Full Length Research Paper

ABSTRACT
Co-digestion of water hyacinth (WH) (E. crassipes) with ruminal slaughterhouse waste (RSW) has the potential for improving biogas production by complementing process parameters. This study evaluated microbial communities in co-digestion of WH with RSW at 32ËšC by isolation, phenotype analysis, DNA extraction and PCR amplification of the 16S rRNA genes. Digestion of WH and RSW separately exhibited lag times of 5 to 20 days that were associated with pH drop to acidic zone. The pH drop was attributed to rapid production of volatile fatty acids by acidogenesis process without corresponding consumption by methanogenesis. Co-digestion at 30% RSW maintained alkaline pH and increased biogas yield for WH from 47 to 95% of the 42.1 L CH4/kg observed for RSW suggesting synergy in the co-digestion. Morphologies of colonies isolated from the reactors were dominated by short and long rods bacilli with some cocci, and streptococcus mainly in WH samples. About 77% of the isolates were Gram positive, suggesting dominance of Firmicutes phyla that includes Bacillus genus. Molecular analysis observed a shift in microbial community during the acidic lag phase from Bacillus genus to acetogenic bacteria Lysinibacillus and Solibacillus genera that consume volatile acids increasing pH. Recovery of alkaline conditions resulted in emergence of diverse species of Bacillus sp. associated with fermentation and syntrophic processes that included Bacillus aerophilus, Bacillus pumilus, Bacillus cereus, Bacillus thuringiensis, Bacillus licheniformis, Bacillus glycinifermentans, and Bacillus aquimaris. Co-digestion of WH with RSW collated processes parameters and constrained drop in pH to alkaline levels conducive for growth of Bacillus sp.
Key words: Microbial communities, methanogenesis, 16S rRNA genes, polymerase chain reaction (PCR), methanogens.
INTRODUCTION
Water hyacinth (Eichhonia crassipes), a freshwater plant, is of environmental concern in many water bodies such as the East African Lake Victoria because of its capacity to grow fast and multiply and form floating mats (Gichuki et al., 2012; Tao et al., 2016). However, these characteristics make the plant available inexpensively and in large quantities with potential for use as biomass in biogas production. Moreover, co-digestion of the water hyacinth (WH) with other substrates could assist to overcome some of the limitations of single substrate digestion by complementing process parameters (Mata-Álvare et al., 2014). Previously, Omondi et al. (2019) found ruminal slaughterhouse waste (RSW), a potential co-substrate for WH, comprised greater proportions of carbohydrates and crude protein than the WH, about the same proportion of crude fiber (cellulose and lignin), and less of ash, which would make it more amenable to anaerobic digestion. However, some process parameters such as C/N and C/P ratios for both substrates required optimization. Co-digestion of water hyacinth (WH) with ruminal RSW reduced inhibitory drop in pH for RSW and increased biogas yield for WH. Because microbial communities are key players in the digestion process, knowledge of their dynamics would be useful in understanding the synergies in co-digestion.
Anaerobic digestion (AD) involves a cascade of steps mainly hydrolysis, acidogenesis, acetogenesis and methanogenesis in which a consortium of bacteria and archaea convert organic matter into biogas. During hydrolysis, extracellular enzymes secreted by fermentative bacteria degrade organic macromolecules such as proteins, carbohydrates and fats, into amino acids, sugars and long chain fatty acids, which are then absorbed into the cell (Gerber and Span, 2008; Meyer and Edwards, 2014). Acidogenic bacteria convert the products of hydrolysis to volatile fatty acids such as benzoic, butyrate and propionic acids and esters of p-hydroxybenzoic acid (parabens). Other products include alcohols and ketones, acetate, CO2 and H2. Acetogenic bacteria then convert volatile fatty acids and alcohols into acetate, H2 and CO2, which are then used by methanogenic archaea in syntrophic reactions to form methane (Ahring et al., 2003). Although the steps of the digestion are sequential, reactions within the digesters take place concurrently allowing continuous biogas production.
Hydrolysis is generally the rate limiting step in AD, which has been attributed to limited surface area for enzymatic action (Zeeman and Sanders, 2001). For example, in ligneous substrates such as WH the biodegradable polymer, cellulose, is shielded by both lignin and hemicellulose (Taherzadeh and Karimi, 2007). The succeeding acidogenesis step is exothermic and takes place rapidly producing volatile fatty acids. If the following acid consuming syntrophic reactions are slower and do not match the acid production, the reactor pH drops to acidic levels (Taconi et al., 2008). The acidic environment can become inhibitory to methanogens causing a lag in biogas production (Czatzkowska et al., 2020).
