ABSTRACT
The first report of Maize lethal necrosis (MLN) disease in Africa was in Bomet, Kenya, nine years ago. It has since spread in East and Central African (ECA) countries, causing massive yield losses. Currently, 90% of the preferred commercial maize germplasm grown by farmers in Kenya is susceptible to MLN disease. As such, the disease has continued to pose a serious challenge to food security in the ECA region. This study sought to characterize the MLN, causing viruses present in the maize leaf samples collected from the South-Rift region. Using total RNA extracted from 60 leaf samples collected from Bomet, Kericho, and Kisumu Counties, reverse transcription-polymerase chain reaction (RT-PCR) was carried out. The PCR products with the strongest bands were purified and sequenced using the Sanger sequencing technique. The results showed that samples from the three counties were positive for maize chlorotic mottle virus (MCMV) (MH645622 and MH645621) and sugarcane mosaic virus (SCMV) (MH645623, MH645624 and MH645625) and negative to wheat streak mosaic virus (WSMV). The coat protein (CP) sequences of MCMV isolates were closely related to the sequences of MCMV isolates, which had been previously reported from Eastern and Sub-Saharan Africa. For the CP sequences of SCMV isolates, only one sequence of the isolate KCO59 was similar to the sequence of a Kenyan isolate (JX286708). Sequences of isolates KCO5 and KCO24 were not identical to those of the Kenyan isolate (JX286708). Based on these results, in the surveyed counties, isolates of SCMV are genetically diverse, while those of MCMV are not. There exists a new variant of SCMV, which appears to be the main potyvirus in synergism with MCMV in causing MLN disease in Kenya.
Key words: Maize lethal necrosis (MLN), maize chlorotic mottle virus (MCMV), sugarcane mosaic virus (SCMV), wheat streak mosaic virus (WSMV).
Maize lethal necrosis (MLN) disease has emerged as one of the major threats to food security in Sub-Saharan Africa as maize is a staple food crop (Mbega et al., 2016). The disease has become a significant concern in
Eastern Africa, having been first reported in Bomet County in the South-Rift region of Kenya in 2011 (Wangai et al., 2012). Since then, it has spread to other maize-growing areas in Central, Nyanza, Western, South, and North-Rift regions of Kenya (Davis, 2012). The disease has also been reported in other countries in East Africa where maize is an outstanding staple food. These countries include; Rwanda (Adams et al., 2014), Democratic Republic of Congo (DRC) (Lukanda et al., 2014), Tanzania (Kiruwa et al., 2016)Uganda (Kagoda et al., 2016)and more recently from South Sudan and Ethiopia (Mahuku et al., 2015). Yield losses as high as 59% or more than 300,000 tons have been reported in moist transitional zones, mostly in Western Kenya (De Groote et al., 2016)due to MLN disease. The disease has been reported to be a result of synergistic interaction between Maize chlorotic mottle virus (MCMV) with any potyvirus such as Sugarcane mosaic virus (SCMV), Maize dwarf mosaic virus (MDMV) or Wheat streak mosaic virus (WSMV) (Isabirye and Rwomushana, 2016). The synergistic interactions have been reported to be more pronounced and result in severe damage that usually kills the infected plant (Makone et al., 2014). Due to such magnitude of loss posed by the disease, it has discouraged some farmers from planting maize in the second season (Kamau and Snipes, 2014), a situation that is likely to affect maize production in the region.
Though there are commercial varieties that have already been released to farmers with appreciable tolerance to MLN, most farmers have little knowledge of how the disease spread and its control measures (Mahuku et al., 2015). Due to such inadequacies, most of the farmers practice continuous maize cropping, a practice that has been reported as one of the factors contributing to the build-up of the virus inoculum (Mezzalama, 2015). With this in mind, there is a need to characterize the virus isolates from the South-Rift region to identify the virus causing MLN disease in this region and assess their diversity. Therefore, this study sought to isolate viruses from the maize leaf samples collected from the South-Rift region in Kenya and sequence their coat protein gene.
