Review

ABSTRACT
Currently, inorganic nitrogen fertilizer becomes a serious threat to the environment and human health. Thus, finding of alternate source of nitrogen is a viable option in assuring of sustainable agricultural system. Biological nitrogen fixation is a critical and key process in sustainable agricultural systems in tropical soils, which are frequently deficient in N and susceptible to leaching of plant nutrients. This process transforms atmospheric nitrogen to ammonia, nitrate and nitrogen dioxide. Several key abiotic and biotic factors limit legume productivity and biological nitrogen fixation in world agriculture, especially in sub-Saharan Africa. Within the soil, rhizobia frequently encounter various stresses that affect their growth, their initial steps of symbiosis and the capability of nitrogen fixation. Biotic and abiotic stresses impose a major threat to agriculture and symbiotic nitrogen fixation is dependent on host cultivar and rhizobia, but as well may be limited by pedoclimatic factors. The most common factors affecting biological nitrogen fixation and symbiosis activity in western parts of Ethiopia are soil acidity, quality of inoculants and low soil fertility. In most cases, the microsymbiont is the more affected partner, with plants growing on mineral N usually less sensitive to these stresses. Thus, it can be concluded that, particularly in a western part of Ethiopia, many studies should be focused on acidity related constraints on biological nitrogen fixation, screening of acid tolerant inoculants and low soil fertility improvements to enhance biological nitrogen fixation in smallholder farming system.
Key words: Abiotic and biotic factors, legume, Rhizobium, symbiosis.
INTRODUCTION
The population of sub-Saharan Africa is projected to double in coming 2050 and increases in food production are much needed (World Bank, 2014). As the potential to expand agricultural land is limited in many areas with high population densities, sustainable intensification of agricultural production is crucial (Ronner et al., 2016). A potential pathway for sustainable intensification is the integration of grain legumes in farming systems (Peoples et al., 1995). Legumes have the capacity to fix nitrogen from the air in symbiosis with Rhizobium bacteria. Legumes can therefore contribute to improved soil fertility through biological nitrogen fixation and enhance nutrient release from organic matter decompositions in maize dominated cropping systems in Western Ethiopia.
Biological nitrogen fixation (BNF) is the process in which nitrogen gas (N2) from the atmosphere is incorporated into the tissue of legume plants, with the help of soil microorganisms. It is important for smallholder farmers as it is relatively cheaper source of N compared to inorganic fertilizers, less prone to losses through leaching and denitrification (Mhango et al., 2017). The ability of legumes to fix N2 and its ability to produce nodules, has brought about its important and uniqueness (Wilcox and Shibles, 2001). During formation and emergence of root hairs, N2 fixation is affected by many factors such as the presence and density of nodulating bacteria in the root zone, the physical and chemical properties such as humidity, temperature, and salt concentration in the soil, pH levels and deficiencies of several mineral nutrients (Abdul-Jabbar and Saud, 2012).
Legumes are unique for their ability to fix nitrogen from atmosphere by symbiotic relationship with Rhizobium bacteria (Giller, 2001). Rhizobia require a plant host; therefore, they cannot independently fix nitrogen. These bacteria are located around root hair and fix atmospheric nitrogen using particular enzyme called nitrogenase (Coskan and Dogan, 2011). When this mutualistic symbiosis is established, rhizobia can use plant resources for their own reproduction whereas fixed atmospheric nitrogen is used to meet nitrogen requirement of both itself and the host plants. Supply of nitrogen through biological nitrogen fixation has ecological and economical benefits (Ndakidemi et al., 2006). BNF offers an economically attractive and ecologically sound means of improving crop yield, reducing external N inputs and enhancing the quality of soil resources which consequently reduce the dependence on mineral fertilizers that could be costly and unavailable to smallholder farmers (Massawe et al., 2016). Leguminous crops can hold promise in this regard.