Microbial communities involved in the AD process are largely dependent on substrate composition and reactor design as well as operating conditions (McHugh et al., 2003). Sequence analyses of bacteria domains in anaerobic digestion of sludge by Rivière et al. (2009) revealed four dominant phyla; namely, Chloroflexi, Proteobacteria, Bacteroidetes, and Firmicutes and that the Archaea community that are responsible for methanogenesis were affiliated with methanosarcinales and methanomicrobiales groups. The proteobacteria are Gram-negative bacteria while Firmicutes are a low-G+C group, mostly Gram-positive consisting of Bacilli and Clostridia classes. Heeg et al. (2014) associated the higher efficiency of anaerobic digestion in the thermophilic environment to abundance of Firmicutes and Methanosarcina species. In anaerobic digestion of lignocellulosic residues of palm oil mill and wheat straw, bacterial community included Ruminococcus, Thiomargarita, Clostridium, Anaerobacter, Bacillus, and Sporobacterium species (Heeg et al., 2014; Suksong et al., 2016). This study aimed to establish the dynamics of the microbial community in anaerobic digestion of WH with RSW that could assist in understanding of synergies in co-digestion.
METHODOLOGY
This study investigated microbial communities in anaerobic digestion of WH and RSW, separately and in co-digestion. Water hyacinth samples were obtained from Winam Gulf in Lake Victoria, near Kisumu city in Kenya (coordinates 0° 53’ 9.71”S, 34° 45’2.44”E) while RSW was obtained from Dagoretti Slaughterhouse, located in the western part of Nairobi (coordinates 1°17'3.71"S, 36°41'1.98"E). The substrates were digested in laboratory batch reactors and the cumulative biogas production measured. Bacterial communities in three digestion stages; namely, acclimatization, lag and active methanation were isolated and identified using culture and biochemical phenotype analyses, and through DNA extraction and PCR amplification of the 16S rRNA genes.
Anaerobic digestion
Anaerobic digestion was conducted in three 1,000 mL round bottom flasks fitted with long probe digital thermometer DT501LAB and pH meter HI98103 from Hanna Instruments (Figure 1). Substrate samples were prepared by crushing and weighing into digesters. The digesters labeled D0, D30, and D100, were fed with 150 g of WH and RSW substrate mixture with respective RSW proportions of 0, 30 and 100% in 100 ml water. Anaerobic digestions were performed at mesophilic temperature of 32°C for a retention time of 60 days. The gas generated passed through an alkaline scrubber solution for removal of CO2 and other minor gases. The volume of resultant methane gas was measured by water displacement into a graduated measuring cylinder (Esposito et al., 2012). The temperature and pH of the reactor were monitored daily.
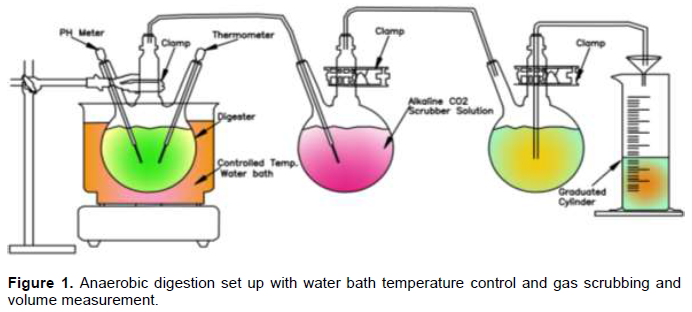
Sampling of sludge for analysis of microbial communities
Microbial communities in each of the digestion stages of acclimatization, lag and active biomethanation were determined from live sludge samples. An initial test run was carried out where retention times for initial alkaline, subsequent acidic and final alkaline pH values that indicate the specific biomethanation stages were determined for investigation of microbial activity. The pre-determined sampling days were Day 5, 20, and 35 of digester operation. Approximately 10 g of sludge was sampled in triplicate from each digester and kept in the laboratory at 4°C awaiting analysis.
Isolation and identification of microbial community
Isolation and identification of microbial communities took place at the Institute for Biotechnology Research, Jomo Kenyatta University of Agriculture and Technology (JKUAT). Microbial colonies were isolated by spread plate culture method and identified using Gram staining method. Approximately 1 g of the substrate collected from active digesters was transferred aseptically into a sterile 250 ml conical flask containing 90 ml of sterile distilled water. The mixture was agitated gently at 150 rpm for 30 min to homogenize the samples and release bacteria into solution. Nutrient agar media was prepared, autoclaved and poured into Petri dish plates for inoculation. Serial dilution procedure was carried out up to 10-4 times the original sample. The solutions were vortexed and 100 mL was transferred onto plates and spread out uniformly. The inoculated plates were sealed and incubated at 37°C for 14 h. Organisms of interest were identified using colony morphology before being inoculated into sterile nutrient agar plates for identification of bacteria colonies. The plates were incubated at 37°C for 12 h. Four distinct organisms were selected and streaked on to fresh plates. To avoid frequent culturing, pure bacterial colonies were preserved in glycerol until DNA extraction and identification of different bacterial groups.
