Full Length Research Paper

ABSTRACT
Salinity is a major detrimental abiotic factor for plant growth. The main purpose of this study was to analyze the effects of different NaCl concentrations on growth and some physiological parameters related to gas exchanges and water relations in amaranth (Amaranthus cruentus) plants. Three weeks old amaranth plants from the cultivar ‘Locale’ were exposed in nutrient solution to 0, 30 or 90 mM NaCl (electrical conductivities of 1.915; 4.815 and 11.70 dS.m-1 respectively) in phytotron conditions. Shoot elongation as well as fresh and dry masses of shoot and root were determined after two weeks of stress exposure. Net photosynthesis (A), intercellular CO2 concentration (Ci), instantaneous transpiration (E), stomatal conductance (gs), osmotic potential (Ψs) as well as the efficiency of the instantaneous carboxylation (A/Ci), intrinsic (A/gs) and instantaneous (A/E) water use efficiency were estimated. Results reveal that salt stress induced a significant reduction in growth of aerial part as well as net photosynthesis, instantaneous transpiration, stomatal conductance and leaf and root osmotic potentials. In contrast, no significant reductions were recorded for root growth, shoot water content, intercellular CO2 concentration and instantaneous carboxylation efficiency. However, a significant increase was observed for intrinsic (A/gs) and instantaneous (A/E) water use efficiency. The plant growth reduction observed hinges upon a drop in photosynthetic activity due mainly to stomatal closure. These data suggest that photosynthetic activity may be used as a reliable criterion for physiological estimation of salt-tolerance in A. cruentus cultivars.
Key words: Saline stress, net photosynthesis, stomatal conductance, osmotic potential, water use efficiency.
INTRODUCTION
MATERIALS AND METHODS

RESULTS AND DISCUSSION
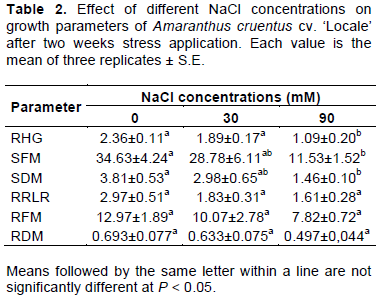
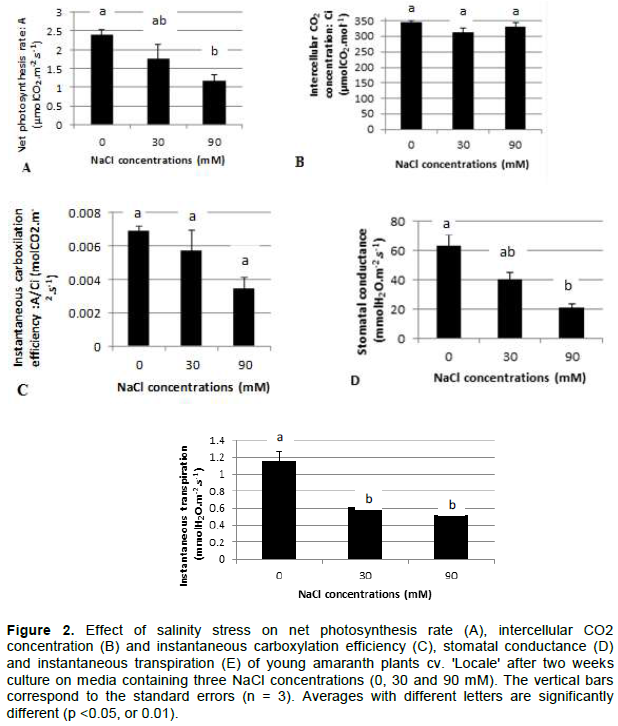
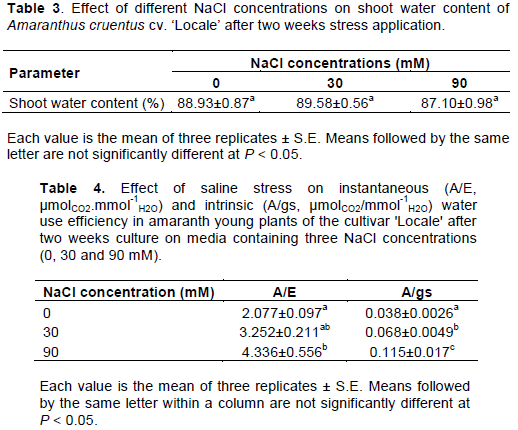

CONCLUSION
The reduction of plant growth under salt stress is due to a loss in photosynthetic activity mostly related to stomatal closure. The maintenance of high water use efficiency appeared as an important strategy to face salt stress in A. cruentus. The results exposed here provide specific physiological cues for improvement of salt-tolerance in amaranth.