Sample collection
Maize leaf samples were collected between July and August 2016 in three counties of the South-Rift region of Kenya; Bomet, Kericho, and Kisumu. These counties were selected as MLN occurrence was first reported in September 2011 in Longisa in Bomet (Wangai et al., 2012), which is in Kenya's South-Rift region. Random sampling was used to select the maize farms and sampling sites. Ten farms were randomly selected from each county, and in each farm, five samples were collected. Sampling points within each maize farm were restricted to symptomatic plants, and the upper young leaf from maize plants showing MLN disease symptoms was collected. The collected leaf samples were stored in Ziploc paper storage bags in a cool box, labeled with the maize variety, where the isolate was obtained from, and the county. They were shipped to Botany and Plant pathology laboratory at Purdue University in USA for isolation of RNA and sequencing.
Extraction of RNA
From the 150 maize samples collected, 20 samples were randomly selected from each county for RNA isolation. The total RNA was extracted from individual maize leaves using the Trizol reagent (Invitrogen) as described by Chomzynski and Mackey (1995) followed by the RNeasy (QIAGEN) method as described by Adams et al. (2012)at Purdue University, Botany and Plant pathology laboratory, USA. The quantity of RNA and its purity was determined using the NanoDrop machine (2000c model) and stored at -80°C until the RT-PCR was done.
Reverse transcription (RT)-PCR amplification
RT-PCR amplification was carried out according to Jarugula et al. (2010)with modifications. It was a two-step RT-PCR reaction where cDNA was prepared from 1 μl of RNA using a Maxima First Strand cDNA Synthesis kit (Thermo Fisher Scientific, Wilmington, DE) as per the instruction manual. The cDNA primers (Table 1) were designed using the qPCR Primer Quest tool, Integrated DNA Technologies Inc.
The specific primers for amplification of the CP gene were designed based on the reported sequence of MCMV (JX286709) and SCMV (Adams et al., 2012)(Table 2). The RT-PCR was carried out using a thermal cycler (GeneAmp PCR systems 9700, Applied Biosystems) to amplify CP-specific DNA fragments. The cycling conditions were one cycle at 95°C for 5 min to allow denaturation, followed by 35 cycles at 94°C for 30s, 58°C for 30 s, 72°C for 60 s; and a final extension at 72°C for 7 min. An aliquot of the RT-PCR assay product was resolved on 1.5% agarose gels, stained with ethidium bromide, and the DNA bands were visualized under a UV-transilluminator.
Sequencing of the CP gene of MCMV and SCMV variants
Polymerase chain reaction products from the samples with the strongest bands when viewed under the U.V. light were purified for sequencing using the Qiagen kit following the manufacturer's instructions (QIAGEN Inc., Valencia, CA). The eluted DNA from MCMV and SCMV was used for sequencing, while WSMV was not sequenced since all the samples tested negative. Sequencing was done at Purdue Genomics Core Facility, USA, using the Sanger sequencing technique. The resulting sequences were then compared to published sequences in the GenBank-NCBI database (Benson et al., 2013)to give the identities of the sequences using blast+ (Camacho et al., 2009).
Sequence alignment and construction of phylogenetic trees
Multiple nucleotide sequence alignments were performed using Clustal Omega version 7. Manual editing was done to obtain consensus sequences. Aligned MCMV and SCMV sequences were assessed using DnaSP version 5.1 (83) software to estimate genetic diversity, and phylogenetic trees were constructed using the MEGA program version 7 (Kumar et al., 2016).
Maize lethal necrosis (MLN) symptoms observed in the field
As shown in Figure 1, the MLN symptoms illustrate the extent of the damage the disease poses on maize plants, especially among the susceptible varieties viz a viz the clean, uninfected maize plant. Maize plants in picture A show excessive chlorotic mottling with short internodes, and necrosis on leaf margin that progresses towards the midrib; while in picture B, the maize plants appear dwarf with severe chlorotic mottling on the upper leaves. The lower leaves have dried up due to excessive necrosis resulting in premature aging of the plants. As for picture C, the maize plants show no MLN symptoms.