DNA extraction
DNA was extracted from isolated bacteria colonies using phenol chloroform procedures (Thikra, 2013). An overnight broth culture was transferred into a sterile centrifuge tube and centrifuged at 6,000 rpm for 5 min. The supernatant was discarded and the pellet re-suspended in 200 mL of TE (Tris EDTA) buffer and cells washed by vortex. The homogenate was centrifuged for 5 min at 6,000 rpm. The supernatant was discarded and the pellet re-suspended in 200 mL of TE buffer. Approximately 20 mL of 10% sodium dodecyl sulphate (SDS), 10 mL of lysozyme (20 mg/mL) and 10 mL RNase were added and mixed gently before incubation at 37°C for 1 h. Approximately 10 mL of proteinase K were added and mixed gently and then incubated at 55°C for 1 h. DNA extraction was carried out by adding an equal volume of phenol/chloroform/isoamyl alcohol at a ratio 25:24:1 to the sample. The samples were mixed gently by inversion then centrifuged at 10,000 rpm for 10 min. The supernatant was transferred into a clean tube and an equal volume of chloroform/isoamyl (24:1) added, mixed gently and centrifuged. The DNA extraction with chloroform/isoamyl was repeated twice. The supernatant was transferred into sterile tubes and its volume determined. Absolute ethanol 2.5 mL was added to the supernatant and the contents mixed well and incubated overnight at 20°C. The samples were centrifuged at 10,000 rpm for 10 min and supernatant discarded. 100 µL of 70% ethanol was added to the pellet and incubated at room temperature for 15 min. The samples were again centrifuged at 13,000 rpm for 10 min and the supernatant discarded. The pellets were dried in an incubator at 37°C for 30 min and then suspended in 100 µL of ultra-pure water. The DNA solution was kept at 4°C. Electrophoresis of DNA was carried out on 1% agarose gel in Tris-Borate-EDTA (TBE) buffer and 0.1 µL of ethidium bromide added. The gel was visualized under UV.
PCR amplification of 16S rRNA gene
There are several methods for quantification of microbial communities (Raskin et al., 1994; Akarsubasi et al., 2005; Stainberg and Regan, 2009). The 16S rRNA and its genes that are that most frequently used biomarkers for the determination of methanogenic populations in environments (Takashi and Yuji, 2011) was selected for this study. Amplification of DNA was carried out at the Institute for Biotechnology Research, Jomo Kenyatta University of Agriculture and Technology. Bacterial 16S rRNA genes of the pure isolates were amplified using PeQlab advanced Primus 96 Hamburg thermal cycler (Applied Bio systems). Universal primer pair 8F forward 5’-AG (A/G) GTTTGATCCTGGCT-3’) and 1492R- reverse, 5’-CGGCTACCTTGTTACGACTT-3’ were used (Lane, 1991). DNA solutions were amplified by the polymerase chain reaction (PCR) with the first denaturing at 95°C for 5 min, the second at 95°C for 30 s, annealing at 55°C for 45 s, and extension at 72°C for 45 s, for 35 cycles. The product was stored at -20°C. PCR amplification was confirmed using Gel electrophoresis that visualized the gel under UV trans-illuminator. Screening for bacterial diversity was conducted by sequencing using Basic Local Alignment Search Tool (BLAST) technique.
RESULTS AND DISCUSSION
Biomethanation profiles
Cumulative biogas production and digester pH for anaerobic digestion of 150 g WH and RSW, separately and in co-digestion at 70:30 ratio varied with retention time as depicted in Figure 2a and b, respectively. At 60 days retention time, WH yielded 19.6 L CH4 kg-1 about 47% of the 42 L CH4 kg-1 yield for RSW. However, co-digestion of WH with 30% RSW increased the biogas yield to 40 L CH4 kg-1 or 95% of the RSW yield indicating synergy in co-digestion.