ACKNOWLEDGEMENTS
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Abbas W, Ashraf M, Akram NA (2010). Alleviation of salt-induced adverse effects in eggplant (Solanum melongena L.) by glycine betaine and sugarbeet extracts. Scientia Horticulturae 125: 188-195. |
|
|
Acosta-Motos JR, Ortu-o MF, Bernal-Vicente A, Diaz-Vivancos P, Sanchez-Blanco MJ, Hernandez JA (2017). Plant responses to salt stress: Adaptive Mechanisms. Agronomy pp. 1-38. |
|
|
Akhtar S, Wahid A, Rasul E (2003). Emergence, growth and nutrient composition of sugarcane sprouts under NaCl salinity. Biologia Plantarum 46(1):113-116. |
|
|
Akram NA, Ashraf M, Al-Qurainy F (2012). Aminolevulinic acid-induced changes in some key physiological attributes and activities of antioxidant enzymes in sunflower (Helianthus annuus L.) plants under saline regimes. Scientia Horticulturae 142:143-151. |
|
|
Allemann J, Van Den Heever E, Viljoen J (1996). Evaluation of Amaranthus as a possible vegetable crop. Applied Plant Science 10:1-4. |
|
|
Almansouri M, Kinet JM, Lutts S (1999). Compared effects of sudden and progressive impositions of salt stress in three Durum wheat (Triticum durum Desf.) cultivars. Journal of Plant Physiology 154:743-752. |
|
|
Álvarez S, Gómez-Bellot MJ, Castillo M, Ba-ón S, Sánchez-Blanco MJ (2012). Osmotic and saline effect on growth, water relations, and ion uptake and translocation in Phlomis purpurea plants. Environmental and Experimental Botany 78:138-145. |
|
|
Álvarez S, Sánchez-Blanco MJ (2014). Long-term effect of salinity on plant quality, water relations, photosynthetic parameters and ion distribution in Callistemon citrinus. Plant Biology 16:757-764. |
|
|
Amukali O, Obadoni BO, Mensah JK (2015). Effects of different NaCl concentrations on germination and seedlings growth of Amaranthus hybridus and Celosia argentea. African Journal of Environmental Science and Technology 9:301-306. |
|
|
Apel K, Hirt H (2004). Reactive oxygen species: metabolism, oxidative stress and signal transduction. Annual Review of Plant Biology 55:373-99. |
|
|
Ashraf M (2001). Relationships between growth and gas exchange characteristics in some salt-tolerant amphidipoid Brassica species in relation to their diploid parents. Environmental and Experimental Botany 45:155-163. |
|
|
Ashraf M, Akram NA, Al-Qurainy F, Foolad MR (2011). Drought tolerance: roles of organic osmolytes, growth regulators and mineral nutrients. In Advances in Agronomy Academic Press 111:249-296. |
|
|
Ashraf M, Harris JC (2004). Potential biochemical indicators of salinity tolerance in plants. Plant Science 166:3-16. |
|
|
Ashraf M, Waheed A (1993). Responses of some local/exotic accessions of lentil (Lens culinaris Medic.) to salt stress. Journal of Agronomy and Crop Science 170:103-112. |
|
|
Ashraf MY, Bhatti AS (2000). Effect of salinity on growth and chlorophyll content in rice. Pakistan Journal of Scientific and Industrial Research 43:130-131. |
|
|
Baker NR, Rosenqvist E (2004). Applications of chlorophyll fluorescence can improve crop production strategies: an examination of future possibilities. Journal of Experimental Botany 55:1607-1621. |
|
|
Bayuelo-Jiménes JS, Debouck DG, Lynch JP (2002). Salinity tolerance of Phaseolus species during early vegetative growth. Crop Science 42:2184-2192. |
|
|
Bhattacharjee S (2008). Triadimefon pretreatment protects newly assembled membrane system and causes up-regulation of stress proteins in salinity stressed Amaranthus lividus L. during early germination, Journal of Environmental Biology 29(5):805-810. |
|
|
Brugnoli E, Lauteri M (1991). Effects of salinity on stomatal conductance, photosynthetic capacity, and carbon isotope discrimination of salt-tolerant (Gossypium hirsutum L.) and salt-sensitive (Phaseolus vulgaris L.) C3 non-halophytes. Plant Physiology 95:628-635. |
|
|
Chaves MM, Flexas J, Pinheiro C (2009). Photosynthesis under drought and salt stress: regulation mechanisms from whole plant to cell. Annnals of Botany 103: 551-560. |
|
