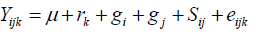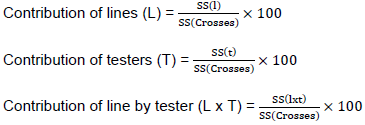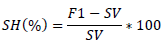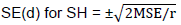ABSTRACT
Generation of information on heterosis and combining abilities of newly developed maize inbred lines is necessary for a successful hybrid and synthetic maize varieties development. Accordingly, this study was conducted to estimate the combining ability of QPM inbred lines for grain yield and yield related traits and to determine the magnitudes of standard heterosis for grain yield and yield related traits in line × tester QPM hybrids. Fifty test crosses together with two standard checks were evaluated using alpha lattice design with three replications at three mid-altitude sub humid trial sites (Bako, Hawassa and Jimma) in Ethiopia during 2016 main cropping season. Combined analysis of variance showed highly significant differences among the three locations for all the studied traits indicating the presence of considerable variation among locations for genotype performance. The interaction between sites and genotypes were highly significant and significant (P<0.05) for grain yield and ear height, indicating that the performances of the genotypes and crosses were not consistent for these traits. The significance of both general combining ability (GCA) and specific combining ability (SCA) mean square for some traits indicates the role of additive and non-additive gene action in the inheritance of the traits. However, for all the traits, the contribution of GCA variance was greater than the contribution of SCA variance, revealing the predominance of additive gene action in the inheritance of all the traits studied. L1 and L3 had significant positive GCA effects and are considered as good combiners for grain yield. In addition, L1 and L9 were good combiners for earliness. In this study, none of the crosses showed positive and significant standard heterosis for grain yield.
Key words: General combining ability, grain yield, specific combining ability, standard heterosis.
Abbreviation:
BNMRC, Bako National Maize Research Center; CM, conventional maize; EIAR, Ethiopian Institute of Agricultural Research; GCA, general combining ability; LSD, least significant difference; masl, meters above sea level; QPM, quality protein maize; SCA, specific combining ability.
Maize is one of the most important field crops cultivated in Ethiopia to ensure food security. Maize contributes the greatest share of production and consumption together with other major cereal crops, such as tef [Eragrostis tef (Zucc.) Trotter], wheat (Triticum aestivum L.) and sorghum [Sorghum bicolor (L.) Moench]. Among the cereal crops, maize ranks second in area coverage and first in total annual production and productivity in Ethiopia (CSA, 2016). Despite its widespread and increased consumption as a source of carbohydrates/energy, maize, like all cereal crops, is known to be poor in its kernel protein quality. The maize protein is limited in two essential amino acids- lysine (C6H14N2O2) and tryptophan (C11H12N2O2) (Bressani, 1991). Protein malnutrition is therefore a serious problem, especially among children, where maize and other cereal crops are the predominant staple foods. Quality protein maize (QPM) is a type of maize variety with improved quality protein content developed after the discovery of maize mutant in the mid 1960’s containing the opaque-2 gene which enhances levels of lysine and tryptophan in the endosperm protein (Mertz et al., 1964).
Consumption of QPM instead of the conventional maize (CM) that has low protein quality can substantially improve the protein status and greatly reduce the malnutrition problem of impoverished people that are dependent on maize as their staple food (Leta et al., 2003). Cognizant of the potential benefits of QPM varieties, the National Maize Research Program of Ethiopia initiated a systematic QPM research in collaboration with CIMMYT in the early 1990s, which led to the identification and release of the first QPM hybrid, BHQP542 in 2002 (Legesse et al., 2012), Melkassa 6Q in 2008 (Gezahegn et al., 2012) and the subsequent release of other several QPM varieties (Adefris et al., 2015). Information on combining ability of parental maize inbred lines, that is, general combining ability (GCA) and specific combining ability (SCA), which determine their performances in hybrid combination, is an important input for designing breeding strategy aimed at exploiting the genetic potential of maize for achieving higher productivity (Chawla and Gupta, 1984).
Combining ability studies can help understand the type of gene action involved in controlling quantitative characters, thereby assisting breeders in selecting suitable parent materials (Hallauer and Miranda, 1988). Heterosis is also important in maize breeding and is dependent on level of dominance and differences in gene frequency. The manifestation of heterosis depends on genetic divergence of the two parental varieties (Hallauer and Miranda, 1988). It is manifested as an increase in vigor, size, growth rate, yield or some other characteristics. But in some cases, the hybrid may be inferior to the weaker parent, which is also considered as heterosis. That means heterosis can be positive or negative. The interpretation of heterosis depends on the nature of trait under study and the way it is measured.
Generally, heterosis is an important trait used by breeders to evaluate the performance of offspring in relation to their parents. It estimates the enhanced performance of hybrids as compared to their parents. Often, the superiority of F1 is estimated over the average of the two parents, or the mid parent. Breeding efforts are underway to convert elite mid-altitude CM inbred lines to QPM through back crossing in recent years in Ethiopia by the breeding program of Bako National Maize Research Center (BNMRC) of the Ethiopian Institute of Agricultural Research (EIAR). This effort has led to the development of many QPM inbred lines, including inbred lines used in this study. Thus, this study was conducted to estimate the combining ability of QPM inbred lines for grain yield and yield related traits and magnitudes of standard heterosis for grain yield and yield related traits in line x tester QPM hybrids.
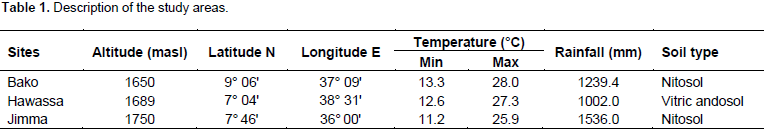
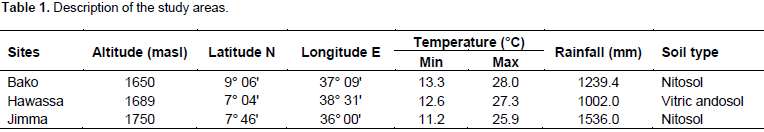
Description of experimental sites
The study was conducted at three locations in the mid-altitude sub-humid agro ecologies of Ethiopia, namely, Bako, Hawassa and Jimma Agricultural Research Centers in the main cropping season of 2016 (Table 1).
Experimental materials
A total of 52 entries composed of 50 test crosses, formed by crossing 25 QPM inbred lines with two single cross testers (referred to as tester A and tester B), and two standard checks (BHQPY545, yellow QPM and BH546, white CM) were studied. The QPM inbred lines were previously developed by BNMRC through backcross breeding technique using elite CM inbred lines as recurrent parents and elite QPM lines as donor parents. The list and the pedigrees of the inbred lines used in the line by tester crosses and that of the testers are given in Table 2. A standard QPM conversion procedure developed by CIMMYT was used to develop the QPM inbred lines, which involved kernel light table screening for endosperm modification, laboratory analysis for tryptophan and lysine contents, as well as field evaluation for agronomic traits. The testers used in this study were identified by CIMMYT Zimbabwe and introduced to Ethiopia by BNMRC breeding program in 2014 main season.