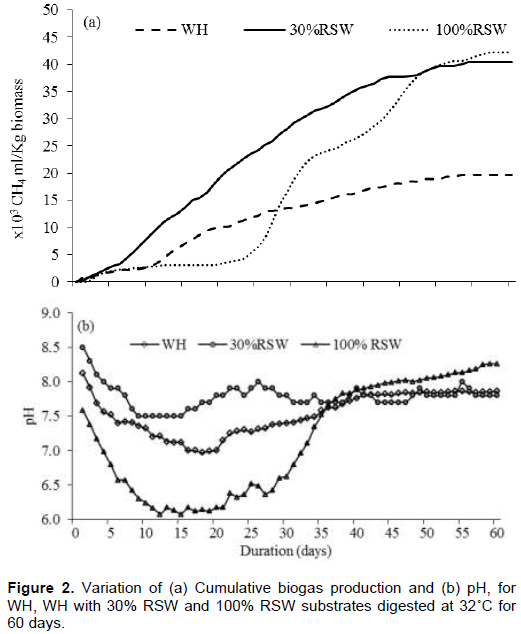
Biomethanation of RSW digested separately experienced stagnation for 20 days from days 5 to 25, which corresponded to a drop in pH from alkaline to slightly acidic (6.2). Comparatively, WH samples experienced stagnation of only 7 days from days 5 to 12 with the pH remaining alkaline, above 7.0. The co-digestion sample exhibited uniform biogas production with no stagnation and pH remaining above 7.4.
The biogas production profiles depicted the output of methanogens, the last anaerobic digestion step, which, however, depended on the rates of the preceding steps of hydrolysis, acidogenesis and acetogenesis, and the operating environment for methanogens. On the other hand, the pH profiles indicated the operating environment for the microbes involved in the digestion. Consequently, the profiles can help deduce comparative intensities of reactions especially those of hydrolysis and acidogenesis as compared to methanogenesis. Accordingly, the pH drop for RSW to acidic levels was associated with rapid hydrolysis and acidogenesis processes that produced volatile fatty acids without matching consumption by acetogenic and methanogenic microorganisms. On one hand, the result indicated that the RSW was amenable to rapid hydrolysis, which can be attributed to significant concentrations of carbohydrates, proteins and cellulose (Omondi et al., 2019). On the other hand, lack of balance between the rates of fermentation and methanogenesis steps resulted in accumulation of acids in the reactor. The acids reduced reactor pH and may have had inhibitory effect on methanogens affecting the efficiency of the biomethanation process (Yang et al., 2011; Ingrid et al., 2014). Restoration of alkaline conditions for RSW occurred probably by growth of acid tolerant acetogenic organisms that consumed the acids, raising pH sufficiently to allow the action of methanogens.
Co-digestion of WH with RSW at 70:30 ratio, maintained a more alkaline pH suggesting that co-digestion matched the rate of acidogenesis to that of methanogenesis. The result may be attributed partly to dilution of RSW substrate by WH thereby reducing the rate of hydrolysis. It may also have been contributed by collating of process parameters such as supply of buffer capacity to allow faster methanogenesis, supply of methanogens by ruminal waste, and improvement of C/N and C/P ratios. The result was uniform biogas production that improved yield.
Morphological characteristics of bacteria isolates
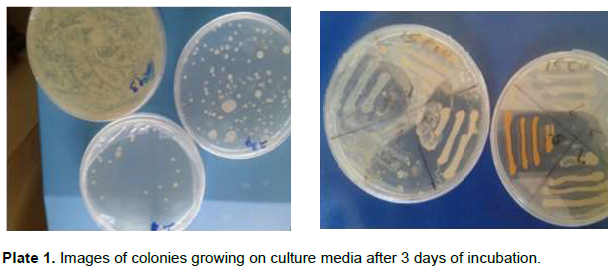
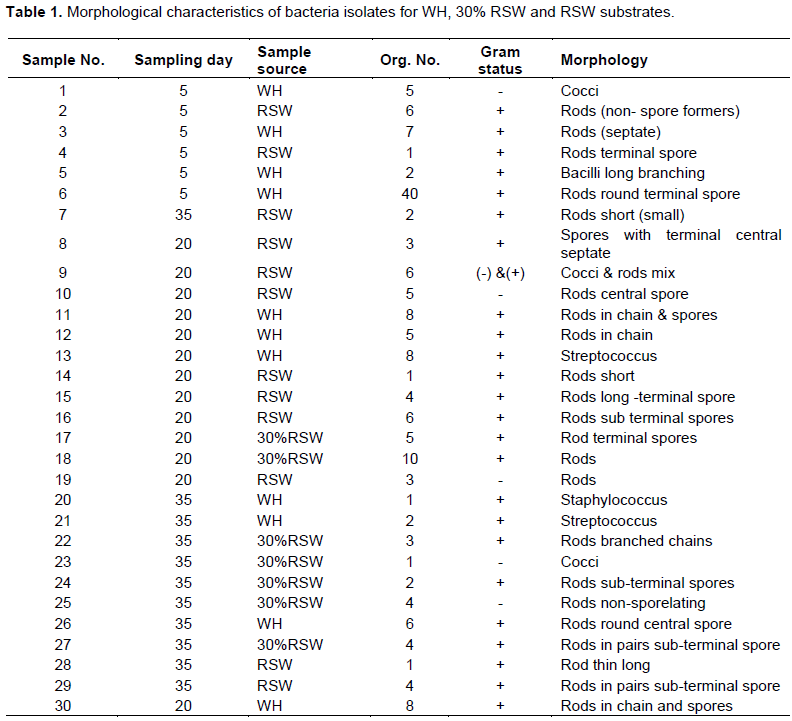
The morphology of microbial communities present in the digestion of WH and RSW separately and in co-digestion at 30% RSW were studied by isolation of colonies from digester sludge. Most colonies aggressively grew within 2 to 3 days of incubation (Plate 1). Morphologies of the isolates varied between different colonies and included circular, covering the entire plate, flat and filamentous morphologies. The colors ranged from white to cream and bluish clear with elevations categorized as cocci, short rods, long rods, bacilli long and streptococcus (Table 1). About 77% of the isolates were Gram positive, mostly spore-forming, while 23% were Gram negative. The result indicated possible dominance of the largely Gram positive spore-forming Firmicutes phyla that comprises the Bacilli and Clostridia classes. Overall, the morphological characteristics did not reveal definite distinction between the two substrates and their mixture or trend with retention times. Instead, morphological characteristics were distributed generally across substrates and retention times.
