Full Length Research Paper

ABSTRACT
INTRODUCTION
MATERIALS AND METHODS
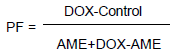
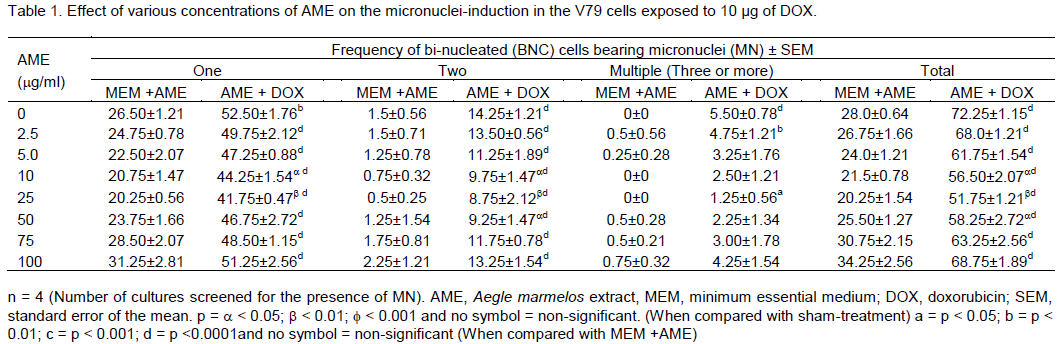
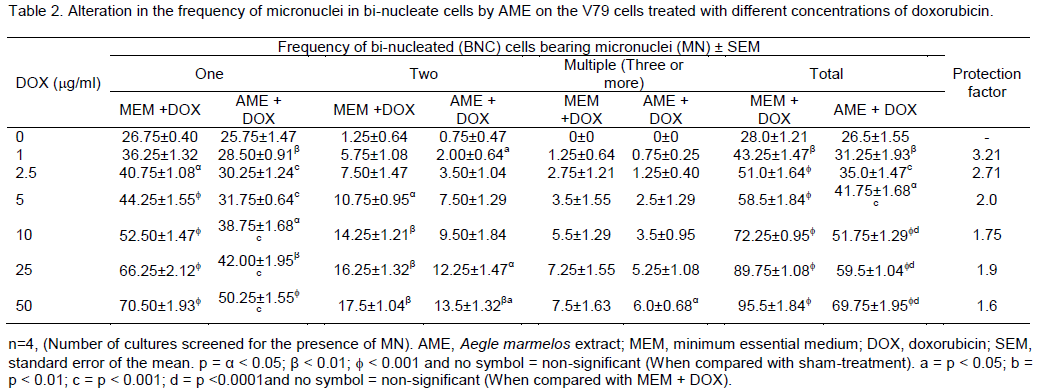
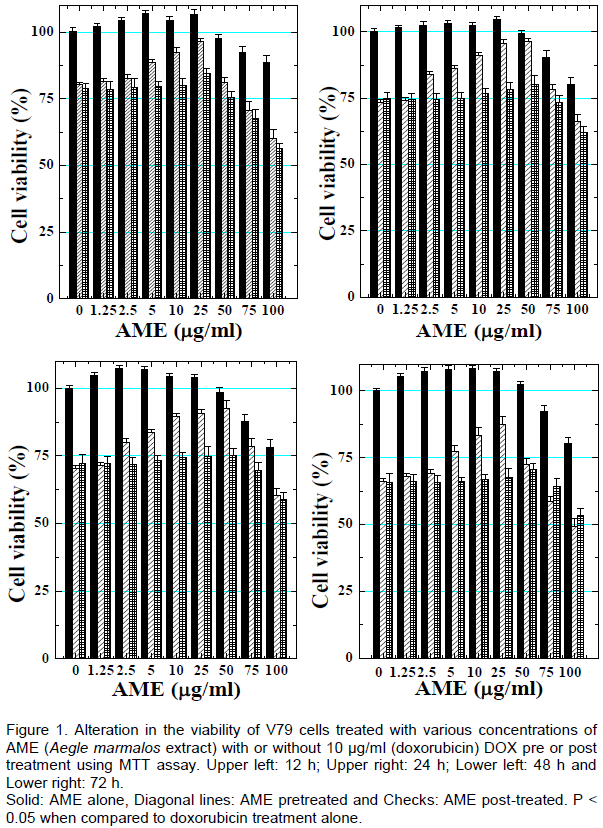
RESULTS
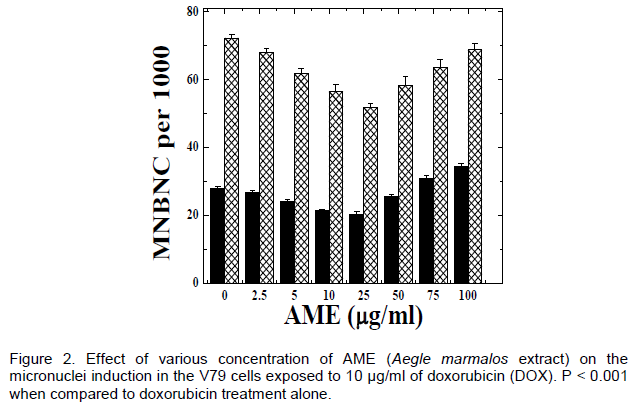
