ABSTRACT
Maize lethal necrosis disease (MLND) in Kenya has been reported to be caused by a coinfection between a non-potyvirus Maize chlorotic mottle virus (MCMV) and a potyvirus Sugarcane mosaic virus (SCMV). The control of the disease in Kenya has been a challenge owing to the synergistic interactions that exist between the two viruses. This study, sought to determine the stage of synergism between the two viruses and its role in influencing the severity of the disease. Three maize hybrids were grown in a greenhouse and were mechanically inoculated with MCMV, SCMV and MCMV+SCMV at the vegetative stage, V4-5. The synergism was studied for a period of 90-days and double-antibody sandwich (DAS)-ELISA was used to estimate the viral titer of MCMV and SCMV under individual and co-infection states of maize plants. The results showed that the viral titers of the two viruses in both single and double infection followed a normal curve. Synergistic effect was observed between the 21- and 28-days post-inoculation (dpi). A significant increase in the titers of MCMV was observed at this time in days, while that of SCMV was more or less constant. Also, the study revealed that viral titers of SCMV in both individual and co- infected maize plants remained constant; while the viral titers of MCMV in co- infected maize increased significantly as compared to the individual infections. Furthermore, there was a positive correlation between increased symptom severity and synergism. Based on these results, SCMV plays a major role in the severity and spread of MLN disease in the South-Rift region.
Key words: Maize lethal necrosis, maize chlorotic mottle virus, Sugarcane mosaic virus, maize virus diseases.
Studies on synergistic interactions between two pathogenic viruses have been reported to be common among plants (Mahuku et al., 2015). Potyvirus-associated synergisms are the most common type of synergism, in which one of the viruses is a member of the potyvirus group and the other is not (Mbega et al., 2016). Maize lethal necrosis (MLN) disease in maize is a classic example of potyvirus-associated synergism because it involves a synergistic relationship between Maize chlorotic mottle virus (MCMV) and any potyvirus that has been confirmed to cause the disease, such as Sugarcane mosaic virus (SCMV), Maize dwarf mosaic virus (MDMV), or Wheat streak mosaic virus (WSMV) (Isabirye and Rwomushana, 2016). SCMV has been reported as the most common potyvirus in synergy with MCMV causing MLN in Kenya and other countries of East Africa (Adams et al., 2014). Leitich et al. (2020) also reported that SCMV was the primary potyvirus causing MLN disease in Kenya's South-Rift region, in synergy with MCMV. Furthermore, Mbega et al. (2016) reported MCMV as a primary disease-causing virus of MLN with the potential to establish itself alone in warm, arid, semi-arid, and sub-humid tropics in his review (Isabirye and Rwomushana, 2016). Synergism has been reported to increase symptom severity such as; chlorotic mottling, leaf necrosis from the margins to the midrib, stunted growth, premature death or ageing, male sterility and failure to tassel, rotten or small cobs with little or no grain (Xia et al., 2016; Wangai et al., 2012). Such symptoms make MLN a devastating maize disease. The synergistic interactions are more pronounced (Mbega et al., 2016)and result in serious damage that usually kills the infected plant (Makone et al., 2014). The disease has been reported to have caused an estimated loss of $187 million equivalent to $364/ton in Kenya since it was first reported in 2011 (De Groote et al., 2016). This is a direct loss to farmers especially those who rely on the crop for food production and income (Bulegeya, 2016).
In Kenya, the disease has since spread to other maize-growing areas since its first report in 2011. The areas include; Central, Nyanza, Western, South and North-Rift regions of Kenya (Karanja et al., 2018). Although there were efforts to contain the spread of the disease in the country, the farmers in MLN hotspot areas have continued experiencing significant yield losses as high as 100% due to the severity of the disease (Kagoda et al., 2016). The huge losses of the crop yield have been linked to virus synergism (Mbega et al., 2016). Therefore, these huge losses could be attributed to limited information on the effect of synergism between the two viruses in the severity of MLN and management. It has been reported that understanding the mechanism behind synergism and the time it occurs can contribute to more effective management through resistance breeding targeted at the components, specifically potyvirus leading to reduction in yield losses (Mahuku et al., 2015). Therefore, this study sought to establish the stage at which synergism is at the peak when the two viruses (MCMV + SCMV) co-infect the maize plants at the same time. The results will be useful to maize breeders in breeding for tolerant/resistant against the MLN disease.