Experimental design and field managements
The experimental design was (0, 1) alpha lattice design (Patterson and Williams, 1976) with 4 plots per an incomplete block and 13 incomplete blocks with three replicates. Each entry was planted in a two row 5.1 m long plot with spacing of 0.75 m between rows and 0.30 m between plants within a row. The experimental materials were hand planted with two seeds per hill, which were later thinned to one plant to get the recommended planting density for the testing sites, 44,444 plants per hectare. Planting was conducted on the onset of the main rainy season after an adequate soil moisture level was reached to ensure good germination and seedling development. Other agronomic practices were carried out as per the recommendation for the test areas.
Data collection
Data on grain yield and other important agronomic traits were collected on a plot and sampled plants/ears bases. Data collected on a plot basis include days to 50% anthesis, days to 50% silking, number of ears per plant, actual moisture content, field weight (kg/plot), plant aspects, ear rot and bad husk cover; while data recorded on sampled plants basis were ear height (cm) and plant height (cm). Yield in t/ha was calculated using CIMMYT fieldbook software (Banziger and Vivek, 2007).
Data analysis
Analysis of variance
Data were subjected to analyses of variance (ANOVA) using the PROC MIXED procedure in SAS® computer program (SAS Institute, 2004). Entries were used as fixed factor while replications and incomplete blocks within replication were considered as random factors. Least significant difference (LSD) was used for mean separation. For traits that displayed significant differences among crosses, line by tester analysis was performed to further partition the variances due to crosses into lines, tester and line by tester effects (Dabholkar, 1999; Singh and Chaudhary, 1985) using SAS program (SAS institute, 2004).
Line by tester analysis
Line by tester analyses was performed for traits that showed significant differences among genotypes as suggested by Dabholkar (1999) and Singh and Chaudhary (1985) to partition the mean square due to crosses into lines (denoting GCA due to lines or males, GCAm), tester (denoting GCA due to testers or females, GCAf) and line x tester interactions (denoting SCA of lines by testers crosses, SCAmf). The following mathematical model was used for the combining ability analysis:
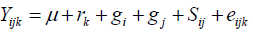
Where, Yijk = the value of a character measured on cross of line i by tester j in kth replication; µ = population mean; rk= effect of kth replication; gi = general combining ability (GCA) effects of ith line; gj = general combining ability (GCA) effect of the jth tester; Sij = specific combining ability (SCA) of ith line and jth testers such that Sij equals Sji; eijk = experimental error for ijkth observation. GCA and SCA of lines were computed for characters that showed significant differences among crosses following line by tester (LxT) analysis as suggested by Singh and Chaudhary (1985). The proportional contributions of lines (GCAL), testers (GCAT), and their interaction (SCALxT) with the sum square of crosses were calculated as the ratio between sum of squares of each component and the cross sum of squares as given by Singh and Chaudary (1985) as follows:
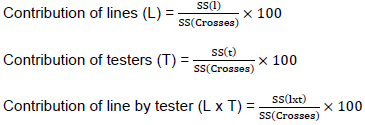
The significance of GCA and SCA effects were tested by dividing the corresponding SCA and GCA values by their respective standard error, to obtain the calculated t values, and comparing the calculated t value with tabular t-value at the error degree of freedom. Standard heterosis (SH) in percent was calculated for those traits that showed statistically significant differences among genotypes as suggested by Falconer and Mackay (1996). These were computed as percentage increase or decrease of the cross performances over best standard check as follows:
Where, F1 = mean value of a cross; SV = mean value of standard check variety. Test of significance for heterosis was done using the t-test. The standard errors of the difference for heterosis were calculated as follows:
Where, SE (d) is standard error of the difference, MSe is error mean square and r is number of replications and calculated t value was compared against the tabulated t-value at degree of freedom for error.
t (standard check) = F1– SV/SE(d)
Analysis of variance
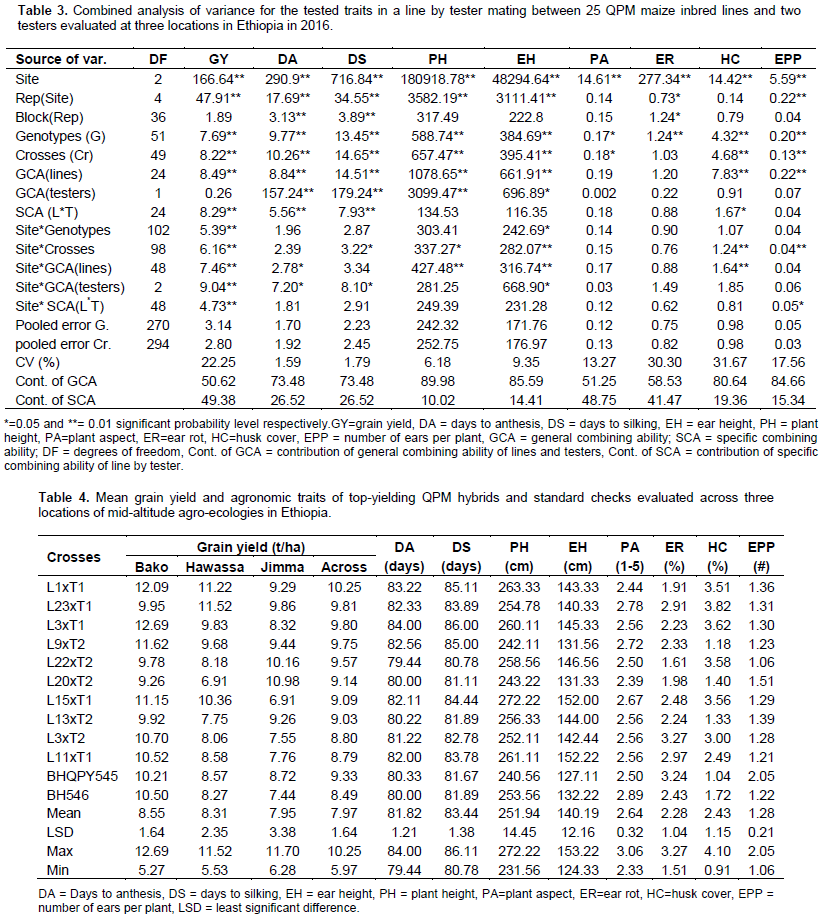
Combined analysis of variance showed highly significant differences among the three locations for all the studied traits (Table 3). The result also showed highly significant (P<0.01) mean squares due to genotypes for all characters studied. The interaction between sites and genotypes (S x G), were highly significant and significant (P< 0.05) for grain yield, days maturity and ear height, indicating that the performances of the genotypes and crosses were not consistent for these traits. However non significant interaction effects of S x G were observed for most of the traits, indicating that the genotypes were performed uniformly across sites for those traits. Generally, the traits which showed significant S x G interaction had a differential genotypic response to variable environmental conditions and this resulted in change in the ranks of genotypes and limited the identification of superior genotypes for all sites. This revealed the site specificity of the genotypes tested (Bayisa et al., 2008).