The use of legumes is a promising option of increasing yields, profits and nutrition for smallholder farmers in sub-Saharan Africa (SSA), especially in areas where soil nutrient availability is low. They increase soil fertility due their ability to establish symbiotic associations with soil microorganisms, known as rhizobia, capable of ï¬xing nitrogen from the atmosphere (Ahmad et al., 2013; Mhango et al., 2017). However, they are frequently exposed to biotic and abiotic stress conditions such as legume genotype, pesticide, various climatic and edaphic factors as well as the status of native rhizobia in the soil. Inoculation might be necessary to overcome the latter constraint. Solomon et al. (2012) indicated that legumes can obtain between 50 and 80% of their nitrogen requirements through BNF. However, the success of inoculation depends not only on high quality inoculants and good inoculation practices but also on the establishment of effective and efficient BNF through optimization of the factors that affect its performance such as legume genotype, climatic, edaphic and management factors (Giller, 2001).
Therefore, BNF is important for smallholder farmers as it is relatively cheaper source of nitrogen compared to inorganic fertilizers, less prone to losses through leaching and denitrification. Recently, a diversity of legumes including soybean, common bean, groundnut, pigeon pea and cowpea are the major food legume crops that have been produced in sub-humid tropics of western Ethiopia but the average yield of these legumes are below their potential yield. Thus, there is a need to conduct a study that will explore the response of legumes inoculated with rhizobia, soil acidity amelioration, and response to nutrients (both macronutrients and micronutrients) on nitrogen fixation so as to add knowledge on existing information. Furthermore, studies are also required to quantify the amount of nitrogen that can be fixed by specific legumes in different environments and cropping systems. Therefore, the objective of this review was getting an overview of biotic and abiotic stresses that limit biological nitrogen fixation in legumes particularly in western Ethiopia.
ABIOTIC FACTORS
BNF is one of the important soil microbial activities which is affected by all ongoing processes in soil as well as other soil microorganisms. BNF process depends on the occurrence and survival of Rhizobium in soils and also on its efficiency (Adamovich and Klasens, 2001). The rate of nitrogen fixation was affected by many different physiological and environmental factors in soil, such as temperature, water holding capacity, water stress, salinity, nitrogen level, pH and other nutrients. Many of these factors, including temperature, affect many aspects of nitrogen fixation and assimilation, as well as factors such as respiratory activity, gaseous diffusion and solubility of dissolved gases, which ultimately affect plant growth (Coskan and Dogan, 2011).
Establishment of effective N2 fixing symbiosis between legumes and their N2 fixing bacteria is dependent upon many environmental factors, and can be greatly influenced by farm management practices (Peoples et al., 1995). There are several environmental factors affecting BNF. The severe environmental conditions such as salinity, unfavorable soil pH, nutrient deficiency, mineral toxicity, extreme temperature conditions, low or extremely high levels of soil moisture, inadequate photosynthesis, and disease conditions can affect the plant growth and development. As a result, even the persistent Rhizobium strains will not be able to perform root infection and N2-fixation in their full capacity (Panchali, 2011).
High soil temperatures
Temperature has a profound influence of N2 metabolism. Little activity is observed at low temperature and warming promotes the microbial N2 fixation and uptake of fixed gas (Saha et al., 2017). The plant nitrogenase activity reduces dramatically as a result of formation of ineffective nodules at high temperature (40°C) (Hungria and Franco, 1993; Hungria and Vargas, 2000). In addition, the relative activity of the rhizobia is altered by temperature, so that Rhizobium that is highly effective at specified range of temperature is less active at another range of temperatures. For these reasons, greater nitrogen gains probably can be achieved by improvements in the heat resistance of the symbiosis. The optimum temperatures for growth in culture vary among strains and species, values between 27 to 39°C have been noted. The maximum temperatures are generally 35 to 39°C, but proliferation may take place up to 42°C (Al-Falih, 2002). Rhizobial survival in soil exposed to high temperature is greater in soil aggregates than in non-aggregated soil and is favored by dry rather than moist conditions (Zahran, 1999).