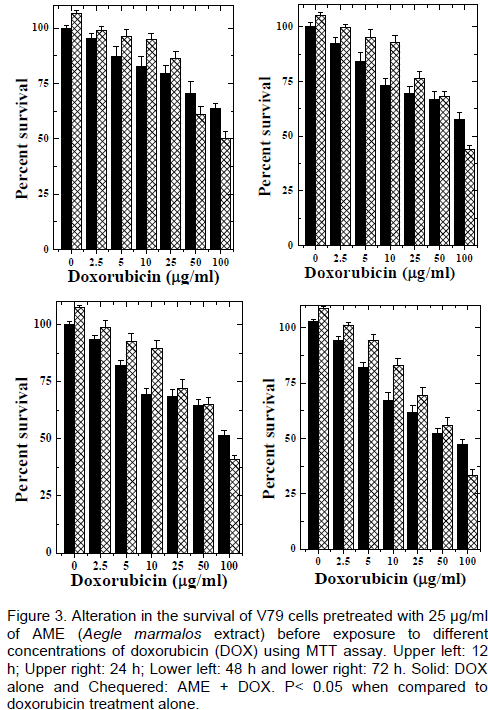
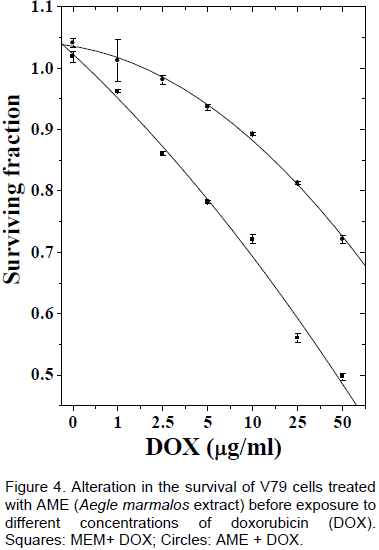
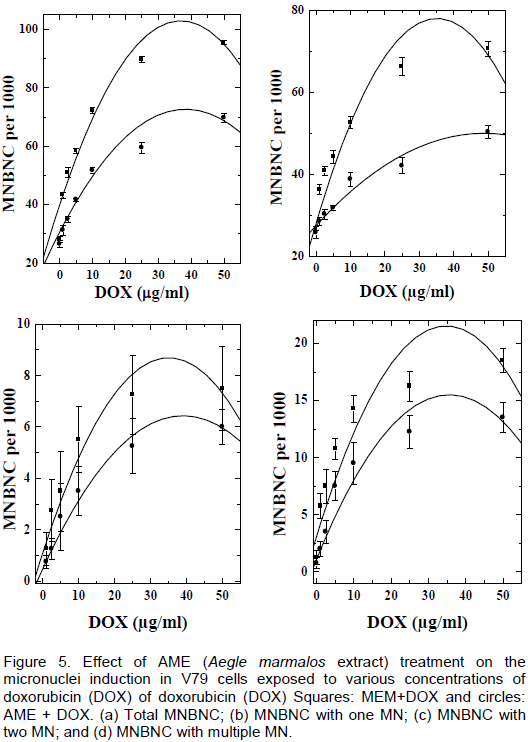
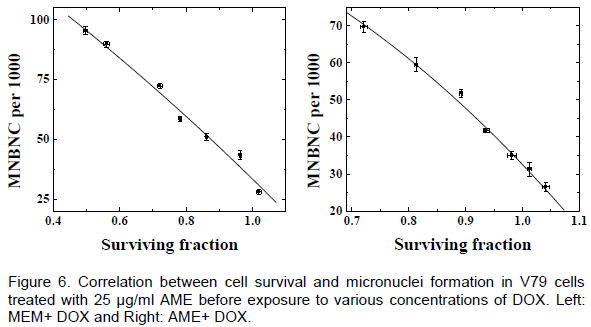
DISCUSSION
CONCLUSION
CONFLICT OF INTERESTS
The authors did not declare any conflict of interest.