|
Chen XL, Zhao KF (1996). Effect of NaCl stress on germination of maize and the alleviation of exogenous Ca2+. Acta Agriculturae Boreali-Sinica 11:89-92. |
|
|
Cramer GR, Alberico GJ, Schidt C (1994). Salt tolerance is not associated with the sodium accumulation of two maize hydrids. Australian Journal of Plant Physiology 21:675-692. |
|
|
Cramer GR, Bowman DC (1991). Kinetics of maize leaf elongation. I. Increased yield threshold limits short-term, steady-state elongation rates after exposure to salinity. Journal of Experimental Botany 42:1417-1426. |
|
|
Cunningham AB, Dejager PJ, Hansen LCB (1992). The indigenous plant use programme. Foundation for Research Development, Pretoria. |
|
|
da Silva EN, Ribeiro RV, Ferreira-Silva SL, Viégas RA, Silveira JAG (2011). Salt stress induced damages on the photosynthesis of physic nut young plants. Scientia Agricola 68:62-68. |
|
|
Davies WJ, Kudoyarova G, Hartung W (2005). Long-distance ABA signaling and its relation to other signaling pathways in the detection of soil drying and the mediation of the plant's response to drought. Journal of plant growth regulation 24:285-295. |
|
|
Delfine S, Alvino A, Zacchini M, Loreto F (1998). Consequences of salt stress on conductance to CO2 diffusion, Rubisco characteristics and anatomy of spinach leaves. Australian Journal of Plant Physiology 25:395-402. |
|
|
Fisarakis I, Chartzoulakis K, Stavrakas D (2001). Response of sultana vines (V. vinifera L.) on six rootstocks to NaCl salinity exposure and recovery. Agricultural Water Management 51:13-27. |
|
|
Foyer CH, Noctor G (2005). Oxidant and antioxidant signalling in plants: a re-evaluation of the concept of oxidative stress in a physiological context. Plant, Cell and Environment 28:1056-1071. |
|
|
Franco JA, Ba-ón S, Vicente MJ, Miralles J, Martínez-Sánchez JJ (2011). Root development in horticultural plants grown under abiotic stress conditions- A review. Journal of Horticultural Science and Biotechnology 86:543-556. |
|
|
Fricke W, Akhiyarova G, Veselov D, Kudoyarova G (2004). Rapid and tissue-specific changes in ABA and in growth rate response to salinity in barley leaves. Journal of Experimental Botany 55:1115-1123. |
|
|
Gandonou CB, Bada F, Gnancadja SL, Abrini J, Skali Senhaji N (2011). Effects of NaCl on Na+, Cl- and K+ ions accumulation in two sugarcane (Saccharum sp.) cultivars differing in their salt tolerance, International Journal of Plant Physiology and Biochemistry 3(10):155-162. |
|
|
Geissler N, Hussin S, Koyro HW (2009). Elevated atmospheric CO2 concentration ameliorates effects of NaCl salinity on photosynthesis and leaf structure of Aster tripolium L. Journal of Experimental Botany 60:137-151. |
|
|
Hussain K, Majeed A, Nawaz K, Khizar HB, Nisar MF (2009). Effect of different levels of salinity on growth and ion contents of black seeds (Nigella sativa L.). Current Research Journal of Biological Sciences 1:135-138. |
|
|
Jaleel CA, Manivannan P, Sankar B (2007). Induction of drought stress tolerance by ketoconazole in Catharanthus roseus is mediated by enhanced antioxidant potentials and secondary metabolite accumulation. Colloids and surfaces B: Biointerfaces 60:201-206. |
|
|
James RA, Rivelli AR, Munns, R, von Caemmerer S (2002). Factors affecting CO2 assimilation, leaf injury and growth in salt-stressed durum wheat. Functional Plant Biology 29:1393-1403. |
|
|
Kerstiens G, Tych W, Robinson MF, Mansfield TA (2002). Sodium-related partial stomatal closure and salt tolerance of Aster tripolium. New Phytologist 153:509-515. |
|
|
Koyro HW (2006). Effect of salinity of growth, photosynthesis, water relations and solute composition of the potential cash crop halophyte Plantago coronopus (L.). Environmental and Experimental Botany 56:136-146. |
|
|
Lavini A, Pulvento C, d'Andria R, Riccardi M (2016). Effects of saline irrigation on yield and qualitative characterization of seed of an amaranth accession grown under Mediterranean conditions. The Journal of Agricultural Science 154(5):858-869. |