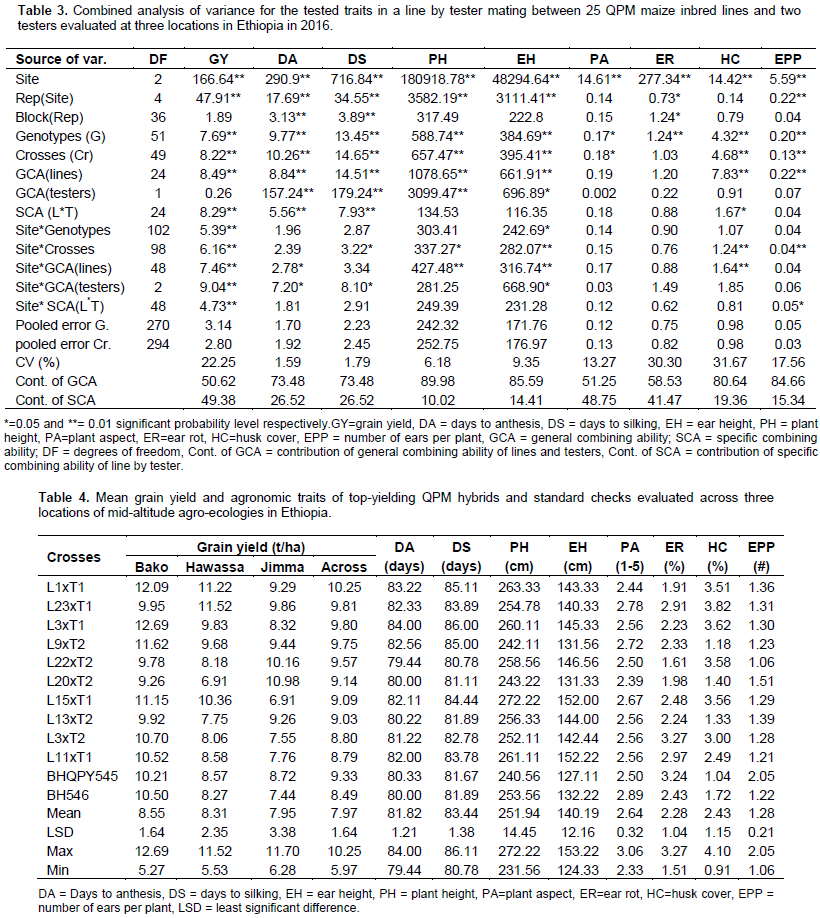
Mean performance of genotypes
The mean performances of the genotypes (the 50 hybrid progenies and two checks) across site are given in Table 4. The mean grain yields (GY) of the genotypes across sites ranged from 5.97 to 10.25 t/ha with overall mean of 7.97 t/ha. The L1 x T1, which was the highest yielding cross (10.25 t/ha) out yielded the high yielding check, BHQPY545 (9.33 t/ha) by 9.86% and the other check BH546 (8.49) by 20.73%. The presence of crosses having mean values better than the standard checks indicate the possibility of obtaining good hybrid (s) for future use in breeding program or for commercial use. Days to anthesis and silking ranged from 79.44 to 84.00 and 80.78 to 86.11 days, with overall means of 81.82, and 83.44 days, respectively. The shortest numbers of days were recorded for crosses L22 x T2 (79.44) days to anthesis and L22 x T2 (80.78) days to silking (Table 3). Most of the crosses showed longest number of days to anthesis and silking.
This shows that those crosses could be grouped as late maturing types. Late maturing crosses are important in the breeding programs for development of high yielding hybrids in areas that receive sufficient rain fall (Girma et al., 2015). Further evaluation and recommendation of this group of materials should be based on agro-ecological suitability. Plant and ear height ranged from 231.56 to 272.22 and 124.33 to 153.22 cm with mean values of 251.94 and 140.19 cm, respectively. The lowest mean values for both plant and ear heights were observed for the cross L19 x T2, while the highest mean values were measured from the crosses L15 x T1 for plant height and L6 x T1 for ear height. Two crosses were significantly taller than the check BH546. Of these crosses, L15 x T1 gave higher grain yield than the best check BH546 (253.56 cm). In line with this finding, Girma et al. (2015) reported higher GY from taller plants and the authors also suggested that this could be attributed to high photosynthetic products accumulation during long period for grain filling.
Number of ears per plant ranged from 1.06 (L22 xT2) to 2.05 (BHQPY545), with an overall mean of 1.28. Among the top yielding crosses, only L20 x T2 had the number of ears per plant greater than 1.5. The mean performances of hybrids for plant aspect (PA) ranged from 2.33 to 3.06, with an overall mean of 2.64. The high yielding cross L1 x T1 was scored 2.44, While the worst (unattractive) PA scored 3.06 was observed from L6 x T2. The mean percentages of ear rot (ER) damage among the hybrids ranged from 1.51 to 3.27%. In general, all the crosses and both standard checks showed small percentage score for ER, which means they could be taken as resistant to this disease under natural infestation. Regarding bad husk cover (HC), the percentage mean value range from 0.91 to 4.1%, with overall mean of 1.15%. Nearly all the crosses evaluated in these trials were free of bad husk cover problem.
Combining ability
In the combined analysis of variance, mean squares due to lines (GCAm) were highly significant for all the studied traits, except plant aspect and ear rot, while mean squares due to testers (GCAf) were highly significant for days to anthesis, days to silking, plant height and ear height. Furthermore, mean squares due to lines by tester interaction (SCAmf) of crosses were significant for grain yield, days to anthesis, days to silking and bad husk cover. The significance of both GCA and SCA mean squares for some traits indicates the role of additive and non-additive gene action in the inheritance of these traits (Table 3). Therefore, recurrent selection which exploits both additive and non-additive gene effects simultaneously could be useful in genetic improvement of the traits studied. However, for all of the traits, the contribution of GCA variance was greater than the contribution of SCA variance, revealing the predominance of additive gene action in the inheritance of all traits.
This showed that parents with good GCA and per se performance could be used to predict the performance of their crosses. Therefore, these parents can be crossed to develop high-yielding QPM hybrids that can potentially be used in further breeding work (inbred line development) and/or directly released for commercial use. Similar results were reported by other authors in their study on combining ability for yield and yield related traits in maize (Bayisa et al., 2008; Chandel and Mankotia, 2014; Seyoum et al., 2016). Highly significant and significant variations were observed due to interaction between sites and GCA of lines and testers for grain yield, days to anthesis and ear height, indicating that the GCA of inbred lines and testers were affected by the environmental conditions under which the hybrids were grown. SCA x site interaction mean square was only highly significant for the grain yield and significant (P< 0.05) for ears per plant.
