fruit surface micromorphology of some Cynoglossum L. (Boraginaceae) species. Bangladesh Journal of Botany 37(2)115-119. |
|
|
Barrett RP (1990). Legume species as leaf vegetables. In Advances in new crops. Proceedings of the first national symposium' New crops: research, development, economics', Indianapolis, Indiana, USA, 23-26 October 1988. pp. 391-396. |
|
|
Barthlott W, Ehler N (1977). Raster- Elektronenmikroskopie der Epidermis- Oberflachen von Spermatophyten. Tropische und Subtropische Pflanzenwelt 19:1-110. |
|
|
Barthlott W (1981). Epidermal and Seed Surface Characters of Plants: Systematic Applicability and Some Evolutionary Aspects. Nordic Journal of Botany 1(3):345-355. |
|
|
Barthlott W (1990). Scanning Electron Microscopy of the Epidermal Surface in Plants." In Scanning Electron Microscopy in Taxonomy and Functional Morphology, edited by D. Claugher, 69-94. Oxford: Clarendon. |
|
|
Bisht IS, Bhat KV, Lakhanpaul S, Latha M, Jayan PK, Biswas BK, Singh AK (2005). Diversity and genetic resources of wild Vigna species in India. Genetic Resources and Crop Evolution 52:53-68. |
|
|
Buss CC, Lammers TG, Wise RR (2001). Seed coat morphology and its systematic implications in Cyanea and other genera of Lobelioideae (Campanulaceae). American Journal of Botany 88:1301-1308. |
|
|
Chandel KPS, Malik SK, Nayar ER (1991). SEM Studies of Seed Coat in Some Asiatic Vigna Species and Their Taxonomic and Phylogenetic Significance. Indian Journal of Plant Genetic Resources 4(1):1-11. |
|
|
Chowdhury KA, Buth GM (1970). Seed coat structure and anatomy of Indian Pulses. Botanical Journal of the Linnean Society 63:169-179. |
|
|
Christenhusz MJ, Byng JW (2016). The number of known plants species in the world and its annual increase. Phytotaxon 261:201-217. |
|
|
El-Gazzar A, Abd EL-Ghani, MM, EL-Husseini N, Khattab A (2013). Classification of the Leguminosae-Papilionoideae: A Numerical Re-assessment. Notulae Scientia Biologicae 5(4):499-507. |
|
|
El-Ghamery AA, El-Kholy MA, Gaafar A (2012). Taxonopmic significance of seed protein and iso-enzyme of some species of the genus Vigna Savi (Fabaceae). Journal of Jazan University - Applied Sciences Branch 1(2):37-46. |
|
|
El-Hadidy AMH (2004). Morphological studies on fruits and seeds of the genus Lotus L. In Egypt, Proceedings of First International Conference on Strategy of Egyptian Herbaria. pp. 9-11. |
|
|
Esau K (1953). Anatomy of Seed Plants. John Wiley and Sons, New York. |
|
|
Fabiana SO, Patricia SH, Maria TAG (2013). Morphology of seeds and seedling of four species of Vigna Savi (Leguminosae, Phaseolinae). Acta Botanica Brasillica 27(3):483-489. |
|
|
Fery RL (2002). New opportunities in Vigna. In: J. Janick and A. Whipkey (eds.), Trends in new crops and new uses. ASHS Press, Alexandria, VA. |
|
|
Gaafar A (2007). Cytological Studies on Some Species of the genus Vigna Savi (Fabaceae). M. Sc. Thesis, Botany & Microbiology Department, Faculty of Science, Al-Azhar University. |
|
|
Garba M, Pasquet RS (1998). Isozyme polymorphism within section reticulatae of genus Vigna (Tribe Phaseolae: Fabaceae). Biochemical Systematics and Ecology 26:297-308. |
|
|
Gohary I, Mohammed AH (2007). Seed morphology of Acacia in Egypt and its taxonomic significance. International Journal of Agriculture and Biology 9(3):435-438.. |
|
|
Gontcharova SB, Gontcharova AA, Yakubov VV, Kondo K (2009). Seed surface morphology in some representatives of the genus Rhodiola sect. Rhodiola (Crassulacea) in Russian Far East. Flora 204:17-24. |
|
|