Microbial diversity
Screening for bacterial diversity was conducted using Basic Local Alignment Search Tool (BLAST) for anaerobic digestion for days 5, 20 and 35 that represented acclimatization, lag and active biomethanation stages. All the samples had clear DNA gel and PCR amplified images as visualized under UV and UV–trans-illuminator, respectively (Figures 3 and 4).
Table 2 presents the most likely identity of the observed bacteria communities in the three reactors. Generally, diverse Bacillus spp. dominated the alkaline pH of acclimatization and active biomethanation phases, while Lysinibacillus and Solibacillus species dominated acidic and low alkaline pH, which corresponded with depressed biogas yield.
At day 5, the microbes observed in the WH reactor were Alcaligenes faecalis, Bacillus spp. (thurigiensis, toyonensis and cereus). The A. faecalis, is a Gram-negative bacterium of Proteobactria phylum, which is known for its ability to aerobically desaturate saturated fatty acids to mono-saturated fatty acids (Ghaneker and Nair, 1973). This bacterium may have used the initial oxygen in the reactor to degrade the fatty acids, which in concert with other microbes, contributed to the initial gas production noted in all three reactors (Figure 2a). Nevertheless, rapid hydrolysis of the more amenable substrate such as carbohydrate especially in RSW resulted in net acid production over consumption by methanogens leading to a drop in pH (Figure 2b). The reduced pH down to 6.2 for RSW had an inhibitory effect on the methanogens stagnating biogas production in the single substrate reactors.
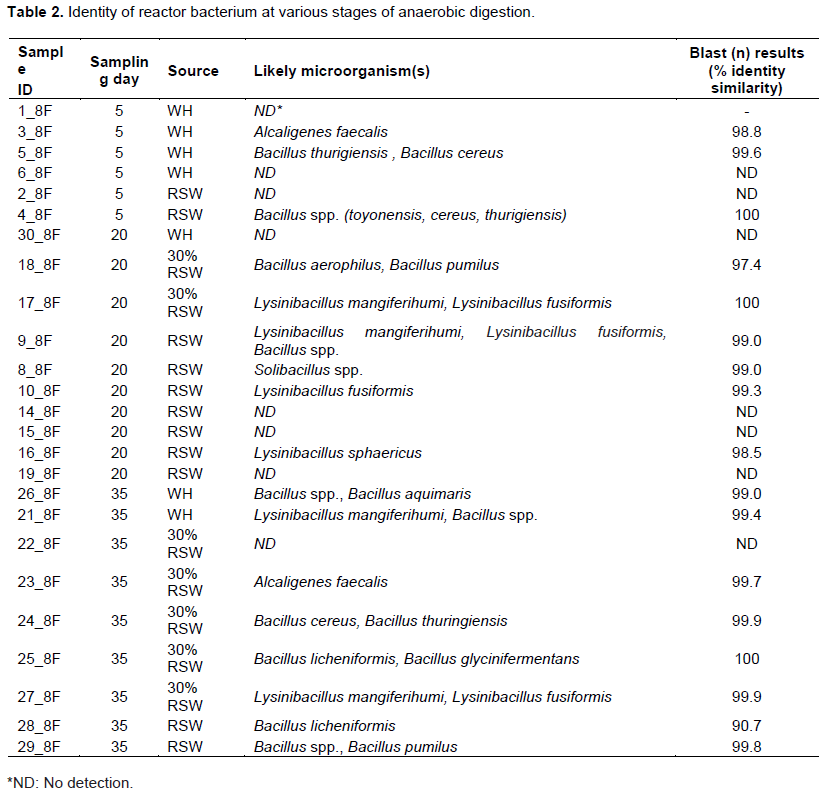
Anaerobic digestion Day 20 was characterized by stagnation of biomethanation for RSW in acidic pH but active biomethanation for co-digestion and WH substrates in an alkaline environment. Microbes present in the RSW reactor were predominantly Lysinibacillus spp., Mangiferihumi, Sphaericus, and Fusiformis, and Solibacillus genus that are of the same Class bacilli as the Bacillus genus in Firmicutes phylum. Both genuses were observed by Zainudin et al. (2014) in decomposition of the ligneous empty fruit bunch of oil palm. Lysinibacillus spp. are a diverse group of bacteria that inhabit various environments including farming soil and factory wastewater. Ahmed et al. (2007) observed that they grow in the pH range 5.5 to 9.5 with an optimum of 7.0 to 8.0. The bacteria are Gram-positive, mesophilic, rod-shaped and typically facultative anaerobes (Todar, 2012). The observation of Solibacillus spp. at acidic pH of 6.0 to 7.0 was consistent with observation by Sielaff et al. (2017) that a strain of the genus grew at a pH range of 6.0 to 10 and that it was negative for hydrolysis of starch and casein which ruled out its role in hydrolysis and acidogenesis steps. Instead, Sielaff et al. (2017) found the strain was differentiated from other species by hydrolysis of gelatin and utilization of several acids including the L-aspartic acid, guadinine HCl and quinic acid as sole carbon substrates. Similar utilization of volatile acids by Solibacillus spp. in the RSW reactor may have been responsible for removal of acidity and recovery of the biomethanation process.