ACKNOWLEDGEMENTS
The authors are thankful to the Indian council of Medical Research, Govt. of India for financial support vide grant No. 45/2/2004/BMS/TRM to carry out this study.
REFERENCES
|
Abraham R, Basser RL, Green MD (1996). A risk-benefit assessment of anthracycline antibiotics in antineoplastic therapy. Drug Safety 15:406-429. |
|
|
|
|
|
Al-Harbi MM (1993). Effect of captopril on the cytological and biochemical changes induced by adriamycin. Food Chem. Toxicol. 31: 209-212. |
|
|
|
|
|
Al-Shabanah OA (1993). Inhibition of adriamycin-induced micronuclei by desferrioxamine in Swiss albino mice. Mutat. Res. 301:107-111. |
|
|
|
|
|
Antunes LM, Takahashi CS (1998). Effects of high doses of vitamins C and E against doxorubicin-induced chromosomal damage in Wistar rat bone marrow cells. Mutat. Res. 419:137-143. |
|
|
|
|
|
Antunes LM, Takahashi CS (1999). Protection and induction of chromosomal damage by vitamin C in human lymphocyte cultures.Teratog. Carcinog. Mutagen. 19:53-59. |
|
|
|
|
|
Babudri N, Pani B, Tamaro M, Monti-Bragadin C, Zunino F (1984). Related Articles, Mutagenic and cytotoxic activity of doxorubicin and daunorubicin derivatives on prokaryotic and eukaryotic cells. Br. J. Cancer 50:91-96. |
|
|
|
|
|
Baker ES, Connor TH (1996). Monitoring occupational exposure to cancer chemotherapy drugs. Am. J. Health Syst. Pharmacol. 53: 2713–2723. |
|
|
|
|
|
Bardeleben RV, Dunkern T, Kaina B, Fritz G (2002). The HMG-CoA reductase inhibitor lovastatin protects cells from the antineoplastic drugs doxorubicin and etoposide. Int. J. Mol. Med. 10: 473-479. |
|
|
|
|
|
Bean CL, Armstrong MJ, Galloway SM (1992). Effect of sampling time on chromosome aberration yield for 7 chemicals in Chinese hamster ovary cells. Mutat. Res. 265:31-44. |
|
|
|
|
|
Bhuyan BK, Zimmer DM, Mazurek JH, Trzos RJ, Harbach PR, Shu VS, Johnson MA (1983). Related Articles, Comparative genotoxicity of adriamycin and menogarol, two anthracycline antitumor agents. Cancer Res. 43: 5293-5297. |
|
|
|
|
|
Bogdanović G, Kojić V, Dordević A, Canadanović-Brunet J, Vojinović-Miloradov M, Baltić VV. (2004) Modulating activity of fullerol C60(OH)22 on doxorubicin-induced cytotoxicity.Toxicol. InVitro. 18(5):629-637. |
|
|
|
|
|
Bonner JA, Lawrence (1989). TS Protection of doxorubicin cytotoxicity by cycloheximide. Int. J. Radiat. Oncol. Biol. Phys. 16:1209-1212. |
|
|
|
|
|
Boucher R, Livingston GK, Que Hee SS (1993). In vitro micronucleus bioassay of human peripheral lymphocytes for adriamycin in the presence of cyclophosphamide and urines of patients administered anticancer drugs. Environ. Mol. Mutagen. 21:372-382. |
|
|
|
|
|
Carter SK (1975). Adriamycin – A review. J. Natl. Cancer Inst. 55:1265-1274. |
|
|
|
|
|
Chen SH, Chan NL, Hsieh TS (2013). New mechanistic and functional insights into DNA topoisomerases .Annu Rev Biochem. 82:139-147 |
|
|
|
|
|
De Graff WG, Myers Jr LS, Mitchell JB, Hahn SM (2003). Protection against Adriamycin cytotoxicity and inhibition of DNA topoisomerase II activity by 3, 4-dihydroxybenzoic acid. Int. J. Oncol. 23:159-163. |