RT-PCR detection of viruses causing MLN disease
The DNA from the 60 samples was amplified in the RT-PCR reaction. Out of these, 20 samples were positive to MCMV, while 18 samples were positive to SCMV. As for WSMV, all the 60 samples amplified were negative (Table 3).
Coat protein (CP) gene sequence of MCMV
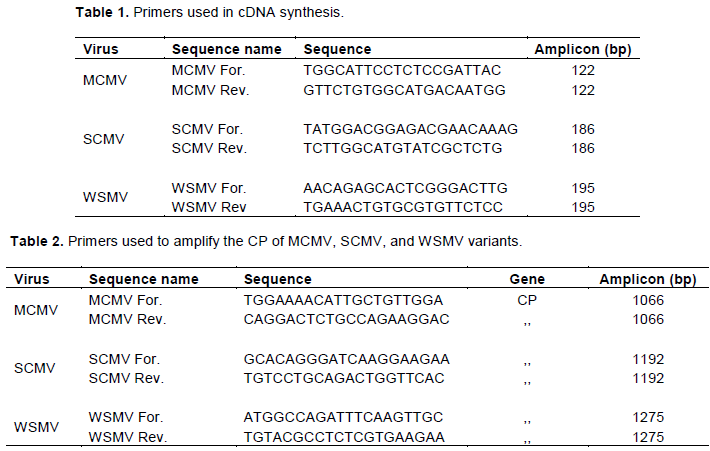
The CP sequence of MCMV isolates KCO 24 (accession # MH645622) and KSM 5 (accession #MH645621) were 100% identical to each other (Table 4). Also, the two isolates had 100% identity with Sub-Saharan isolate (KP851970), Ethiopia isolate (KP772217), and 100% identical to Rwanda isolate (KF744393). The sequences also had 99.86% identity to one isolate previously reported from Kenya (JX286709), 99% identity with MCMV isolates from Thailand (AM490791), and China (GU594293). Similarly, the sequences of the MCMV from the two Counties had 96.62% identity with sequences of MCMV isolates from Kansas (KS1) and Nebraska (EU358605) in the United States (Figure 2).
Coat protein (CP) gene sequence of SCMV
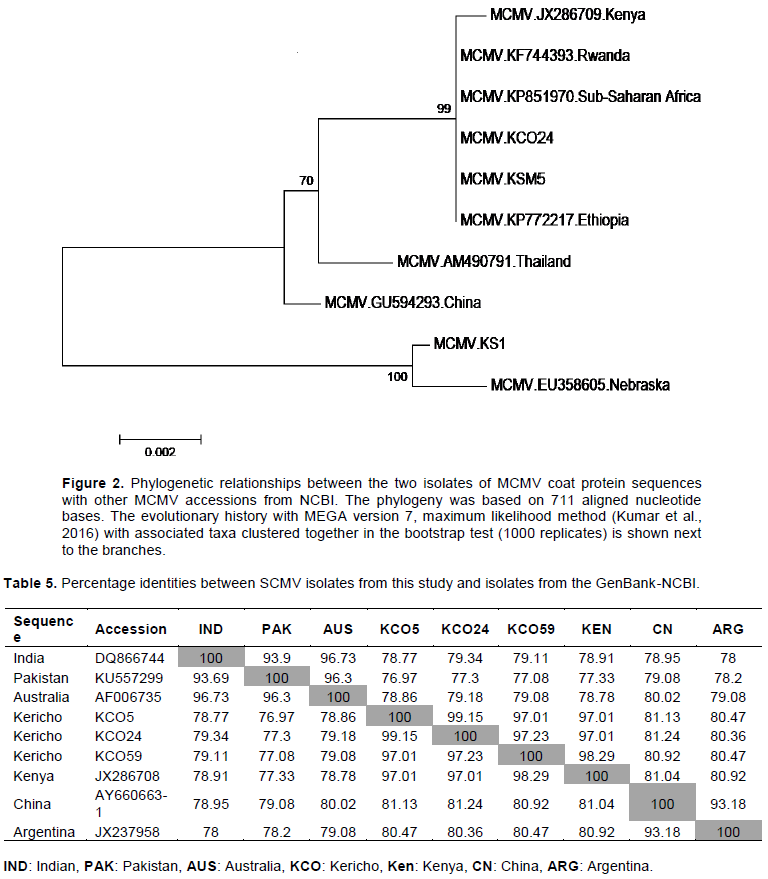
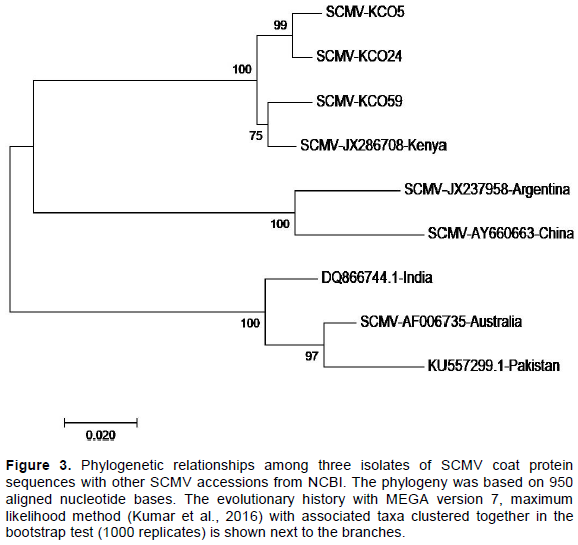
The KCO 5 (accession #MH645623) and KCO 24 SCMV (accession #MH645624) isolates had 99% identity with each other (Table 5), and 97% identity to KCO 59 (accession #MH645625) isolate and Kenya isolate (JX28708). Similarly, the SCMV isolate KCO 59 had 98.29% identity with the Kenya isolate (JX28708). The three SCMV isolates KCO5, KCO24, and KCO 59 had a similarity ranging from 76.97 to 81.24% with the other SCMV isolates from India, Pakistan, Australia, China, and Argentina (Figure 3).