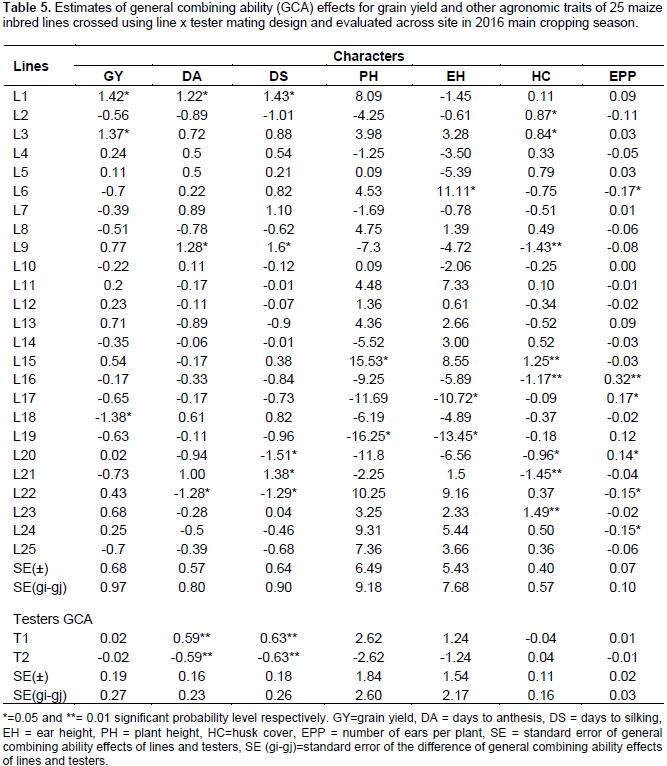
General combining ability effects
Estimates of GCA effects due to lines and testers various traits combined over site are presented in Table 5. Out of the 25 inbred lines studied in line x tester cross, only two exhibited positive and significant GCA effects for grain yield, while one inbred line displayed negative and significant GCA effects for the same trait (Table 5). L1 and L3 had significant positive GCA effects and are considered as desirable good combiner; while only L18 had significant negative GCA effects and considered as undesirable/poor combiner. However, high positive, non-significant and desirable GCA effects were also revealed by L9 and L13. The significant positive GCA effect of lines indicates the potential advantage of the parents for developing high-yielding hybrids. Similar results were reported by various researchers (Kanagarasu et al., 2010; Beyene et al., 2011; Girma et al., 2015; Ram et al., 2015).
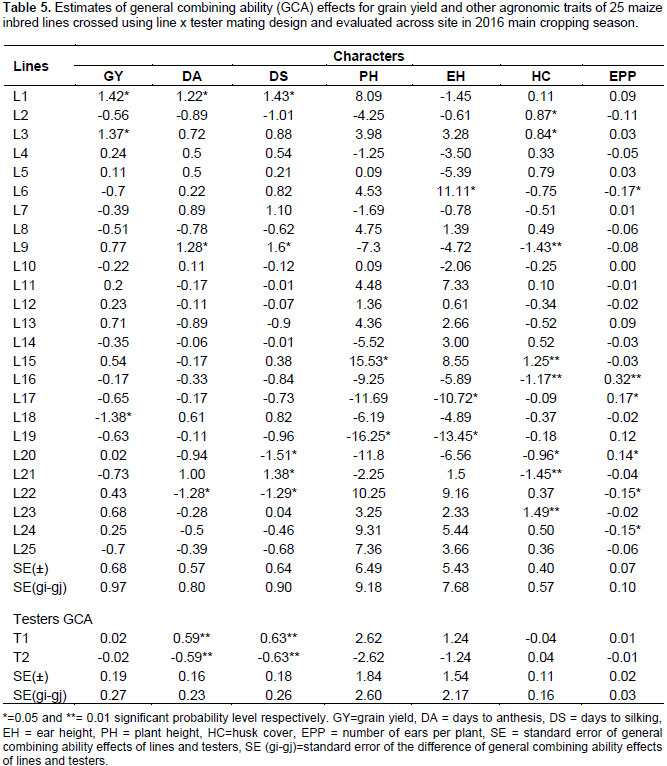
For days to anthesis, L1 and L9, and days to silking, L1, L9 and L21 showed positive and significant GCA effects, while negative and significant GCA effects were observed for L22 for days to anthesis and for L20 and L22 for days to silking. Lines with negative GCA effects for days to anthesis and days to silking are desirable lines, as these lines tend to flower earlier than other lines. Even though there is adequate rainfall in mid altitude agro-ecologies of Ethiopia, effort should be made to develop early maturing varieties to fit fluctuating weather condition. Thus, there is possibility of making effective selection for these traits, which could lead to considerable genetic improvement for earliness. Desirability of negative GCA for days to anthesis and silking was suggested by various authors’ (Iqbal et al., 2007; Shushay et al., 2013; Umar et al., 2014). In addition, T1 had positive and highly significant GCA effects on both days to anthesis and silking, while T2 revealed negative and highly significant GCA effects for both days to anthesis and silking, indicating T2 is a desirable tester for making earliness when crossed with other lines.
The GCA estimates of lines ranged from -2.88 to 1.73 for maturity date (DM). Only L20 showed negative and significant GCA effects for this trait. Inbred lines that showed negative GCA effects for DM could be considered as good general combiners for developing early maturing hybrids to escape late coming disease and pest infestation as well as terminal moisture stress. In line with the current study, other authors reported both positive and negative GCA effects of inbred lines for DM (Habtamu, 2015; Ram et al., 2015). In addition, Girma et al. (2015) reported significant negative and positive GCA effects for DM and suggested that lines with highly significant GCA effects in the negative direction could be used in breeding programs for the introgression of gene for early maturation. For PH and EH, L16, L17, L18, L19 and L20 showed negative GCA effects. For EH, positive significant GCA estimates was observed for L6, while L15, showed positive and significant GCA for PH. Negative GCA effects for EH and PH indicates shorter plant height and lower placement of ear, which is very important for development of genotypes resistant to lodging.
Therefore, inbred lines with significant negative GCA effects are good combiners for hybrid development. Similar results were reported by several authors (Bhatnagar et al., 2004; Aminu and Izge, 2013; Alamerew and Warsi, 2015; Seyoum et al., 2016). About half of the inbred lines revealed positive GCA effects for ear aspect. However, none of the parents showed significant GCA effects for this trait and found to be a good general combiner for quality ear. On the other hand, only L22 revealed significant negative GCA effect for EA. Therefore, L22 is a poorest general combiner for this trait relatively. In case of bad husk cover (HC), L2, L3, L15, and L23 showed positive and significant GCA effects, while negative and significant GCA effects were observed for L9, L16, L20 and L21. A significant negative GCA effect for HC indicates having closed (firm) husk cover and considered as a good combiner in the desired direction (Girma et al., 2015). Regarding ears per plant (EPP), L16, L17, and L20 revealed significant positive GCA effects for EPP, whereas L6, L22 and L24 showed significant negative GCA effects for the same trait. The positive and significant GCA effect for number of ears per plant indicates prolificacy, which is a desirable trait in increasing maize productivity to some extent (Aminu and Izge, 2013; Alamerew and Warsi, 2015).