Site of the experiment
A screen house experiment was carried out between June and October of 2017 at the CIMMYT-MLN screening site in Naivasha (latitude 0°43′S, longitude 36°26′E, 1896 m.a.s.l.) to assess the stage of synergistic interaction between MCMV and SCMV.
Experimental design
The experiment was carried out in a netted screenhouse where insect vectors were controlled. The trial was a two-level factorial experiment (3×4) in a complete randomized design with three replicates. The factors studied were; hybrids at three levels namely; (Tolerant to MCMV-CKMLN150078, CKH12603 and Susceptible to MCMV-DUMA43) and inoculation combinations at four levels namely; (MCMV, SCMV, MCMV+SCMV and uninoculated control).
Planting, inoculum preparation and inoculations
To check virus purity, inoculation, and disease evaluation for MCMV and SCMV, the serological assay ELISA was used. Prior to planting, the seeds were tested in the laboratory using DAS-ELISA to ensure that they were virus-free. Three clean seeds from each entry (tolerant and susceptible hybrids) were planted in a pot of sterile soil mixture of red soil, pit moss, and compost manure at a ratio of 3:1:1 respectively. The MLN inoculum was achieved by harvesting the leaves of plants that had been artificially inoculated with two viruses at 3-4 leaf stage. The infected MCMV and SCMV leaves were harvested from the greenhouse in a 1:4 ratio (MCMV: SCMV). The infected MCMV and SCMV leaves were blended separately in a cold 0.1 M phosphate extraction buffer using a dilution ratio of 1:10 (leaf material: buffer). The extract was then sieved through folded cheesecloth to remove any debris. For the double inoculations, the inoculum from MCMV and SCMV was mixed in a bucket, and 1 g of celite, abrasive agent was added per litre and stirred thoroughly to ensure even distribution of the celite. All the plants were mechanically inoculated at the 3-4 leaf stage by rubbing the two youngest leaves together (Karanja et al., 2018). The inoculum was then kept cool during the inoculations using ice cubes.
Synergistic interactions between MCMV and SCMV
To determine the stage of synergism between MCMV and SCMV, the plants were mechanically inoculated with MCMV, SCMV, and (MCMV+SCMV) at the 3-4 leaf stage using equal volumes of 5 ml of inoculum per plant. Seven plants from each treatment were randomly selected and tagged for sampling one week after inoculation. For a span of 90 days, severity ratings on a scale of 1 to 5 were done at weekly intervals. The leaf sample was collected using sampling bags, labelled, and placed in a cool box. The collected samples were taken to the laboratory for virus detection using a double-antibody sandwich (DAS)-ELISA.
Symptom’s identification/rating
Plants were assessed for virus symptoms starting at 7 days post-inoculation (dpi) and then every 1 week for the next 90 days, using a 1 to 5 scale, with 1 indicating no symptoms, 3 indicating moderate symptoms, and 5 indicating severe chlorosis. The diagnoses included whether the signs were local lesions on inoculated leaves or systemic infections with mosaics, mottles, flecks or mottles that were limited or general.
Virus detection using DAS-ELISA
Relative amounts of MCMV and SCMV in leaf samples were determined using DAS-ELISA as described by Dijksta and De Jager (1998). Samples were taken weekly for a period of ninety days to understand the disease development and interaction between the two viruses. The leaf material (5 g) of the infected plant was extracted using phosphate-buffered saline (10 mM potassium phosphate, 150 mM sodium chloride), pH 7.4, containing Tween 20 at 5 ml/L and polyvinyl pyrrolidone at 20 g/L, using a mixer mill (Retsch, Germany). Each sample was tested in duplicate wells in microtiter plates and commercial MCMV and SCMV antiserum (DSMZ, Braunschweig) were included in paired wells as controls. The substrate used was p-nitrophenyl phosphate at 0.6 mg/ml in diethanolamine at 100 ml/L, pH 9.8. The plates were measured at an absorbance of 405 nm in a microplate reader (Bio-Rad Laboratories).