|
|
|
|
|
Delvaeye M, Verovski V, De Neve W, Storme G (1993). DNA breakage, cytotoxicity, drug accumulation and retention in two human ovarian tumor cell lines AZ224 and AZ364 treated with adriamycin, modulated by verapamil. Anticancer Res. 13: 1533-1538. |
|
|
|
|
|
Dhawan A, Kayani MA, Parry JM, Parry E, Anderson D (2003). Aneugenic and clastogenic effects of doxorubicin in human lymphocytes. Mutagenesis 18:487-490. |
|
|
|
|
|
Dulout FN, Larramendy ML, Olivero OA (1981). Effect of caffeine on the frequency of chromosome aberrations induced in vivo by triethylenemelamine (TEM) and adriamycin (ADR) in mice. Mutat. Res. 82:295-304. |
|
|
|
|
|
Fenech M, Morley AA (1985). Measurement of micronuclei in lymphocytes. Mutat. Res. 147:29-36. |
|
|
|
|
|
Fenech M, Kirsch-Volders M, Natarajan AT, Surralles J, Crott JW, Parry J, Norppa H, Eastmond DA, Tucker JD, Thomas P (2011) Molecular mechanisms of micronucleus, nucleoplasmic bridge and nuclear bud formation in mammalian and human cells. Mutagenesis. (1):125-32. |
|
|
|
|
|
Gewirtz DA (1999). A critical evaluation of the mechanisms of action proposed for the antitumor effects of the anthracycline antibiotics adriamycin and daunorubicin. Biochem. Pharmacol. 57:727-741. |
|
|
|
|
|
Guano F, Pourquier P, Tinelli S, Binaschi M, Bigioni M, Animati F, Manzini S, Zunino F, Kohlhagen G, Pommier Y, Capranico G (1999). Topoisomerase poisoning activity of novel disaccharide anthracyclines. Mol. Pharmacol. 56:77-84. |
|
|
|
|
|
Gumulec J, Fojtu M, Raudenska M, Sztalmachova M, Skotakova A, Vlachova J, Skalickova S, Nejdl L, Kopel P, Knopfova L0, Adam V, KizekR,Stiborova M, Babula P, Masarik M (2014) Modulation of induced cytotoxicity of doxorubicin by using apoferritin and liposomal cages. Int. J. Mol. Sci. 15(12):22960-22977 |
|
|
|
|
|
Heddle JA, Hite M, Kirkhart B, Mavournin K, MacGregor JT, Newell GW, Salamone MF (1983). The induction of micronuclei as a measure of genotoxicity. A report of the U.S. Environmental Protection Agency Gene-Tox. Program. Mutat. Res. 123:61-118. |
|
|
|
|
|
Helbig R, Speit G (1995). The pattern of adriamycin-induced mutations in V-E5 Chinese hamster cells with chromosomal instability. Mutat. Res. 348:1-6. |
|
|
|
|
|
Hozayen WG (2012). Effect Of Hesperidin and Rutin On Doxorubicin Induced Testicular Toxicity in Male Rats. Int. J. Food. Nutrit.Sci., 1(1):31-42. |
|
|
|
|
|
Iliakis G, Lazar W (1987). Reduction by caffeine of adriamycin-induced cell killing and DNA damage in Chinese hamster cells: correlation with modulation in intracellular adriamycin content. Cancer Res. 47: 2224-2229. |
|
|
|
|
|
Jagetia GC, Baliga MS (2002). Syzygium cumini (Jamun) reduces the radiation-induced DNA damage in the cultured human peripheral blood lymphocytes: a preliminary study. Toxicol. Lett. 132: 19-25. |
|
|
|
|
|
Jagetia GC, Reddy TK (2014) The grape fruit flavononenaringin protects mice against doxorubicin-induced cardiotoxicity. J. Mol. Biochem. 3:34-49. |