|
|
Lee GJ, Carrow RN, Duncan RR (2004). Photosynthetic responses of salinity stress of halophytic seashore Paspalum ecotypes. Plant Science 166:1417-1425. |
|
|
Leidi EO, Saiz JF (1997). Is salinity tolerance related to Na accumulation in Upland cotton (Gossypium hirsutum) seedlings? Plant and Soil 190:67-75. |
|
|
Liu Y, Du H, Wang K, Huang B, Wang Z (2011). Differential photosynthetic responses to salinity stress between two perennial grass species contrasting in salinity tolerance. HortScience 46(2):311-316. |
|
|
Logan BA (2005). Reactive oxygen species and photosynthesis. In Antioxidants and Reactive Oxygen Species in Plants, ed. N Smirnoff. Oxford: Blackwell pp. 250-267. |
|
|
Lutts S, Bouharmont J, Kinet J.M (1999). Physiological characterization of salt-resistant rice (Oryza sativa) somaclones. Australian Journal of Botany 47:835-849. |
|
|
Lutts S, Kinet JM, Bouharmont J (1996). Effects of salt stress on growth, mineral nutrition and proline accumulation in relation to osmotic adjustment in rice (Oryza sativa L.) cultivars differing in salinity resistance. Plant Growth Regulation 19:207-218. |
|
|
Lycoskoufis IH, Savvas D, Mavrogianopoulos G (2005). Growth, gas exchange, and nutrient status in pepper (Capsicum annuum L.) grown in recirculating nutrient solution as affected by salinity imposed to half of the root system. Scientia Horticulturae 106:147-161. |
|
|
Makus DJ (2003). Salinity and Nitrogen Level Affect Agronomic Performance, Leaf Color and Leaf Mineral Nutrients of Vegetable Amaranth. Subtropical Plant Science 55:1-6. |
|
|
Megdiche W, Hessini K, Gharbi F, Jaleel CA, Ksouri R, Abdelly C (2008). Photosynthesis and photosystem-efficiency of two salt adapted halophytic seashore Cakile maritima ecotypes. Photosynthetica 46:410-419. |
|
|
Munns R (2002). Comparative physiology of salt and water stress. Plant, Cell and Environment 25: 239-250. |
|
|
Munns R (2005). Genes and salt tolerance: Bringing them together. Plant phytology 167:645-663. |
|
|
Munns R, Guo J, Passioura JB, Cramer GR (2000). Leaf water status controls day-time but not daily rates of leaf expansion in salt-treated barley. Australian Journal of Plant Physiology 27:949-957. |
|
|
Munns R, James RA, Laüchli A (2006). Approaches to increasing the salt tolerance of wheat and other cereals. Journal of Experimental Botany 57:1025-1043. |
|
|
Munns R, Tester M (2008). Mechanisms of salinity tolerance. Annual Review of Plant Biology 59:651-681. |
|
|
Niu G, Rodriguez DS, Starman T (2010). Response of bedding plants to saline water irrigation. HortScience 45 (4): 628-636. |
|
|
Noreen Z, Ashraf M, Akram NA (2010a). Salt-induced modulation in some key physio-biochemical processes and their use as selection criteria in potential vegetable crop pea (Pisum sativum L.). Crop Pasture Science 61:369-378. |
|
|
Noreen Z, Ashraf M, Akram NA (2010b). Salt-induced regulation of some key antioxidant enzymes and physio-biochemical phenomena in five diverse cultivars of turnip (Brassica rapa L.). Journal of Agronomy and Crop Science 196:273-285. |
|
|
Noreen Z, Ashraf M, Akram NA (2012). Salt-induced regulation of photosynthetic capacity and ion accumulation in some genetically diverse cultivars of radish (Raphanus sativus L.). Journal of Applied Botany and Food Quality 85:91-96. |
|
|
Ornami EN, Hammes PS (2006). Ameliorative effects of calcium on growth and mineral uptake of salt-stressed amaranth, South African Journal of Plant and Soil 23(3):197-202. |
|
|
Parida A, Das AB, Das P (2002). NaCl stress causes changes in photosynthetic pigments, proteins, and other metabolic components in the leaves of a true mangrove, Bruguiera parviflora, in hydroponic cultures. Journal of Plant Biology 45:28-36. |
|
|
Passioura JB, Munns R (2000). Rapid environmental changes that affect leaf water status induce transient surges or pauses in leaf expansion rate. Australian Journal of Plant Physiology 27:941-948. |
|
|