Soil moisture
The moisture stress can adversely affect the nodule functions. The drought conditions can reduce nodule weight and nitrogenase activity. After exposure to the moisture stress for 10 days, the nodule cell wall started to degrade resulting in senescence of bacteroids (Ramos et al., 2003). The occurrence of rhizobial populations in desert soils and the effective nodulation of legumes growing there in emphasize the fact that rhizobia can exist in soils with limiting moisture levels. Viable strains of Rhizobium usually cannot tolerate or function under high levels of osmotic stress caused by drought. N2-fixing legumes are especially sensitive to water deficit and other environmental stresses, with drought being one of the major environmental factors affecting plant productivity (Zahran, 1999; Niste et al., 2013). Nodules and N2 fixation response to water stress depends on the stage of plant development. Water stress during growth has a direct effect on the development of nodules than in other stages and the possibility of recovery is almost impossible. Thus, in Western Ethiopia, there is no shortage of moisture that limits BNF because the area receives enough average annual rainfall.
Pesticide applications
The compatibility of rhizobia with pesticides is poorly understood except for fungicides. Insecticides have little adverse effect on nodulation when not directly applied on seed. The effect of herbicides on rhizobial survival is unknown. However, studies have shown the incompatibility between Bradyrhizobium species and practices of seed treatment with fungicides and insecticides in soybean (Rampim et al., 2015). Seed treatment with pesticides, especially fungicides may rapidly kill the inoculated bacteria, affecting the establishment and outcome of the symbiosis (Araujo et al., 2017). The compatibility of legume strains with different pesticides was not studied in Ethiopia in general and in western Ethiopia in particular. All farmers in maize belt areas of western Ethiopia are using pre-emergence herbicide like glyphosate in order to control weeds. Due to the variability of the effect it is recommended to test the particular Rhizobium inoculum and its behavior in respect to the product to be used, before application. The effect of pesticide on N2 fixation should be minimized by separate placement of rhizobia and pesticide. Thus, the compatibility of herbicides with legume inoculants should be studied to generate information for herbicide users, especially on legume fields.
Soil acidity
Acidity refers to concentration of hydrogen cations in a soil solution (FAO, 2006). The natural pH of a soil depends on the nature of the material from which it was developed (TSO, 2010). In soils of low pH, containing high amounts of Al and Fe oxides, P is deficient in the soil solution because it is precipitated or surface adsorbed with Al and Fe as insoluble compounds (Kanyanjua et al., 2002). Several other essential plant nutrients, which are present in the soil solution as cations, are deficient. In acid soils, soybean is affected directly and indirectly. These effects include injury on plant roots therefore reducing water and nutrient uptake, reduced availability of essential plant nutrients, toxicity of Al and Mn; and survival of microorganisms in the soil (Crawford et al., 2008).
Soil acidity has become a serious threat to crop production in most highlands of Ethiopia in general and in the South, South western and western part of the country in particular. About 41% of potential arable land of Ethiopia is acidic (Workneh, 2013). Currently, it is estimated that about 67% of the total arable land of Wollega is affected by soil acidity (Abdenna et al., 2007). Soil acidity has long been known to decrease symbiotic nitrogen ï¬xation in legumes, negatively affecting growth and yield, especially in plants depending exclusively on symbiosis to acquire nitrogen (Mohammadi et al., 2012; Bekere et al., 2013). Soil acidity constrains symbiotic N2 fixation (Munns, 1986), limiting Rhizobium survival and persistence in soils and reducing nodulation and causes nutrient imbalance (Foy, 1984; Abubakari et al., 2016).
Increased soil acidity may lead to reduced yields, poor plant vigour, and nodulation of legumes. The activities of rhizobia in the legume root rhizosphere can be reduced by the extreme soil pH or low soil pH. The characteristics of highly acidic soils (pH < 4) are low level of phosphorus, calcium, and molybdenum along with aluminum and manganese toxicity, which affects both plant and the rhizobia (Bordeleau and Prevost, 1994; Ferreira et al., 2016). As a result, under low soil pH conditions, nodulation and N fixation are more severely affected than the plant growth.