GRIN (2005). National Genetic Resource Program. Germplasm Resources Information Network (GRIN). {Online Database} Germplasm Resources Laboratory, Beltsville, Maryland. Available at: |
|
|
Gunn CR (1981). Seeds of Leguminosae. In: Polhill R.M., Raven P.H. (eds.). Advances in Legume Systematics. Part 2. Royal Botanic Garden, Kew, UK. pp. 913-925. |
|
|
Hymowitz T (1990). Grain legumes. In Advances in new crops. Proceedings of the first national symposium' New crops: research, development, economics', Indianapolis, Indiana, USA, 23-26 October 1988. Timber Press. pp. 154-158. |
|
|
Ismail AMA, El-Ghazaly GA (1990). Phenological studies on Zygophullum qatarense Hadidi from contrasting habitats. Journal of Arid Environments 18(2):195-205. |
|
|
Javadi F, Yamaguchi H (2004). A note on seed coat and plumule morphological variation in the genus Cicer (Fabaceae). Science Report Graduate School of Agriculture and Biological Sciences 56:7-16. |
|
|
Khafagi AAF, El-Ghamery AA, Ghaly NO, Ragab GO (2018). Fruit and Seed Morphology of Some Species of Solanaceae. Taeckholmia 38:123-140. |
|
|
Karcz J, Weiss H, Maluszynska J (2005). Seed coat patterns in Rapid-cycling Brassica forms. Acta Biologica Cracoviensia Series Botanica 47(1):159-165. |
|
|
Khedia R, Manju S, Sharma Kp, Sharma Kc (2017). Scanning electron microscope studies of spermoderm patterns of the three species of Vigna and their cultivars. Research Journal of Biology 5(2):46-53. |
|
|
Koul KK, Ranjan N, Rainas SN (2000). Seed coat micro-sculpturing in Brassica and allied genera (subtribe Brassicinae, Raphaninae, Moricandiinae). Annals of Botany 86(2):285-397. |
|
|
Kumar D, Rangaswamy NS (1984). SEM studies on seed surface of wild and cultivated species of Vigna Savi. Proceedings: Plant Sciences 93(1):35-42. |
|
|
Legume Phylogeny Working Group (LPWG) (2017). Anew subfamily classification of the Leguminosae based on ataxonomically comprehensive phylogeny. Taxon 66(1):44-77. |
|
|
Maréchal R, Mascherpa JM, Stainier F (1978). Etude taxonomique d'un groupe complexe d'espèces des genres Phaseolus et Vigna (Papilionaceae) sur la base de données morphologiques et polliniques traitées par l'analyse informatique [Taxonomic Study of One Complex Group of Species from the Phaseolus -Vigna Genera Based on Morphology and Palynology Data Treated to Computer Analysis]. Boissiera 28:1-273. |
|
|
Maréchal R, Mascherpa JM, Stainier F (1981). Taxonomic study of the Phaseolus-Vigna complex and related genera. In: Polhill RM, Raven PH (eds.). Advances in Legumes Systematics. pp. 329-335. Royal Botanic Gardens, Kew, England. |
|
|
Mbagwu FN, Endeoga HO (2006). Palynological studies on some Nigerian species of Vigna Savi. Journal of Biological Science 6(6):1122-1125. |
|
|
Ng NQ, Marechal R (1985). Cowpea taxonomy, origin and germplasm. Cowpea research, production and utilization, pp. 11-21. |
|
|
Ng NQ (1990). Recent developments in cowpea germplasm collection, conservation, evaluation and research at the Genetic Resources Unit, IITA. Cowpea genetic resources. pp. 13-28. |
|
|
Nath D, Dasgupta T (2015). Study of Some Vigna Species following Scanning Electron Microscopy (SEM). International Journal of Scientific and Research Publications 5(9):1-6. |
|
|
Padulosi S, Ng NQ (1993). A useful and unexploited herb, Vigna marina (Leguminosae-Papilionoideae) and the taxonomic revision of its genetic diversity. Bulletin du Jardin botanique national de Belgique/Bulletin van de Nationale Plantentuin van Belgie 62:119-126. |
|
|