The objective of the study is to characterize the viruses causing MLN in the South-Rift region of Kenya. From the results, only two viruses except for WSMV were detected in the samples from all the three counties studied. As reviewed in Mbega et al. (2016), MLN disease is caused by synergistic co-infection of maize MCMV with any cereal virus in the potyvirus MDMV, WSMV, or SCMV.
This study's findings have demonstrated that SCMV is the main potyvirus that causes the MLN disease in the South-Rift region. These findings concur with Adams et al. (2014), who also reported that SCMV is the most frequent potyvirus in Kenya and East Africa. Comparison of CP sequences of MCMV isolates (accession # MH645622 & #MH645621) from this study with other CP sequences of MCMV available in the GenBank-NCBI shows that they are closely related to Sub-Saharan Africa isolate (KP851970), Ethiopia isolates (KP772217), Rwanda isolates (KF744393), and Kenya isolate (JX286709). This high degree of MCMV sequence similarity with those from Eastern and Sub-Saharan Africa suggests a common point of origin for the Eastern and Sub-Saharan African MCMV isolates. This is a piece of evidence that coat protein is highly conserved and an indication that MCMV virus has not undergone a lot of variation. The low variation could be attributed to the natural selection pressure which has been reported to influence the genetic variation of a virus (Roossinck and Schneider, 2006). The virus is still adapting to the new conditions and environment as a result of its newness in the area as it was first reported in September 2011 in Kenya (Wangai et al., 2012). The sequences were also similar with China isolate (GU594293), and Thailand isolates (AM490791). These findings corroborate with Adams et al. (2012)and Mahuku et al. (2015). They also reported a high degree of similarity of CP sequences of MCMV within Eastern Africa and some Asian MCMV isolates.