|
|
|
|
|
Jagetia GC (2008). Cardioprotection by Bael. Recent Progress in Medicinal Plants: Phytopharmacology and Therapeutic values. 20 Edited by Govil JN, Singh VK, Mishra SK Studium Press LLC, Houston USA pp 147-162. |
|
|
|
|
|
Jagetia GC, Adiga SK (1998). Influence of vindesine exposure on the micronucleus formation and cell survival in V79 cells. Mutat. Res. 421:55-63. |
|
|
|
|
|
Jagetia GC, Aruna R (1997). The herbal preparation abana protects against radiation-induced micronuclei in mouse bone marrow. Mutat. Res. 393:157-163. |
|
|
|
|
|
Jagetia GC, Aruna R (2000). Correlation between cell survival and micronuclei-induction in HeLa cells treated with adriamycin after exposure to various doses of gamma-radiation. Toxicol. Lett. 115: 183-193. |
|
|
|
|
|
Jagetia GC, Ganapathi NG (1989). Inhibition of clastogenic effect of radiation by Liv. 52 in the bone marrow of mice. Mutat. Res. 224:507-510. |
|
|
|
|
|
Jagetia GC, Nayak V (1996). Micronuclei-induction and its correlation to cell survival in HeLa cells treated with different doses of adriamycin. Cancer Lett. 110: 123-128. |
|
|
|
|
|
Jagetia GC, Nayak V (2000). Effect of doxorubicin on cell survival and micronuclei formation in HeLa cells exposed to different doses of gamma-radiation. Strahlenther Onkol. 176:422-428. |
|
|
|
|
|
Jagetia GC, Venkatesh P (2005). Radioprotection by oral administration of Aegle marmelos (L.) Correa in vivo. J. Environ. Pathol. Toxicol. Oncol. 24: 315-332. |
|
|
|
|
|
Jagetia GC, Venkatesh P (2007). Inhibition of radiation-induced clastogenicity by Aegle marmelos (L.) correa in mice bone marrow exposed to different doses of gamma-radiation. Hum. Exp. Toxicol. 26: 111-24. |
|
|
|
|
|
Jagetia GC, Venkatesha VA (2005). Effect of mangiferin on radiation-induced micronucleus formation in cultured human peripheral blood lymphocytes. Environment Molec. Mutagen. 46:12-21. |
|
|
|
|
|
Jagetia GC, Venkatesh P, Baliga MS (2003). Evaluation of the radioprotective effect of Aegle marmelos (L.) Correa in cultured human peripheral blood lymphocytes exposed to different doses of gamma-radiation: a micronucleus study. Mutagenesis 18: 387-393. |
|
|
|
|
|
Jagetia GC, Venkatesh P, Baliga MS (2004). Evaluation of the radioprotective effect of bael leaf (Aegle marmelos) extract in mice. Int. J. Radiat. Biol. 80:281-290. |
|
|
|
|
|
Kato FH, Viana NI, Santini CB, de Souza CG, Veneziani RC, Ambrósio SR, Tavares DC (2012). Assessment of the in vitro and in vivo genotoxic and antigenotoxic effects of pimaradienoic acid in mammalian cells. Mutat. Res. 749(1-2):87-92. |
|
|
|
|
|
Kinghorn AD, Farnsworth NR, Soejarto DD, Cordell GA, Swanson SM, Pezzuto SM, Wani MC, Wall ME, Oberlies NH, Kroll DJ, Kramer RA, Rose WC, Vite GD, Fairchild CR, Peterson RW, Wild R (2003). Novel strategies for the discovery of plant-derived anticancer agents. Pharmaceut. Biol. 41 Suppl: 53-67. |
|
|
|
|
|