Symptom expression on maize plants
Symptom expression between single and double infections was compared. The single infections of MCMV and SCMV alone, induced relatively mild symptoms comprising of chlorosis and mottling for MCMV (Figure 1A) and mild mosaic and mottling for SCMV (Figure 1B). As for the double infection with MCMV+SCMV, it resulted in the enhancement of symptoms comprising of severe chlorotic mottling and yellow streaks parallel to leaf veins (Figure 1C).
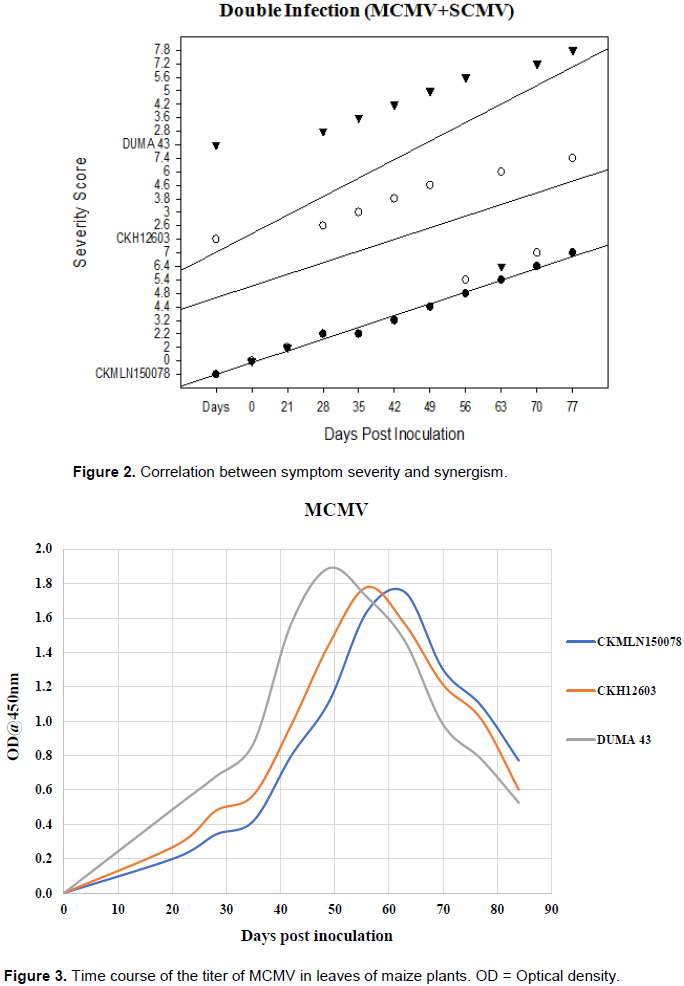
Correlation between symptom severity and synergism
As the days passed, the magnitude of the crop’s symptoms increased significantly, as seen in Figure 1. The severity score for the three hybrids was low during the first 10 days post inoculation (dpi), but increased substantially as the days progressed. The highest severity scores were recorded between 63-84 dpi, with DUMA 43 exhibiting the highest severity scores (6.5), followed by CKH12603 (4.5) and CKMLN150078 (4.0), respectively (Figure 2).