Pasquet RS (1993). Classification infraspécifique des formes spontances de Vigna unguiculata (L.) Walp. á partir de données morphologiques. Bulletin du Jardin botanique national de Belgique/Bulletin van de Nationale Plantentuin van Belgie 62:127-173. |
|
|
Pasquet RS (1998). Morphological study of cultivated cowpea Vigna unguiculata (Le) Walp. Importance of ovule number and definition of cv gr Melanophthalmus. Agronomie 18:61-70. |
|
|
Pasquet RS, Vanderborght T (2000). Isozyme polymorphism Vigna frutescens-V. membranacea complexes (Tribe Phaseoleae, Fabaceae). Plant System Evolution 28:29-43. |
|
|
Peksen A, Peksen E (2013). Agronomic and Morphological Characters of Newly Registered Peksen and Reyhan Vegetable Cowpea Cultivars in Turkey. International Journal of Current Microbiology and Applied Sciences 2(9):133-140. |
|
|
Polhill RM, Raven PH (1981). Advanced in legume systematics part 1. Royal Botanic Gardens, Kew. |
|
|
Popoola JO, Aremu BR, Daramola FY, Ejoh SA, Adegbite AE (2015). Morphometric Analysis of some Species in the Genus Vigna (L.) Walp: Implication for Utilization for Genetic Improvement Journal of Biological Sciences 15(4):156-166. |
|
|
Popoola JO, Adebambo A, Ejoh S A, Agre P, Adegbite A E, Omonhinmin CA (2017). Morphological Diversity and Cytological Studies in Some Accessions of Vigna vexillata (L.) A. Richard. Annual Research and Review in Biology 19(5):1-12. |
|
|
Pule-Meulenberg F, Belane AK, Krasova-Wade T, Dakora F D (2010). Symbiotic functioning and bradyrhizobial biodiversity of cowpea (Vigna unguiculata L. Walp.) in Africa. BMC Microbiology 10(1):89. |
|
|
Sangwan RS, Lodhi G P (1998). Inheritance of flower and pod colour in cowpea (Vigna unguiculata L. Walp.). Euphytica 102:191-193. |
|
|
Shetler SG, Morin NR (1986). Seed morphology in North American Campanulaceae. Annals of the Missouri Botanical Garden. pp. 653-688. |
|
|
Sprent JI, Odee DW, Dakora FD (2010). African legumes: A vital but underutilized resource. Journal of Experimental Botany 61(5):1257-1265. |
|
|
Stearn WT (1996). Botanical Latin. David & Charles, Fourth edit. London. P 546. |
|
|
Takhtajan A (1991). Evolutionary trends in flowering plants: 213-226. New York: Columbia University Press. |
|
|
Tobe H, Wagner WL, Chin HC (1987). A systematic and evolutionary study of Oenothera (Onagraceae): Seed coat anatomy. Botanical Gazette 148(2):235-257. |
|
|
Tomooka N, Maxted N, Thavarasook C, Jayasuriya AHM (2002). Two New Species, Sectional Designation and New Species Combination in Vigna Subgenus Ceratotropis (Piper) Verdc. (Leguminosae, Phaseoleae). Kew Bulletin 57(3):613-624. |
|
|
Tomooka N, Vaughan DA, Moss H, Maxted N (2003). The Asian genus Vigna subgenus Ceratotropis. Genetic Resources. Khuwer Academic publisher, Dordrecht, the Netherlands. pp. 1-270. |
|
|
Umdale SD, Patil PD, Malik SK, Latha MS, Rao R, Yadav SR, Gaikwad NB, Bhat KV (2017). Seed coat sculpture of sub genus Ceratotropis (Piper) Verdc., genus Vigna Savi in India and its taxonomic implication. Botany Letter 1:1-16. |
|
|
Verdcourt B (1970). Studies in Leguminosae-Papilionoideae for the Flora of Tropical East Africa: IV. Kew Bulletin 24:507-569. |
|
|
Yoshizaki M (2003). Millets in prehistoric remain: Paleobotany on barnyard millets and azuki beans in Jaban. In Yamaguchi H, Kawase M (eds). Natural History of Millets. Hokaido University Press, Sapporo. |
|
|
Zhang ZY, Yang DZ, Lu AM, Knapp S (2005). Seed morphology of the tribe Hyoscyameae (Solanaceae). Taxon 54(1):71-83. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