Acidic pH conditions negatively impact on the growth of the Bacillus spp. hindering anaerobic process (Ivanova et al., 2003). Some studies have indicated that benzoic and propionic acids as well as esters of p-hydroxybenzoic acid (parabens) can inactivate Bacillus (Wipat and Colin, 1999). The dominance of Lysinibacillus and Solibacillus genera in acidic environment and the near absence of Bacillus genera indicated a shift in microbial community in the lag phase to adjust to the acidic pH. However, the acidic pH also inhibited the action of methanogens leading to stagnation of biomethanation.
Biomethanation resumed in the WH and RSW reactors, after 7 and 20 days of stagnation, respectively. Taconi et al. (2008) found that the methanogens can operate in an acidic environment provided they had sufficiently long retention time, which they interpreted was to acclimate. However, rather than acclimatization of methanogens, the emergence of Lysinibacillus and Solibacillus observed in this study suggests a shift in the microbial community to species that consumed acidity. Co-digestion of WH with RSW avoided the drop in pH for the co-substrates and, therefore, the change microbial community, which allowed the Bacillus spp. to have continuous biomethanation action in consort with methanogens resulting in steady biogas production.
At day 35, all the reactors had alkaline pH of about 7.4 and depicted active biomethanation. The observation is in line with typical occurrence of the active biomethanation pH of 7.2 to 8.2 that is attributed to the buffer effect of increased ammonium concentration (Kossmann et al., 2007). During this stage of digestion, the reactors had diverse microbial community dominated by species of the Bacillus genus including Bacillus aerophilus, Bacillus pumilus, Bacillus cereus, Bacillus thuringiensis, Bacillus licheniformis, Bacillus glycinifermentans, Bacillus aquimaris, and Staphylococcus xylosus, but also some Lysinibacillus. These species are known to grow in an alkaline environment. For example, according to Ahmed et al. (2007), B. licheniformis is usually cultured in alkaline conditions to obtain protease for use in biological laundry detergent that has an optimum pH at around 9 and 10 while Bacillus fusiformis and Bacillus sphaericus are known to have an optimum pH range of 7.0 to 8.0. Bacillus genera play an important role in biogas syntrophic reactions, whereby they degrade volatile fatty acids, alcohols and acetate to produce H2, which is then used by hydrogenotrophic methanogens to produce methane (Horváth et al., 2016). Rabah et al. (2010) observed Bacillus megaterium, B. licheniformis, and B. pumilus in biogas production using abattoir waste as the inoculum. The results are also in line with that of Onwuliri et al. (2016) observations that microbial isolates responsible for biogas production from cow dung included B. licheniformis, Escherichia coli and Clostridium spp.