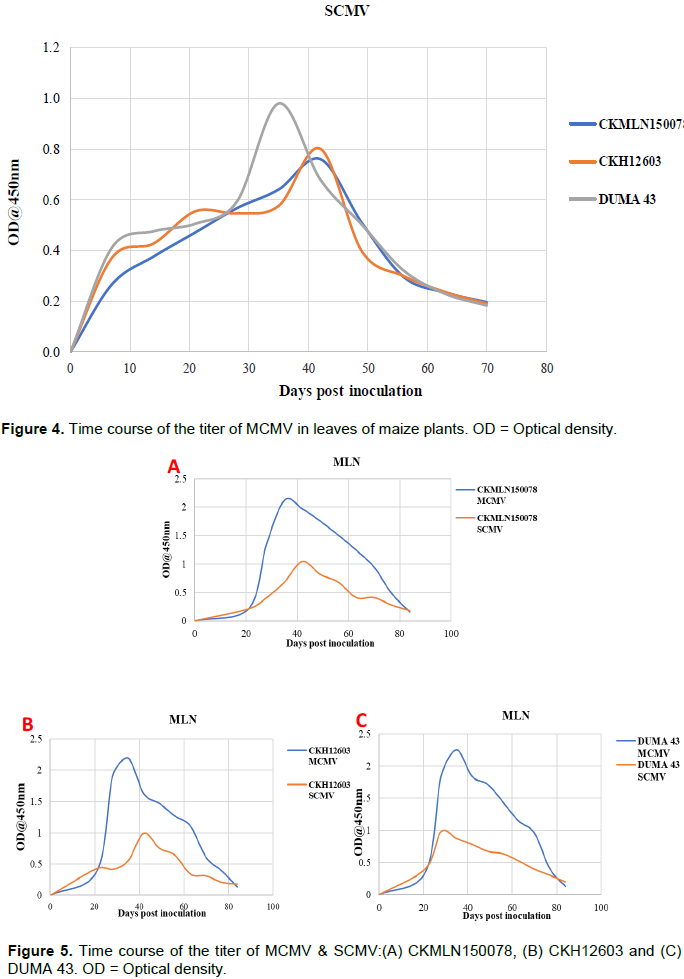
Titer of MCMV and SCMV in singly inoculated maize plants
The MCMV titer in the three hybrids was found to follow a normal curve (Figure 3). During the first 10 dpi, the viral titer was low; however, as the days passed, the viral titer increased dramatically. At different dpi levels, the three hybrids had higher viral titers, with Duma 43 having the highest viral titer (0.98) compared to the other two hybrids. However, there was a decrease in viral titers among the three hybrids between 50, 56, and 64 dpi (Figure 3). The viral titer for the three hybrids of SCMV inoculated maize plants followed a normal curve, similar to the MCMV inoculated maize plants described above (Figure 3). SCMV inoculated alone hybrids, on the other hand, had lower viral titers than MCMV inoculated alone hybrids, which had higher OD values (Figure 4). Between 0 and 35 dpi, the viral titers of the three hybrids increased significantly, with Duma 43 recording a higher viral titer at 35 dpi than the other two hybrids at 42 dpi (Figure 4). The SCMV titer in the three hybrids gradually decreased after that.
Titers of MCMV and SCMV in co-infected maize plants
There was a slight variation in the titers of the two viruses during the first 10 dpi in the doubly infected maize plants with MCMV + SCMV. However, as the days passed, there was a significant increase in MCMV titers compared to SCMV titers (Figure 5). Consequently, MCMV titer values in co-infected maize plants were higher than MCMV titer values in singly infected maize plants, whereas SCMV titer values were less similar. Moreover, the titers of the Duma 43 hybrid were higher than the titers of the other two hybrids (Figure 5). Furthermore, it was observed that Duma 43 took a shorter period of 21 dpi for its titer to reach its maximum, as opposed to the singly infected, which required 35 dpi (Figure 5).
The findings of this study have revealed that the viral titers of the two viruses in both singly and co-infection, followed a normal curve. The normal virus titer curve could be ascribed to the plant virus infection cycle, in which the virus must initially overcome pre-existing chemical and physical barriers in plants (Pallas and Garci, 2011). Therefore, the low titer of the two viruses during the first 10dpi could be attributed to low virus replication, cell-to-cell movement, as well as the long-distance movement of the virus through the vascular tissues of the plant as the virus is overcoming the defensive mechanism of the maize plant (Syller and Grupa, 2016). On other hand, the rapid increase in viral titer after the 21dpi is as result of a high rate of virus multiplication and translocation of virus particles throughout the plant cells through production of RNA-silencing suppressors, which interfere with the maize plant physiological processes which depend on RNA-silencing; hence enhancing the pathogenicity of the virus (Roth et al., 2004). Furthermore, it was noted that the viral titers of the two viruses declined after attaining the maximum concentration/symptom development. The decline in viral titer was linked to increased virulence of the viruses which resulted in cells death of the plant which in turn resulted in a decline in virus particles as viruses reside within the cells of the plants (Santi et al., 2006).
Also, it was observed that the viral titers of DUMA 43 in both singly and co-infections were higher as compared to the other two hybrids. The high viral titer could be due to the susceptibility nature of the variety to MLN disease as previously reported by Mbega et al. (2016) in a review paper. They reported that in a susceptible host the virus particles can move between the cells through the plasmodesmata and the whole plant through the phloem, thus colonizing the plant and eventual expression of the MLN symptoms; while in a resistant host, the virus colonization is only sparingly possible hence no expression of symptoms.