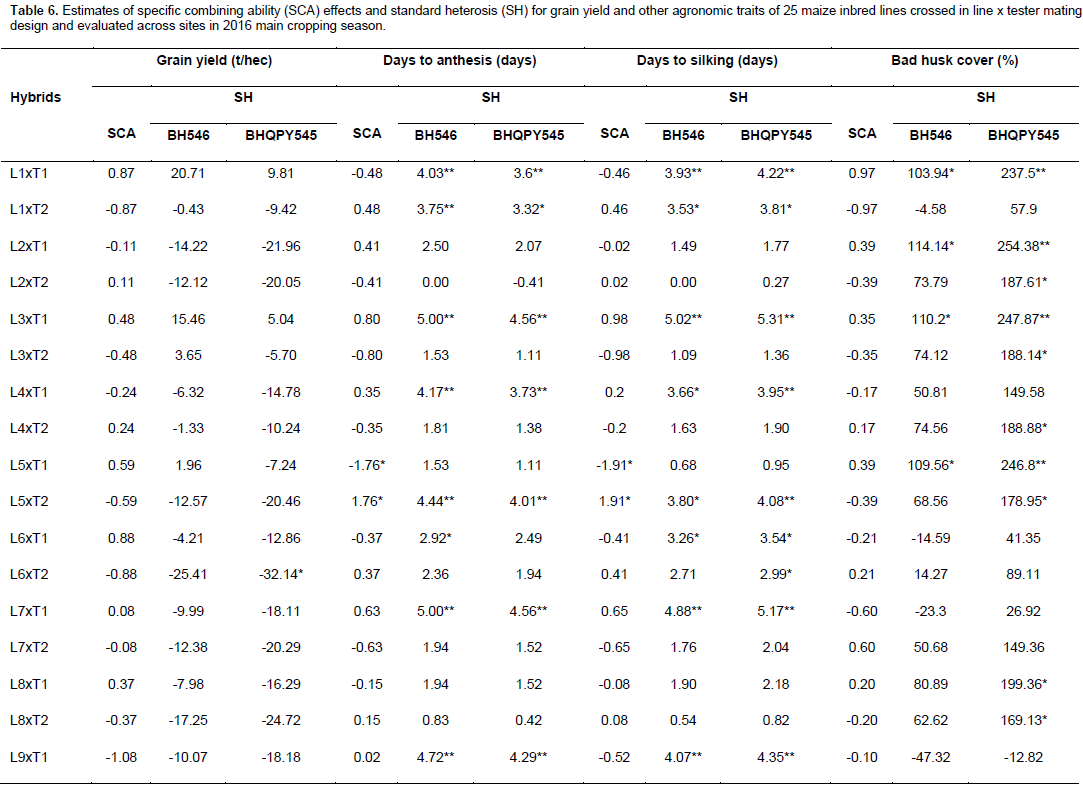
Specific combining ability effects
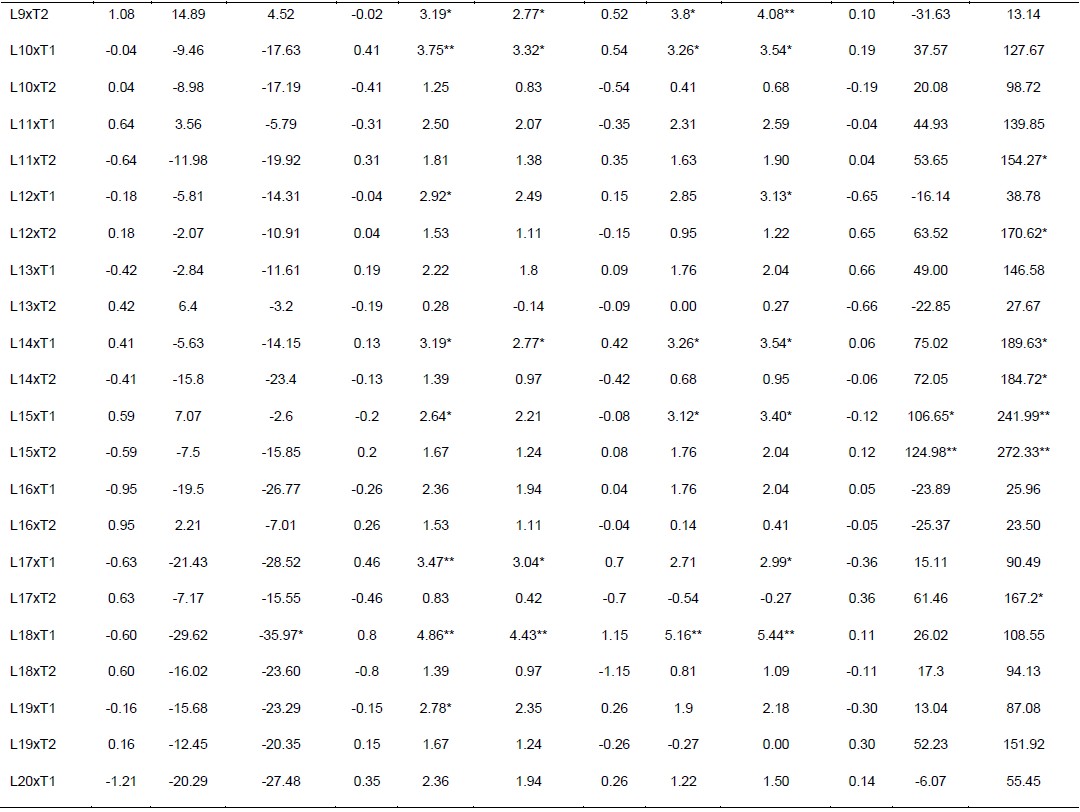
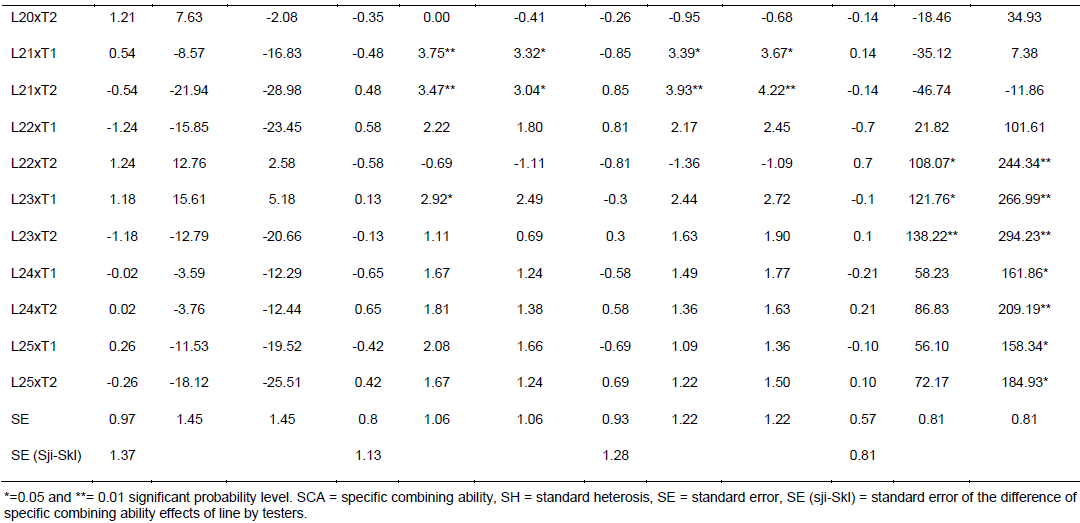
Specific combining ability effects computed for grain yield and other agronomic traits are presented in Table 6. Crosses evaluated in the current study showed limited variation in SCA effects for the traits studied. For GY, crosses L9 x T2, L16 x T2, L20 x T2, L22 x T2 and L23 x T1 revealed highest positive but non-significant SCA effects with SCA values of 1.08, 0.95, 1.21, 1.24, and 1.18, respectively. This indicates that inbred lines involved in these crosses are genetically divergent, and hence could be regarded to be from different heterotic groups. L9 x T1, L16 x T1, L20 x T1, L22 x T1 and L23 x T2 showed lowest negative but non-significant SCA effects for this trait, indicating that these crosses were poor specific combiners for grain yield. All crosses that showed the highest positive SCA effects, except L23 x T1, resulted from poor inbred lines by poor tester for grain yield. This showed that, the crosses performed better than what would be expected from the GCA effects of their respective parents. Therefore, these crosses could be selected for their specific combining ability for higher grain yield. Non significant SCA effect for grain yield was previously reported by Seyoum et al. (2016).