Passioura JB (2010). Scaling up: The essence of effective agricultural research. Functional Plant Biology 37: 585-591. |
|
|
Prakash OM, Zaidi PH (2000). Effect of amaranth (Amaranthus spinosus L.) supplemented maize diet on blood haemoglobin and lipid metabolism in rats. Annals of Agricultural Research 21(2):223-232. |
|
|
Qin L, Guo S, Ai W, Tang Y, Cheng Q, Chen G (2013). Effect of salt stress on growth and physiology in amaranth and lettuce: Implications for bioregenerative life support system. Advances in Space Research 51:476-482. |
|
|
R'him T, Tlili I, Hnan I, Ilahy R, Benali A, R'him JH (2013). Effet du stress salin sur le comportement physiologique et métabolique de trois variétés de piment (Capsicum annuum L.). Journal of Applied Biosciences 66:5060-5069. |
|
|
Sánchez-Blanco MJ, Rodríguez P, Morales MA, Ortu-o MF, Torrecillas A (2002). Comparative growth and water relations of Cistus albidus and Cistus monspeliensis plants during water deficit conditions and recovery. Plant Science 162:107-113. |
|
|
Sánchez-Blanco MJ, Rodríguez P, Olmos E, Morales MA, Torrecillas A (2004). Differences in the effects of simulated sea aerosol on water relations, salt content, and leaf ultrastructure of rock-rose plants. Journal of Environmental Quality 33:1369-1375. |
|
|
SAS Institute (2015). SAS/STAT user's guide, Vol. 1; Release 6.03, ed. SAS Institute Inc. Cary, NC. USA. |
|
|
Shaheen S, Naseer S, Ashraf M, Akram AN (2013). Salt stress affects water relations, photosynthesis, and oxidative defense mechanisms in Solanum melongena L. Journal of Plant Interactions 8(1):85-96. |
|
|
Shahid MA, Pervez MA, Balal RM, Ahmad R, Ayyub CM, Abbas T, Akhtar N (2011). Salt stress effects on some morphological and physiological characteristics of okra (Abelmoschus esculentus L.). Soil and Environment 30(1):66-73. |
|
|
Shannon MC, Grieve CM (1999). Tolerance of vegetable crops to salinity. Scientia horticulturae 78:5-38. |
|
|
Sobrado MA (2005). Leaf characteristics and gas exchange of the mangrove Laguncularia racemosa as affected by salinity. Photosynthetica 43:217-221. |
|
|
Stoeva N, Kaymakanova M (2008). Effect of salt stress on the growth and photosynthesis rate of bean plants. Journal of Central European Agriculture 9:385-392. |
|
|
Villa-Castorena M, Ulery AL, Catalan-Valencia EA, Remmenga MD (2003). Salinity and nitrogen rate effects on the growth and yield of Chile pepper plants. Soil Science Society of America Journal 67:1781-1789. |
|
|
Vysotskaya L, Hedley PE, Sharipova G, Veselov D, Kudoyarova G, Morris J, Jones HG (2010). Effect of salinity on water relations of wild barley plants differing in salt tolerance. AoB Plants 86:407-421. |
|
|
Wahid A (2004). Analysis of toxic and osmotic effects of sodium chloride on leaf growth and economic yield of sugarcane. Botanical Bulletin of Academia Sinica 45:133-141. |
|
|
Went FE (1957). The experimental control of plant growth, Chronica Botanica Co. Ronald Press Co; Waltham M, n° 17: 343 p. New York. |
|
|
Wouyou DA (2017). Réponse de l'amarante (Amaranthus cruentus, l.) au stress salin: caractérisation de cultivars, mécanisme physiologique de résistance et qualité nutritionnelle des feuilles, thèse de doctorat, Université d'Abomey-Calavi, Décembre 2017, 155 p. |
|
|
Wouyou A, Gandonou CB, Montcho D, Kpinkoun J, Kinsou E, Assogba Komlan F, Gnancadja LS (2016). Salinity resistance of six Amaranth (Amaranthus sp.) cultivars cultivated in Benin at germination stage. International Journal of Plant and Soil Science 11(3):1-10. |
|
|
Wouyou A, Gandonou CB, Assogba-Komlan F, Montcho D, Zanklan AS, Lutts S, Gnancadja SL (2017). Salinity resistance of five amaranth (Amaranthus cruentus) cultivars at young plant stage. International Journal of Plant and Soil Science 14:1-11. |
|
|
Zhang S, Li Q, Ma K, Chen L (2001). Temperature-dependent gas exchange and stomatal/nonstomatal limitation to CO2 assimilation of Quercus liaotungensis under midday higher irradiance. Photosynthetica 39:383-388. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