Kirsch-Volders M, Sofuni T, Aardema M, Albertini S, Eastmond D, Fenech M, Ishidate M Jr, Kirchner S, Lorge E, Morita T, Norppa H, Surrallés J, Vanhauwaert A, Wakata A. (2003) Report from the in vitro micronucleus assay working group. Mutat Res. 540(2):153-163. |
|
|
|
|
|
Kiyomiya K, Matsuo S, Kurebe M (2001). Differences in intracellular sites of action of Adriamycin in neoplastic and normal differentiated cells. Cancer Chemother. Pharmacol. 47:51-56. |
|
|
|
|
|
Korina LG, Afanas'ev IB (1997). Antioxidant and chelating properties of flavonoids. Adv. Pharmacol. 38:151-163. |
|
|
|
|
|
López-Alarcón C, Denicola A (2013). Evaluating the antioxidant capacity of natural products: A review on chemical and cellular-based assays. Analytica Chimica Acta 763:1-10. |
|
|
|
|
|
Majer BJ, Laky B, Knasmuller S, Kassie F (2001). Use of the micronucleus assay with exfoliated epithelial cells as a biomarker for monitoring individuals at elevated risk of genetic damage and in chemoprevention trials. Mutat. Res. 489(2-3): 147-172. |
|
|
|
|
|
Martin WM and McNally NJ (1980). Cytotoxicity of adriamycin to tumour cells in vivo and in vitro. Br. J. Cancer 42:881-889. |
|
|
|
|
|
Minotti G, Recalcati S, Menna P, Salvatorelli E, Corna G, Cairo G (2004). Doxorubicin cardiotoxicity and the control of iron metabolism: quinone-dependent and independent mechanisms. Methods Enzymol. 378: 340-361. |
|
|
|
|
|
Mosmann T (1983). Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Meth. 65:55-63. |
|
|
|
|
|
Nadkarni AK (1976). Indian Materia Medica, Ed 3. Popular Press Ltd. Mumbai India. |
|
|
|
|
|
Nitiss JL (2009) Targeting DNA topoisomerase II in cancer chemotherapy.Nat. Rev. Cancer 9(5):338-50. |
|
|
|
|
|
Pendleton M, Lindsey RH Jr, Felix CA, Grimwade D, Osheroff N (2014). Topoisomerase II and leukemia. Ann N Y Acad Sci. 1310:98-110. |
|
|
|
|
|
Pérez-Arnaiz C, Busto N, Leal JM, García B (2014). New insights into the mechanism of the DNA/doxorubicin interaction. J Phys Chem B. 118(5):1288-95. |
|
|
|
|
|
Pletz N, Schön M, Ziegelbauer K, Emmert S, Liu N, Dobbelstein M, Schön MP (2012). Doxorubicin-induced activation of NF-κB in melanoma cells is abrogated by inhibition of IKKβ, but not by a novel IKKα inhibitor. Exp. Dermatol. 21(4):301-304. |
|
|
|
|
|
Pommier Y, Schwartz RE, Zwelling LA, Kohn KW (1985). Effects of DNA intercalating agents on topoisomerase II induced DNA strand cleavage in isolated mammalian cell nuclei. Biochemistry 24:6406-6410. |
|
|
|
|
|
Powis G (1989). Free radical formation by antitumor quinones. Free Radical Biol. Med. 6:63-101. |
|
|
|
|
|
Puck TT, Marcus PI (1955). A rapid method for viable cell titration and clone production with HeLa cells in tissue culture. The use of X-irradiated cells to supply conditioning factors. Proc. Natl. Acad. Sci. USA 41:432-437. |
|
|
|
|
|
Rahman S, Parvin R (2014) Therapeutic potential of Aegle marmelos (L.)-An overview Asian Pac J Trop Dis 4(1):71-77. |
|
|
|
|
|
Rastogi RM, Mehrotra BN (1990). Compendium of Indian medicinal plants (Central drug research Institute Lucknow India) 1: 388-389. |