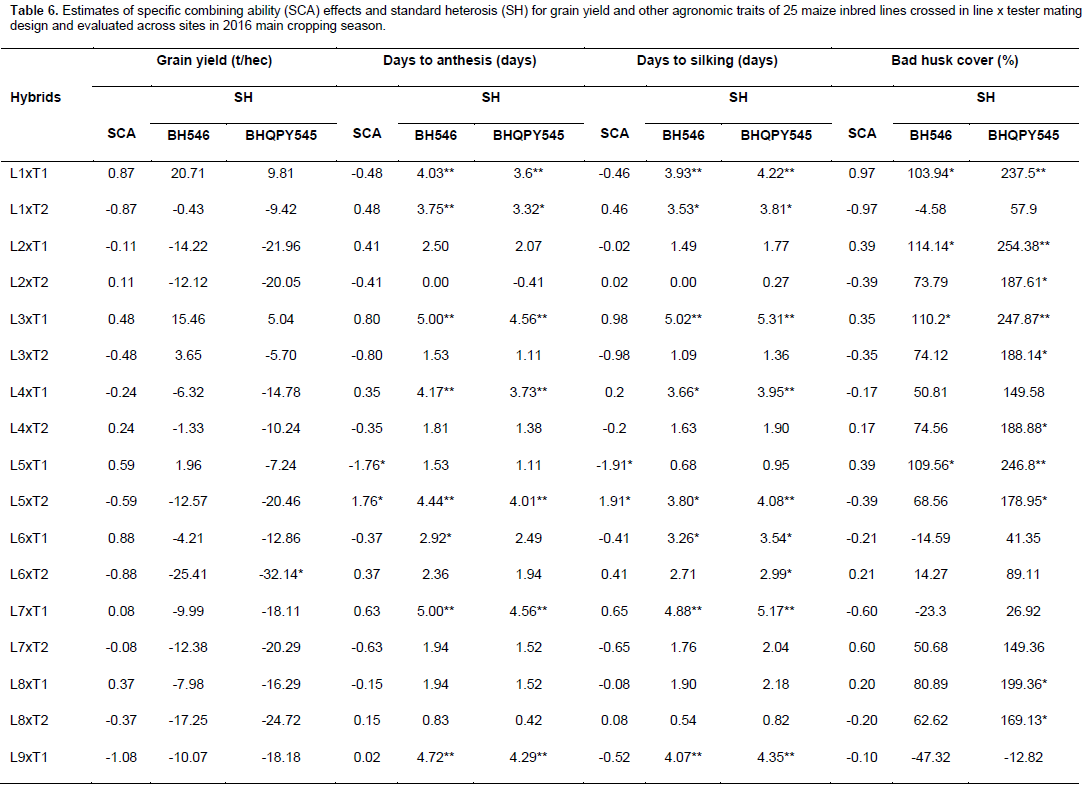
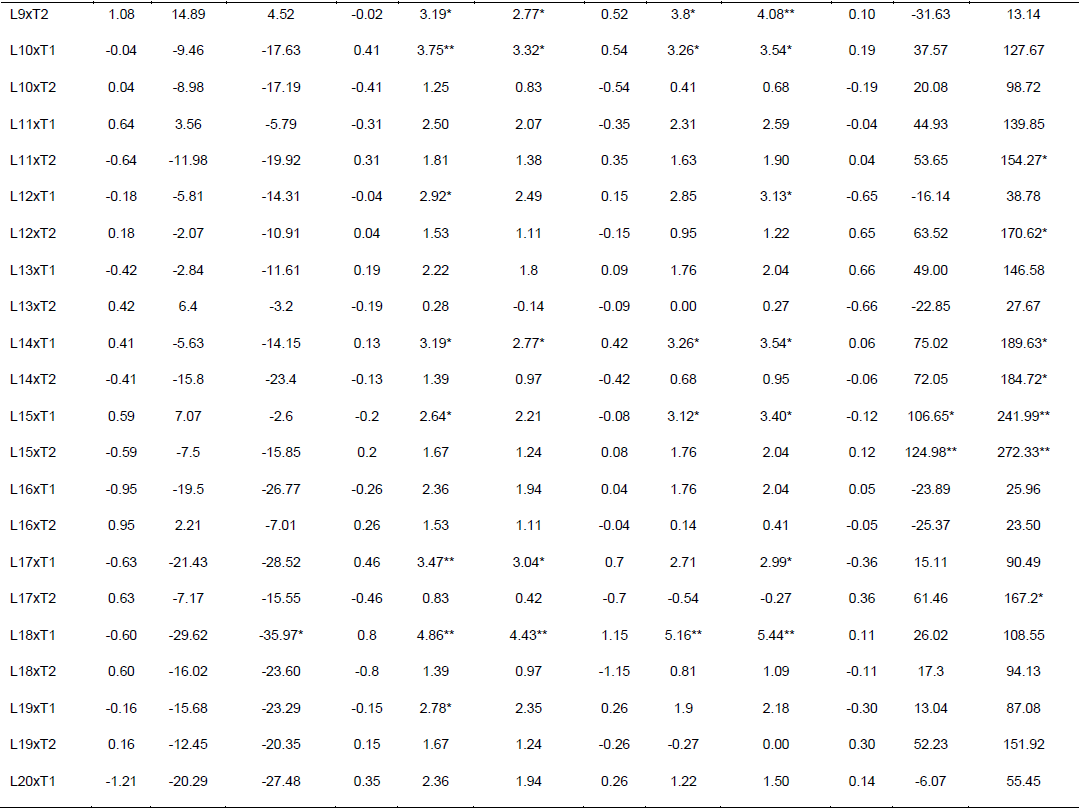
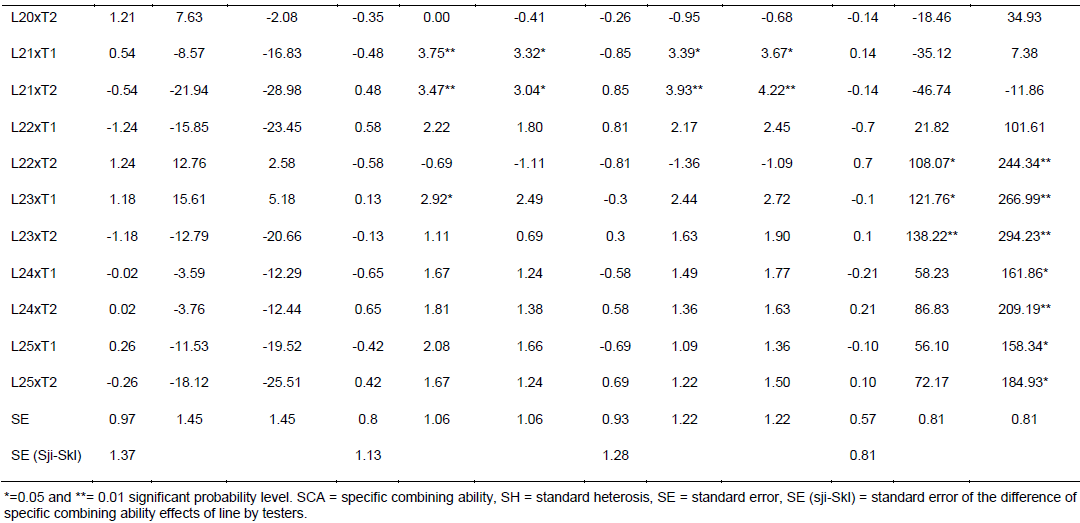
In contrast to this finding, Bullo and Dagne (2016) reported highly significant positive and negative SCA effects for GY and they suggested that, when high yielding specific combinations are desired, especially in hybrid maize development, SCA effects could help in the selection of parental material for maximum exploitation of heterosis. For days to anthesis and silking, only a few crosses showed significant SCA effects in both directions. L5 x T2 showed significant and positive SCA estimate, while L5 x T1 showed significant SCA estimate for DA for both traits. The hybrids with low SCA for days to anthesis and days to silking are desirable as they have earlier anthesis and silking days than what is expected based on GCA of their parents. This finding is in agreement with Kanagarasu et al. (2010), Dagne et al. (2011), Aminu and Izge (2013) and Aminu et al. (2014). None of the inbred lines had significant SCA effect in both directions for bad husk cover. The result demonstrated that most of the crosses evaluated in the current study did not significantly deviate from what would have been predicted based on their parental performance for almost all the traits. This is expected since the proportions of GCA effects were higher than that of SCA.
Standard heterosis
The estimates of standard heterosis over the standard checks were computed for grain yield and yield related traits and presented in Table 6. None of the crosses showed positive and significant heterosis over both standard checks for grain yield. Standard heterosis (SH) for this trait ranged from -29.62 (L18 x T1) to 20.71% (L1 x T1) over BH546, and -35.97 (L18 xT1) to 9.81% (L1 x T1) over BHQPY545. L3 x T1 (15.46 and 5.04%) and L23 x T1 (15.61 and 5.18%) also exhibited positive standard heterosis over both checks. Positive heterosis for this trait indicates increased yield advantage over the existing standard check. Maize hybrids that perform better than the checks could be used for release as hybrid variety after verification. Standard heterosis for DA ranged from -0.69 to 5.00% over BH546 and -1.11 to 4.56% over BHQPY545. For DS, the value of SH ranged from -1.36 to 5.16% over BH546 and -1.09 to 5.44% over BHQPY545. None of the crosses displayed negative and significant SH for DA and DS over both checks.
On the other hand, 14 crosses showed positive and significant heterosis for DA and DS over both checks indicating, those crosses were late maturing as compared to the checks. Heterosis in the negative direction for these traits indicates earliness of the crosses over the standard checks. In contrast to the current finding, Amiruzzaman et al. (2013) and Bello and Olawuyi (2015) reported negative and significant SH for DA and DS in most of the crosses. Standard heterosis for bad husk cover (HC) ranged from -47.32 to 138.22% and -12.82 to 294.23% for BH546 and BHQPY545, respectively. The negative heterosis in this trait indicates desirable crosses with closed ear up to the tip of the cob, while crosses with positive heterosis for this trait showed bad husk cover and may be susceptible to ear rot disease and are predisposed to other damages.