CONCLUSION
Co-digestion of WH with 30% RSW increased the biogas yield from 19.6 L CH4 kg-1, which was 47% of that of RSW to 40 L CH4 kg-1 or 95% of that of RSW indicating synergy in co-digestion. After the acclimatization period, rapid acidogenesis without matching methanogenesis decreased pH to acidic pH range 6.0-7.0 for RSW, which suppressed methanogenesis for up to 20 days. The microbial community shifted from initial dominance of Bacillus spp. to acetogenic Lysinibacillus and Solibacillus spp. bacteria that consumed the acids, allowing active biomethanation. Co-digestion of the WH with RSW collated process parameters and avoided significant drop in pH and the corresponding change of microbial community resulting in steady biogas production. Consequently, the synergy in co-digestion was attributed to both collation of process parameters and the maintenance of alkaline pH that is conducive for the microbes involved in biomethanation. Future work should study the dynamics of the methanogenic archaea, which was not covered in this study.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Ahmed I, Akira Yokota, Atsushi Yamazoe and Toru Fujiwara, (2007). Proposal of Lysinibacillus boronitolerans gen. nov. sp. nov., and transfer of Bacillus fusiformis to Lysinibacillus fusiformis comb. nov. and Bacillus sphaericus to Lysinibacillus sphaericus comb. nov. International Journal of Systematic and Evolutionary Microbiology 57:1117-1125. |
|
|
Ahring, BK, Hofman-Bang J, Zheng D, Westerman P and Raskin L (2003). Molecular Ecology of Anaerobic Reactor Systems. Advances in Biochemical Engineering/ Biotechnology Vol. 81 springer-verlag Berlin Heildeberg. |
|
|
Akarsubasi AT, Ince O, Kirdar B Oz NA, Orhon D, Curtis TP, Head IM, Ince BK (2005). Effect of wastewater composition on archaeal population diversity. Water Research 39:1576-1584. |
|
|
Czatzkowska M, Harnisz M, Korzeniewska E, Koniuszewska I (2020). Inhibitors of the methane fermentation process with particular emphasis on the microbiological aspect: A review. Energy Science and Engineering 8:1880-1897. |
|
|
Esposito G, Frunzo L, Giordano A, Liotta F, Panico A, Pirozzi F (2012). Anaeriobic co-digestion of organic wastes. Reviews in Environmental Science and Bio/Technology 11(4):325-341. |
|
|
Gerber M, Span R (2008). An Analysis of Available Mathematical Models for Anaerobic Digestion of Organic Substances for Production of Biogas. International Gas Union Research Conference, 30 s. |
|
|
Ghaneker AS, Nair PM (1973). Evidence for the Existence of an Aerobic Pathway for Synthesis of Monounsaturated Fatty Acids by Alcaligenes faecalis. Journal of Bacteriology 114:618-624. |
|
|
Gichuki JR, Omondi P, Boera T, Okorut AS, Matano T, Jembe T, Ofulla A (2012). Water Hyacinth Eichhornia crassipes (Mart.) Solms-Laubach Dynamics and Succession in the Nyanza Gulf of Lake Victoria (East Africa): Implications forWater Quality and Biodiversity Conservation. The Scientific World Journal. Volume 2012, Article ID 106429, 10 pages |
|
|
Heeg K, Pohl M, Mumme J, Klocke M, Nettmann E (2014). Microbial communities involved in biogas production from wheat straw as the sole substrate within a two-phase solid-state anaerobic digestion. Systematic and Applied Microbiology 37(8):590-600. |
|
|
Horváth IS, Tabatabaei M, Karimi K, Kumar R (2016). Recent updates on biogas production-overview. Biofuel Research Journal 10:394- 402. |
|
|
Ingrid HFW, Andreas W, Christian E, Insam H (2014). Investigation into the effect of high concentrations of volatile fatty acids in anaerobic digestion on methanogenic communities. Waste Management Volume 34(11):2080-2089. |
|
|
Ivanova N, Sorokin A, Anderson I, Galleron N, Candelon B, Kapatral V, Bhattacharyya A, Reznik G, Mikhailova N, Lapidus A, Chu L (2003). Genome sequence of Bacillus cereus and comparative analysis with Bacillus anthracis. Nature 423(6935):87-91. |
|
|
Kossmann W, Ponitz U, Habermehl S, Hoerz T, Kramer P, Klingler B, Kellner C, Wittur T, Lopotek F, Krieg A, Euler H (2007). Biogas digest. Information and Advisory Service on Appropriate Technology (ISAT) 1:1-46. |