|
|
|
|
|
Rice-Evans CA, Miller NJ, Bolwell PG, Bramley PM, Pridham JB (1995). The relative antioxidant activities of plant-derived polyphenolic flavonoids. Free Radic. Res. 22:375-383. |
|
|
|
|
|
Russell NS, Arlett CF, Bartelink H, Begg AC (1995). Use of fluorescence in situ hypbridization to determine the relationship between chromosome aberrations and cell survival in eight human fibroblast strains. Int. J. Radiat. Biol. 68: 185-196. |
|
|
|
|
|
Sato Y, Sasaki N, Saito M, Endo N, Kugawa F, Ueno A (2015) Luteolin attenuates Doxorubicin-induced cytotoxicity to mcf-7 human breast cancer cells. Biol. Pharm. Bull. 38(5):703-709. |
|
|
|
|
|
Schneider E, Hsiang Y, Liu LF (1990). DNA topoisomerases as anticancer drug targets. Adv. Pharmacol. 21:149-183. |
|
|
|
|
|
Shibamoto Y, Streffer C, Fuhrmann C, Budach V (1991). Tumor radiosensitivity prediction by the cytokinesis-block micronucleus assay. Radiat. Res. 128:293-300. |
|
|
|
|
|
Sugamura K, Keaney J F Jr. (2011). Reactive oxygen species in cardiovascular disease. Free Radic. Med. 51:978-992. |
|
|
|
|
|
Suter W, Brennand J, McMillan S, Fox M (1980). Relative mutagenicity of antineoplastic drugs and other alkylating agents in V79 Chinese hamster cells, independence of cytotoxic and mutagenic responses. Mutat. Res. 73: 171-181. |
|
|
|
|
|
Tacar O, Sriamornsak P, Dass CR (2013). Doxorubicin: an update on anticancer molecular action, toxicity and novel drug delivery systems. J Pharm Pharmacol 65(2) 157-170 |
|
|
|
|
|
Tewey KM, Rowe TC, Yang L, Halligan BD, Liu LF (1984). Adriamycin-induced DNA damage mediated by mammalian DNA topoisomerase II. Science 226: 466–468. |
|
|
|
|
|
Van Acker S, Van den Berg DJ, Tromp MNJL, Griffioen DH, Van Bennekom WP, Van der Vijgh WJF, Bast A (1996). Structural aspects of antioxidant activity of flavonoids. Free Radic. Biol. Med. 20: 331-342. |
|
|
|
|
|
Venkatesh P, Shantala B, Jagetia GC, Rao KK, Baliga MS (2007). Modulation of doxorubicin-induced genotoxicity by Aegle marmelos in mouse bone marrow: a micronucleus study. Integr. Cancer Ther. 6:42-53. |
|
|
|
|
|
Wandl EO, Ono K, Kain R, Herbsthofer T, Hienert G, Hobarth K (1989). Linear correlation between surviving fraction and the micronucleus frequency. Int. J. Radiat. Biol. 56: 771-775. |
|
|
|
|
|
Wang S, Kotamraju S, Konorev E, Kalivendi S, Joseph J, Kalyanaraman B (2002). Activation of nuclear factor-kappaB during doxorubicin-induced apoptosis in endothelial cells and myocytes is pro-apoptotic: the role of hydrogen peroxide. Biochem. J. 367: 729-740. |
|
|
|
|
|
Weiss JF, Landauer MR (2003). Protection against ionizing radiation by antioxidant nutrients and phytochemicals. Toxicology 189:1-20. |
|
|
|
|
|
Wu X, Mao Y (2012). Effect of catalpol on doxorubicin-induced cytotoxicity in H9c2 cells. J. Med. Plants Res. 6(5):849-854. |
|
|
|
|
|
Yates LR, Campbell PJ (2012). Evolution of the cancer genome. Nature Rev. Genet. 13:795-806. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