Combined analysis of variance indicated the presence of considerable variation among locations for genotypes performances. Furthermore, mean squares due to GCA of lines and SCA of crosses were significant for grain yield, days to anthesis, days to silking and bad husk cover. The significance of both GCA and SCA mean squares for these traits, indicate the role of additive and non-additive gene action in the inheritance of these traits. L1, L3, L9 and L13 were identified as good combiner for grain yield and L22 and T2 were identified as good combiner for reducing days to anthesis and silking. The inbred lines having significant negative GCA for days to anthesis and silking identified in this study could be used as parents for breeding quality protein maize for earliness in the mid-altitude sub- humid agro-ecology of Ethiopia.
Likewise, L9 x T2, L16 x T2, L20 x T2, L22 x T2 and L23 x T1 revealed highest positive SCA effects. This indicates that inbred lines involved in these hybrids are genetically divergent, and hence could be regarded to be from different heterotic groups. though, none of the crosses showed positive and significantstandard heterosis for grain yield, some crosses showed positive heterosis over both standard checks. Maize hybrids that perform better than the checks could be used for release as hybrid variety after re-evaluation in multi-location trials. Generally, the results obtained in this study could be helpful to design appropriate breeding strategy for developing QPM hybrids and synthetics adapted to the mid altitude sub-humid agro-ecologies of Ethiopia.
The authors have not declared any conflict of interests.
We would like to express our sincere appreciation to the maize research staff at Bako, Hawassa and Jimma agricultural research centers for hosting the trials and collecting data. We also extend our thanks to the Ethiopian Institute of Agricultural Research (EIAR) for their financial support.
REFERENCES
|
Adefris T, Dagne W, Abraham T, Birhanu T, Kassahun B, Dennis F, Prasanna, BM (2015). Quality Protein Maize (QPM): A Guide to the Technology and Its Promotion in Ethiopia. CIMMYT: Addis Ababa, Ethiopia.
|
|
|
|
Alamerew S, Warsi M (2015). Hetrosis and combining ability of sub tropical maize inbred lines. Afr. J. Crop Sci. 23:123-133.
|
|
|
|
Aminu D, Izge A (2013). Gene action and heterosis for yield and yield traits in maize(Zea mays L.), under drought conditions in Northern guinea and Sudan savannas of Borno State, Nigeria. Peak J. Agric. Sci. 1:17-23.
|
|
|
|
Aminu D, Mohammed S, Kabir B (2014). Estimates of combining ability and heterosis for yield and yield traits in maize population (Zea mays L.), under drought conditions in the northern Guinea and Sudan savanna zones of Borno State, Nigeria. Int. J. Agric. Innov. Res. 2:824-830.
|
|
|
|
Amiruzzaman M, Islam MA, Hasan L, Kadir M, Rohman MM (2013). Heterosis and combining ability in a diallel among elite inbred lines of maize (Zea mays L.). Emir. J. Food Agric. 25:132.