To enable crop production in acid soils, several means to correct nutrient deficiency can be adopted. These include liming, addition of organic matter, and fertilization with mineral fertilizer (Onwango et al., 2010; Masarirambi et al., 2012). Liming reduces Al3+ and H+ ions as it reacts with water leading to the production of OH- ions, which react with Al3+ and H+ in the acid soil to form Al(OH)3 and H2O. The precipitation of Al3+ and H+ by lime causes the pH to increase, enhances microbial activity and nutrient availability (Onwongo et al., 2008). Soybean as leguminous crop relies on microbial nitrogen fixation as source of N. However, under acid soils, the population of rhizobia bacteria is reduced and consequently nodulation and N fixation is impaired. This affects negatively on crop nutrition and yields. Therefore, liming acid soils for legume production improves soils condition for microorganism development. Mineral fertilizers increase nutrient availability in the soil solution since they are readily available, and the addition of organic matter acts as supply of microorganism’s food enhancing their population and therefore mineralization (Crawford et al., 2008).
Moreover, Workneh (2013) in South Western Ethiopia also found positive response of soybean grain yield to lime application combined with Bradyrihizobium and N-fertilizer. Likewise, Zerihun and Tolera (2014) reported that increased faba bean yield ranging from 11% to 23% as the function of increasing lime application rates up to 6 t ha-1. Mesfin et al. (2014a, b) and Hirpa et al. (2013) also observed that considerable differences among common bean genotypes as a result of lime application. For this reason, particularly for western Ethiopia we call up many studies on acid tolerant legume cultivars and Rhizobium strains and soil acidity amendment experiments to achieve intermediate pH at which availability of essential plant nutrient increased.
MACRONUTRIENTS
Nitrogen
Nitrogen (N) is essential for plant growth, participating in several metabolic pathways and in the synthesis of molecules such as proteins, nucleic acids, hormones, and chlorophyll (Saturno et al., 2017; Bruijn, 2016).
Attempts to supplement N2 fixation using inorganic fertilizer have not been successful because the addition of fertilizer-N tends to substitute for, rather than supplement, N2 fixation. Nevertheless, it is generally accepted that symbiotically fixed N2 is inadequate for realizing maximum seed yield, and that an application of small amounts of “starter” fertilizer N is needed to establish seedlings and promote early dinitrogen fixation (Sogut et al., 2013). However, application of large quantities of inorganic N inhibits the growth of rhizobia, nodulation and dinitrogen fixation (Coskan and Dogan, 2011). Herridge et al. (1984) and Goi et al. (1993) stated that under soils low in mineral N, a moderate dose of starter-N has been demonstrated to stimulate seedling growth and subsequently N-fixation. Similarly, Hansen (1994) reported that inorganic N is required by legume plants during the ‘nitrogen hunger period’ for their nodule development, shoot and root growth before the onset of N2-fixation process.
On the other hand, presence of high N levels in the soil inhibits both nodule formation and nitrogenase activity (Saturno et al., 2017). Weisany et al. (2013) reported that mineral N in the soil inhibited symbiotic nitrogen fixation but it was relative to start of nodulation and N2 fixation at early vegetative growth at low concentration. It is a well- established fact that, when legumes are grown in soils high in available nitrogen, the nitrogen fixation rate is reduced (Solomon et al., 2012; Saha et al., 2017 ). Inhibitory effect of nitrate causes the reduction of capillary roots development as well as preventing particular infection’s strands, indicating a shift from symbiotic to inorganic N nutrition (Coskan and Dogan, 2011). Therefore, it is possible to apply small amounts of soil N fertilizer, which may increase yield without reducing the amount of nitrogen fixed (Figure 1).