Furthermore, it was also observed that the viral titers of SCMV in both singly and co-infected maize plants remained constant amongst the three hybrids; while the viral titers of MCMV in co-infected maize (MCMV+SCMV) increased significantly as compared to the singly infected plants. Such findings are consistent with previous reports byAwata et al. (2019)who reported that concentration of SCMV in the mixed infections remained constant. The constant concentration of SCMV in a mixed infection could be associated to viral proteins such as P1 and VPg, which are not strong enhancers of replication and movement (Awata et al., 2019). On the other hand, the observed increase in the concentration of MCMV in co-infected maize plants (as compared to the singly infected) could be traced to the ability of SCMV to suppress regulatory systems of the maize plant that would normally limit concentrations of MCMV in a cell; thus, allowing easy transmission of the MCMV, and hence increased symptom severity (Xia et al., 2016). Rajamäki and Valkonen (2009), in their report, found that potyviruses contain two important genes namely: helper component gene (HC-pro) and nuclear inclusions protein gene. These two genes reduce the capacity of the maize plant to inhibit the replication of MCMV. Moreover, it has been reported that SCMV VPg enhances cell to cell movement and long-distance movement of its virus particles as well as those of MCMV (Scheets, 1998). Furthermore, VPg also has been reported to suppress the post-transcriptional gene (PTGS) of the host plant thereby allowing successful colonization of the plant by the virus (Mbega et al., 2016). Such mechanisms, therefore, explain why there were increased titers of MCMV in a mixed infection, especially between 35-42dpi’s while the titers of SCMV in both infections remained more or less the same.
Also, it's worth noting that, between 35-42 dpi, there was a significant increase in the viral titers of MCMV while that of SCMV was less constant; and thereafter, there was a decline in viral titers of the two viruses. The observed increase in the viral titers of MCMV in co-infection with SCMV as compared to single infection by MCMV alone is hypothesized to be due to the ability of the SCMV to suppress regulatory systems that would normally limit MCMV concentrations in a cell allowing easy transmission of the MCMV and increasing the symptom severity (Mbega et al., 2016). SCMV has been reported to promote its multiplication and movement, as well as suppression of the host plant defense mechanisms (Mbega et al., 2016). The significant increase in the viral titers of MCMV is termed unilateral synergism (Scheets, 1998). The period is a signal of cumulative effects of the synergistic interaction between the two viruses and resulting in the overrunning of the host plant defense barriers (Awata. et al., 2019). At such a stage, there is increased symptom severity in the plant (Xia et al., 2016), which eventually results in plant death. The period serves as a pointer that synergism between the two viruses was at its peak and therefore it is an indication that the synergism between the viruses had started as early as 21dpi, since there was a rapid increase in the viral titers of the two viruses. This is an important observation for recommendation to maize breeders to take cognizance of such a period in breeding for tolerance/resistance against MLN. The findings also serve as a significant recommendation to seed companies that specialize in seed multiplication, as well as small-scale and commercial farmers, to use recommended insecticides to control aphid and thrip species between 7 and 14 days after emergence, before synergism develops. This practice has been shown to reduce MLN incidence and severity by reducing the population of thrip species responsible for MCMV transmission (Ngala et al., 2018).
This study has clearly elucidated that SCMV plays a significant role in the multiplication and movement of MCMV particles in the plant. It has also been observed that the synergistic interaction between the two viruses in the maize plant is at its peak between 35-42dpi. The determination of the synergistic period is critical for plant pathologists and plant breeders in their effort to develop maize varieties that can avoid this period, overcoming synergism establishment and increasing yield.
The authors have not declared any conflict of interests.
The authors sincerely appreciate the International Maize and Wheat Improvement Centre (CIMMYT) for funding this study.
REFERENCES
|
Adams IP, Harju VA, Hodges T, Hany U, Skelton A, Rai S, Boonham N (2014). First report of maize lethal necrosis disease in Rwanda. New Disease Reports 29:22.