Crossref
|
|
|
|
Banzige M, Vivek BS (2007). Fieldbook: Software for Managing a Maize Breeding Program. CIMMYT.
|
|
|
|
Bayisa A, Mohammed H, Zelleke H (2008). Combining ability of transitional highland maize inbred lines. East. Afr. J. Sci. 2:19-24.
|
|
|
|
Bello OB, Olawuyi OJ (2015). Gene action, heterosis, correlation and regression estimates in developing hybrid cultivars in maize. Trop. Agric. 92:102-117.
|
|
|
|
Beyene Y, Mugo S, Gakunga J, Karaya H, Mutinda C, Tefera T, Njoka S, Chepkesis D, Shuma JM, Tende R (2011). Combining ability of maize (Zea mays L.) inbred lines resistant to stem borers. Afr. J. Biotechnol. 10:4759-4766.
|
|
|
|
Bhatnagar S, Betran F, Rooney L (2004). Combining abilities of quality protein maize inbreds. Crop Sci. 44:1997-2005.
Crossref
|
|
|
|
Bressani R (1991). Protein quality of high lysine maize for humans. Cereal Foods World 36:806-811.
|
|
|
|
Bullo NT, Dagne WG (2016). Combining ability of inbred lines in quality protein maize (QPM) for varietal improvement. Int. J. Plant Breed. Crop Sci. 3(1):79-89.
|
|
|
|
Chandel U, Mankotia B (2014). Combining ability in local and cimmyt inbred lines of maize (Zea mays L.) For grain yield and yield components using line× tester analysis. SABRAO J. Breed. Genet. 46:256-264.
|
|
|
|
Chawla HS, Gupta VP (1984). Index India-Agriculture. Indian Calcutta Agriculture 28(4):261- 265.
|
|
|
|
Central Statistical Agency (CSA) (2016). Agricultural Sample Survey 2015/2016: Report on area and production of major crops (private peasant holdings, 'Meher' season). Statistical Bulletin. 1. CSA, Addis Ababa, Ethiopia.
|
|
|
|
Dabholkar AR (1999). Elements of biometrical genetics. Revised and enlarged edition, Concept Publishing Company. New Delhi, India.
|
|
|
|
Dagne W, Vivek B, Tadesse B, Abdissa K, Worku M, Wolde L (2011). Combining ability and heterotic relationships between CIMMYT and Ethiopian maize inbred lines. Ethiop. J. Agric. Sci. 21:82-93.
|
|
|
|
Falconer DS, Mackay TF (1996). Introduction to quantitative genetics, 4th edition. Longman, London, UK.
|
|
|
|
Gezahegn B, Dagne W, Lealem T, Deseta G (2012). Maize Improvement for Low-Moisture Stress Areas of Ethiopia: Achievements and Progress in the Last Decade. In. Worku M, Twumasi-Afriyie S, Wolde L, Tadesse B, Demisie G, Bogale G, Wegary D, Prasanna BM (eds.), 18-20 April 21012. Proceedings of the 3rd National Maize Workshop of Ethiopia. Addis Ababa, Ethiopia. pp. 35-46.
|
|
|
|
Girma C, Alamerew S, Tadesse B, Menamo T (2015). Test Cross Performance and Combining Ability of Maize (Zea mays L.) Inbred Lines at Bako, Western Ethiopia. Glob. J. Sci. Front. Res. 15(4):1-24.
|
|
|
|
Habtamu Z (2015). Heterosis and Combining Ability for Grain Yield and Yield Component Traits of Maize in Eastern Ethiopia. Sci. Technol. Arts Res. J. 4(3):32-37.
|
|
|
|
Hallauer AR, Miranda JB (1988). Quantitative genetics in maize breeding, 2nd edition. Iowa State University Press, Iowa, Ames.
|
|
|
|
Iqbal AM, Nehvi F, Wani S, Qadir R, Dar ZA (2007). Combining ability analysis for yield and yield related traits in maize (Zea mays L.). Int. J. Plant Breed. Genet. 1:101-105.
Crossref
|
|
|
|
Kanagarasu S, Nallathambi G, Ganesan K (2010). Combining ability analysis for yield and its component traits in maize (Zea mays L.). Electr. J. Plant Breed. 1:915-920.
|
|
|
|
Legesse W, Mosisa W, Berhanu T, Girum A, Wande A, Solomon A, Tolera K, Dagne W., Girma D, Temesgen C, Leta T, Habtamu Z, Habte J, Alemu T, Fitsum S, Andualem W, Belayneh A (2012). Genetic improvement of maize for mid-altitude and lowland sub-humid agro-ecologies of Ethiopia. In. Worku M, Twumasi-Afriyie S, Wolde L, Tadesse B, Demisie G, Bogale G, Wegary D, Prasanna BM (eds.), 18-20 April 2012. Proceedings of the Third National Maize Workshop of Ethiopia. Addis Ababa, Ethiopia. pp. 24-34.
|
|
|
|
Leta T, Gelana S, Jemal A, Dagne W, Twumasi AS (2003). Historical Perspectives, Current status, and Strategies for Quality Protein Maize Breeding in Ethiopia. In. CIMMYT 2003. Book of Abstracts: Arnel R.Halauer Internal Symposium on Plant Breeding, 22 August 2003, CIMMY, Mexico. pp. 124-125.
|
|
|
|
Mertz ET, Bates LS, Nelson OE (1964). Mutant gene that changes protein composition and increases lysine content of maize endosperm. Science 145(3629):279-80.
Crossref
|
|
|
|
Patterson HD, Williams ER (1976). A new class resolvable incomplete block designs. Biometrika 63:83-92.
Crossref
|
|
|
|
Ram L, Singh R., Singh S, Srivastava R (2015). Heterosis and combining ability studies for quality protein maize. Ekin J. Crop Breed. Genet. 1-2:8-25.
|
|
|
|
Statistical Analysis System (SAS) (2004). SAS Institute Inc., Cary, NC, USA.
|
|
|
|
Seyoum A, Wegary D, Alamerew S (2016). Combining Ability of Elite Highland Maize (Zea mays L.) Inbred Lines at Jimma Dedo, South West Ethiopia. Adv. Crop Sci. Technol. 4:212.
|
|
|
|
Shushay A, Habtamu Z, Dagne W (2013). Line x tester analysis of maize inbred lines for grain yield and yield related traits. Asian J. Plant Sci. Res. 3:12-19.
|
|
|
|
Singh RK, Chaudhary BD (1985). Biometrical methods in quantitative genetic analysis. Kalyani Publishers New Dehli, India.
|
|
|
|
Umar U, Ado S, Aba D. Bugaje S (2014). Estimates of combining ability and gene action in maize (Zea mays L.) under water stress and non-stress conditions. J. Biol. Agric. Healthc. 4(25):247-253.
|