In the phylogenetic tree containing the SCMV sequences (Figure 3), two isolates KCO5 (accession #MH645623) and KCO24 (accession #MH645624) shared a high degree of identity. In contrast, one isolate, KCO59 (accession #MH645625), had high similarity with the Kenyan isolate (JX286708), which had been previously reported. These results indicate the presence of two groups of SCMV sequences in the region. It suggests they may be new variants of SCMV in the South-Rift region. These findings concur with Gao et al. (2011)and Perera et al. (2009), who found out that new strains of SCMV continue to be reported in many countries. Therefore, these results reveal that SCMV is more diverse than MCMV in the region. The diverse nature of SCMV could be attributed to the fact that SCMV had been reported in Kenya as early as the 1970s (Louie, 1980)and in Eastern Africa in the mid-1930s (Kulkarni, 1972)and therefore it may have undergone some variations with time. Also, the heavy maize losses as high as 50% reported by De Groote et al. (2016)in areas between Kisii and Kericho could be attributed to the new variants of SCMV in the region. Besides, the diverse nature of SCMV in the region could also be attributed to the extensive- scale cultivation of SCMV susceptible crops in Poaceae family (maize, sorghum, sugarcane, napier grass, pasture grasses). This has have been reported by Wu et al. (2012) as facilitating build-up of SCMV in fields and dissemination of the virus over large areas in many countries worldwide.
This research has shown that SCMV is the main potyvirus in synergism with MCMV in causing MLN disease in Kenya. It has also demonstrated that MCMV isolates (accessions #MH645621 and #MH645621) from this study share a common origin with the MCMV isolates from Eastern and Sub-Saharan Africa. Further, the research has revealed that SCMV is more diverse than MCMV in the South-Rift region due to the existence of new variants of SCMV.
The authors have not declared any conflict of interests.
The authors sincerely appreciate the USDA-Norman E. Borlaug Fellowship for funding this study, Botany and Plant Department of Purdue University, for hosting him during the fellowship, Lonni Kucik, for the logistics support, Longfei Wang for the advice and graduate students in Johal’s Lab, Tesfaye and Woloshuk Lab.
REFERENCES
|
Adams IP, Harju VA, Hodges T, Hany U, Skelton A, Rai S, Boonham N (2014). First report of maize lethal necrosis disease in Rwanda. New Disease Reports 29(22).
Crossref
|
|
|
|
Adams IP, Miano DW, Kinyua ZM, Wangai A, Kimani E, Phiri N, Mumford R (2012). Use of next-generation sequencing for the identification and characterization of Maize chlorotic mottle virus and Sugarcane mosaic virus causing maize lethal necrosis in Kenya. Plant Pathology 10:1-9.
|
|
|
|
|
Benson DA, Cavanaugh M, Clark K, Karsch-mizrachi I, Lipman DJ, Ostell J, Sayers EW (2013). GenBank. Nucleic Acids Research 41(November 2012):36-42.
Crossref
|
|
|
|
|
Camacho C, Coulouris G, Avagyan V, Ma N, Papadopoulos J, Bealer K, Madden TL (2009). BLAST + :architecture and applications. BMC Bioinformatics 9:1-9.
Crossref
|
|
|
|
|
Chomzynski P, Mackey K (1995). Substitution of chloroform by bromo-chloropropane in the single-step method of RNA isolation. Analytical Biochemistry 225(1):163-164.
Crossref
|
|
|
|
|
Davis ERM (2012). First Report of Maize chlorotic mottle virus and Maize Lethal Necrosis in Kenya. Plant Disease 96(10):11-12.
Crossref
|
|
|
|
|
Gao B, Cui XW, Li XD, Zhang CQ, Miao HQ (2011). Complete genomic sequence analysis of a highly virulent isolate revealed a novel strain of Sugarcane mosaic virus. Virus Genes 43(3):390-397.
Crossref
|
|
|
|
|
De Groote H, Oloo F, Tongruksawattana S, Das B (2016). Community-survey based assessment of the geographic distribution and impact of maize lethal necrosis (MLN) disease in Kenya. Crop Protection 82:30-35.
Crossref
|
|
|
|
|
Isabirye BE, Rwomushana I (2016). Current and future potential distribution of maize chlorotic mottle virus and risk of maize lethal necrosis disease in Africa. Crop Breeding and Applied Biotechnology 5(2):215-228.