Crossref
|
|
|
|
Awata L, Ifie E, Tongoona P, DanquahE, Jumbo BMG, Suresh LM (2019). Maize lethal necrosis and the molecular basis of variability in concentrations of the causal viruses in co-infected maize plant. Journal of General and Molecular Virology 9(1):1-19.
|
|
|
|
|
Groote HD, Oloo F, Tongruksawattana S, Das B (2016). Community-survey based assessment of the geographic distribution and impact of maize lethal necrosis (MLN) disease in Kenya. Crop Protection 82:30-35.
Crossref
|
|
|
|
|
Isabirye BE, Rwomushana I (2016). Current and future potential distribution of maize chlorotic mottle virus and risk of maize lethal necrosis disease in Africa. Crop Breeding and Applied Biotechnology 5(2):215-228.
Crossref
|
|
|
|
|
Kagoda F, Gidoi R, Isabirye BE (2016). Status of maize lethal necrosis in eastern Uganda. African Journal of Agricultural Research 11(8):652-660.
Crossref
|
|
|
|
|
Karanja J, Derera J, Gubb, A, Mugo S, Wangai A (2018). Response of Selected Maize Inbred Germplasm to Maize Lethal Necrosis Disease and Its Causative Viruses (Sugarcane Mosaic Virus and Maize Chlorotic Mottle Virus ) in Kenya. The Open Agriculture 12:215-226.
Crossref
|
|
|
|
|
Leitich RK, Korir JC, Muoma JO, Wangai A, Bong K, Johal G, Loesch-Fries S (2020). Molecular characterization of viruses causing maize lethal necrosis disease in South-Rift region, Kenya. International Journal of Genetics and Molecular Biology 12(2):71-77.
|
|
|
|
|
Mahuku G, Lockhart BE, Wanjala B, Jones MW, Kimunye JN, Stewart LR, Redinbaugh MG (2015). Maize Lethal Necrosis ( MLN ), an Emerging Threat to Maize-Based Food Security in Sub-Saharan Africa. International Journal of Current Microbiology and Applied Sciences 5(6):85-96.
|
|
|
|
|
Makone SM, Menge D, Basweti E (2014). Impact of Maize Lethal Necrosis Disease on maize yield:A case of Kisii, Kenya. International Journal of Agricultural Extension 2(3):211-218.
|
|
|
|
|
Mbega ER, Ndakidemi PA, Mamiro DP, Mushongi AA, Kitenge KM, Ndomba OA (2016). Role of Potyviruses in Synergistic Interaction Leading to Maize Lethal Necrotic Disease on Maize. International Journal of Current Microbiology and Applied Sciences 5(6):85-96.
Crossref
|
|
|
|
|
Pallas V, Garc? JA (2011). How do plant viruses induce disease? Interactions and interference with host components. General Virology 92(12):2691-2705.
Crossref
|
|
|
|
|
Ngala RM, Kilalo DC, Miano D, Mukunya D (2018). Spatial and temporal spread of maize lethal necrosis disease causing viruses and their vectors within the field. African Journal of Rural Development 3(MCMV):787-798.
|
|
|
|
|
Roth BM, Pruss GJ, Vance VB (2004). Plant viral suppressors of RNA silencing Plant viral suppressors of RNA silencing. Virus Research 102(1):97-108.
Crossref
|
|
|
|
|
Santi L, Huang Z, Mason H (2006). Virus-like particles production in green plants. Methods 40(1):66-76.
Crossref
|
|
|
|
|
Scheets K (1998). Maize Chlorotic Mottle Machlomovirus and Wheat Streak Mosaic Rymovirus Concentrations Increase in the Synergistic Disease Corn Lethal Necrosis 38:28-38.
Crossref
|
|
|
|
|
Syller J, Grupa A (2016). Antagonistic within-host interactions between plant viruses?: molecular basis and impact on viral and host fitness. Molecular Plant Pathology 17(5):769-782.
Crossref
|
|
|
|
|
Wangai AW, Redinbaugh MG, Kinyua ZM, Miano DW, Leley PK, Kasina M, Mahuku G, Scheets K, Jeffers D (2012). Zea mays. Plant Disease 96(10):1582.
Crossref
|
|
|
|
|
Xia Z, Zhao Z, Chen L, Li M, Zhou T, Deng C, Zhou Q (2016). Synergistic infection of two viruses MCMV and SCMV increases the accumulations of both MCMV and MCMV-derived siRNAs in maize. Scientific Reports 6. Article number 20520.
Crossref
|
|