|
|
Lane DJ (1991). 16S/23S rRNA Sequencing. In: Stackebrandt, E. and Goodfellow, M., Eds., Nucleic Acid Techniques in Bacterial Systematic, John Wiley and Sons, New York, pp. 115-175. |
|
|
Mata-Álvare J, Joan D, Maycoll R, Sergi A (2014). A critical review on anaerobic co-digestion achievements between 2010 and 2013. Renewable and Sustainable Energy Reviews 36:412-427. |
|
|
McHugh S, Carton M, Mahony T, O'Flaherty V (2003). Methanogenic population structure in a variety of anaerobic bioreactors. FEMS Microbiology Letters 219(2):297-304. |
|
|
Meyer T, Edwards EA (2014). Anaerobic digestion of pulp and paper mill wastewater and sludge Water Research 65:321-49. |
|
|
Omondi EA, Njuru PG, Ndiba PK (2019). Anaerobic Co-Digestion of Water Hyacinth (E. crassipes) with Ruminal Slaughterhouse Waste for Biogas Production. International Journal of Renewable Energy Development 8(3):253-259. |
|
|
Onwuliri F, Onyimba I, Nwaukwu I (2016). Generation of Biogas from Cow Dung. Journal of Bioremediation and Biodegradation S18:1-2. |
|
|
Rabah A, Baki A, Hassan L, Musa M, Ibrahim A (2010). Production of biogas using abattoir waste at different retention time. Science World Journal 5(4):23-26. |
|
|
Raskin L, Poulsen LK, Noguera DR, Rittmann BE, Stahl DA (1994). Quantification of methanogenic groups in anaerobic biological reactors by oligonucleotide probe hybridization. Applied Environmental Microbiology 60:1241-1248. |
|
|
Rivière D, Desvignes V, Pelletier E (2009). Towards the definition of a core of microorganisms involved in anaerobic digestion of sludge. ISME Journal 3:700-714. |
|
|
Sielaff ACS, Kumar RM, Pal D, Mayilraj S and Venkateswaran K (2017). Solibacillus kalamii sp. nov., isolated from a high-efficiency particulate arrestance filter system used in the International Space Station. International Journal of Systematic Evolution in Microbiology 67:896-901. |
|
|
Suksong W, Kongjan P, Prasertsan P, Imai T, O-Thong S (2016). Optimization and microbial community analysis for production of biogas from solid waste residues of palm oil mill industry by solid-state anaerobic digestion. Bioresource Technology 214:166-174. |
|
|
Taconi Katherine A, Mark E. Zappi W. Todd French, Lewis R. Brown (2008). Methanogenesis under acidic pH conditions in a semi-continuous reactor system. Bioresource Technology 99:8075-8081. |
|
|
Taherzadeh MJ, Karimi K (2007). Acid-based hydrolysis processes for ethanol from lignocellulosic materials, a review. Biology Research 2:472-499. |
|
|
Takashi N, Yuji S (2011). Oligonucleotide primers, probes and molecular methods for the environmental monitoring of methanogenic archaea. Microbiology and Biotechnology 4(5):585-602. |
|
|
Tao R, Zeng R, Xiao-Yan Y, Zhang S, Yang ZH (2016). Water Hyacinth (Eichhornia crassipes) Biomass as a Biofuel Feedstock by Enzymatic Hydrolysis. BioResources 11(1): 2372-2380. |
|
|
Thikra AA (2013). Evaluation of methods for the extraction and purification of DNA of cultured Lactobacillus colony isolated from dairy products. International Journal of Applied Microbiology 1:20-25. |
|
|
Todar K (2012). Staphylococcus aureus and Staphylococcal Disease. In Todar's Online Textbook of Bacteriology. |
|
|
Wipat Anil, Colin RH (1999). The Bacillus subtilis genome sequence: the molecular blueprint of a soil bacterium. FEMS Microbiology Ecology 28(1):1-9. |
|
|
Yang CW, Zhang RH, Liu Q, Li BB, Li SF, Wang CJ, Song QCLA (2011). Phylogenetic diversity and metabolic potential of activated sludge microbial communities in full-scale wastewater treatment plants. Environmental Science and Technology 45:7408-7415. |
|
|
Zainudin MHM, Hassan MA, Shah UKM, Abdullah N, Tokura M, Yasueda H, Shirai Y, Sakai K, Baharuddin AS (2014). Bacterial Community Structure and Biochemical Changes associated with composting of lignocellulosic oil palm empty fruit bunch. Bioresouces 9(1):316-335. |
|
|
Zeeman G, Sanders W (2001). Potential of anaerobic digestion of complex waste (water). Water Science and Technology 44(8):115-122. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