Crossref
|
|
|
|
|
Jarugula S, Alabi OJ, Martin RR, Naidu RA (2010). Genetic Variability of Natural Populations of Grapevine leafroll-associated virus 2 in Pacific Northwest Vineyards. Virology (21).
Crossref
|
|
|
|
|
Kagoda F, Gidoi, R, Isabirye BE (2016). Status of maize lethal necrosis in eastern Uganda. African Journal of Agricultural Research 11(8):652-660.
Crossref
|
|
|
|
|
Kamau CN, Snipes K (2014). Kenya Grain and Feed Annual.1-5.
|
|
|
|
|
Kiruwa FH, Feyissa T, Ndakidemi PA (2016). Insights of maize lethal necrotic disease : A major constraint to maize production in East Africa 10(9):271-279.
Crossref
|
|
|
|
|
Kulkarni HY (1972). Survey of viruses affecting east african major food crops. East African Agricultural and Forestry Journal 39(2):158-164.
Crossref
|
|
|
|
|
Kumar S, Stecher G, Tamura K (2016). MEGA7 : Molecular Evolutionary Genetics Analysis Version 7 . 0 for Bigger Datasets Brief communication. Molecular Biology and Evolution 33(7):1870-1874.
Crossref
|
|
|
|
|
Louie R (1980). Sugarcane Mosaic Virus in Kenya. Plant Disease 64(10):944-947.
Crossref
|
|
|
|
|
Lukanda M, Owati A, Ogunsanya P, Valimunzigha K, Katsongo K, Ndemere H, Kumar PL (2014). First Report of Maize chlorotic mottle virus Infecting Maize in the Democratic Republic of the Congo. Plant Disease 98(10):1448-1448.
Crossref
|
|
|
|
|
Mahuku G, Lockhart BE, Wanjala B, Jones MW, Kimunye JN, Stewart LR, Cassone BJ, Sevgan S, Nyasani JO, Kusia E, Lava Kumar P, Niblett CL, Kiggundu A, Asea G, Pappu HR, Wangai A, Prasanna BM, Redinbaugh MG (2015). Maize Lethal Necrosis (MLN), an Emerging Threat to Maize-Based Food Security in Sub-Saharan Africa. Ecology and Epidemiology 105(7):956-963.
Crossref
|
|
|
|
|
Makone SM, Menge D, Basweti E (2014). Impact of Maize Lethal Necrosis Disease on maize yield:A case of Kisii, Kenya. International Journal of Agricultural Extension 02(03):211-218.
|
|
|
|
|
Mbega ER, Ndakidemi PA, Mamiro DP, Mushongi AA, Kitenge KM, Ndomba OA (2016). Role of Potyviruses in Synergistic Interaction Leading to Maize Lethal Necrotic Disease on Maize. International Journal of Current Microbiology and Applied Sciences 5(6):85-96.
Crossref
|
|
|
|
|
Mezzalama M (2015). MLN Pathogen Diagnosis, MLN-free Seed Production and Safe Exchange to Non-Endemic Countries. CIMMYT Institutional Multimedia Publications Repository.
|
|
|
|
|
Perera MF, Filippone MP, Ramallo CJ, Cuenya MI, García ML, Ploper LD, Castagnaro AP (2009). Genetic diversity among viruses associated with sugarcane mosaic disease in Tucumán, Argentina. Phytopathology 99(1):38-49.
Crossref
|
|
|
|
|
Roossinck MJ, Schneider WL (2006). Genetic Diversity in RNA Virus Quasispecies Is Controlled by Host-Virus Interactions Genetic Diversity in RNA Virus Quasispecies Is Controlled by Host-Virus Interactions. Journal of Virology 299(14):337-348.
|
|
|
|
|
Wangai AW, Redinbaugh MG, Kinyua ZM, Miano DW, Leley PK, Kasina M, Mahuku G, Scheets K, Jeffers D (2012). Zea mays. Plant Disease 96(10):1582.
Crossref
|
|
|
|
|
Wu L, Zu X, Wang S, Chen Y (2012). Sugarcane mosaic virus e Long history but still a threat to industry. Crop Protection 42:74-78.
Crossref
|
|