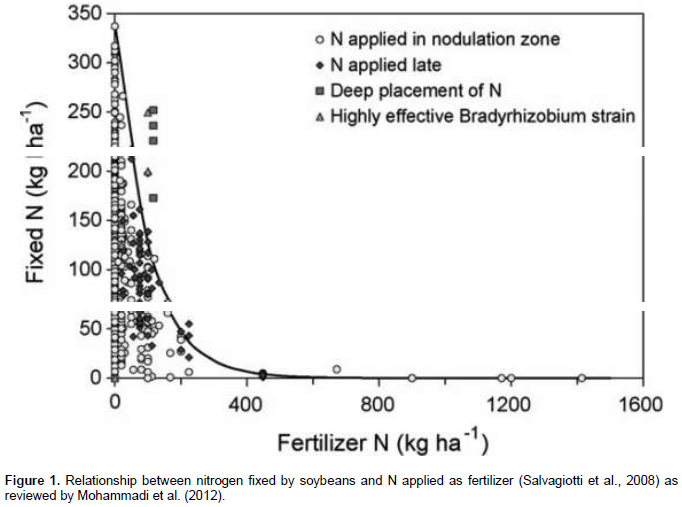
Phosphorus
Phosphorus is used in numerous molecular and biochemical plant processes, particularly in energy acquisition, storage and utilization. P plays an important role in N-fixing process, as adenosine tri-phosphate (ATP) is required in large quantities for legumes to undergo N2 fixation (Mmbaga et al., 2014). The deficiency of phosphorous supply and availability remains a severe limitation on nitrogen fixation and symbiotic interactions. Availability of phosphorus in the soil influences the efficiency of Rhizobium that fixes atmospheric nitrogen in association with nodulating legumes as it is directly involved in BNF via legume-Rhizobium symbiosis. This has attracted considerable research attention world-wide due to its economic viability for resource poor farmers and environmental friendliness (Mahamood et al., 2009; Mmbaga et al., 2014). Phosphorus influences nodule development through its basic functions in plants as an energy source. Furthermore, P increases the number and sizes of nodules and the amount of nitrogen assimilated per unit weight of nodules. Its deficiency in legume plants results in reduced nodule mass, N fixation, and low yield. In western parts of Ethiopia, low soil P availability, due to soil acidity and its high fixation, is a limiting factor to crop production (Zerihun et al., 2015; Abebe, 2017).
Potassium
Potassium plays an important role in the process of nitrogen fixation (Nyoki and Ndakidemi, 2016). Potassium is essential in photosynthesis, as it maintains and balances the electrical charges at ATP production site, and also helps to promote translocation of photosynthetic substances (carbohydrate) to storage organs (fruits or roots) (Uchida, 2000). It facilitates cell division in young meristematic tissue, due to its effect on plant cell turgor and permeability. It is probably best known for its role in the opening of the stomata in the leaves and hence its role in regulating the rate of transpiration (Mmbaga et al., 2014). Due to its essentiality in facilitating plant growth and its function in photosynthesis, potassium application in legume-based production field is inevitable. However, there is no information on legume response to potassium fertilizer in Ethiopia in general and in western Ethiopia in particular because soil test based results showed no deficiency of potassium in Ethiopian soils.
Calcium
Calcium applied to legumes enhanced root development, attachment of rhizobia to root hairs and root nodulation. Inhibition of nodulation is a major limiting factor in N2 fixation of many legume species grown in acid mineral soils. Increase in soil pH by liming is therefore very effective in increasing nodule number, for example in common bean (Bambara and Ndakidemi, 2010). A requirement of calcium has been demonstrated during nitrogen assimilation by cyanobacteria and some species of Azotobacter (Saha et al., 2017). Weisany et al. (2013) reported that legume plants under N2 fixing symbiosis, Ca2+ deficiency decreased nitrogen fixation in nodules and also affects attachment of rhizobia to root hairs and nodulation. Similarly, Hirpa et al. (2013) and Zerihun and Tolera (2014) reported the significant response of common bean and faba bean to the lime application in western Ethiopia. This might be due to the increased availability of calcium which in turn enhances the attachment of rhizobia to root hairs. Therefore, the availability of Ca2+ is an important factor in BNF process.
Sulfur
Sulfur is an essential element for growth and physiological functioning of plants. The sulfur containing amino acids cysteine and methionine play a significant role in the structure, conformation, and function of proteins and enzymes in vegetative plant tissue (Tabatabai, 1986). Sulphur deficiency may also decrease N2 fixation by affecting nodule development and function, reduce leghemoglobin concentration in nodules, and lower ATP concentrations in bacteroids as well as in mitochondria of root nodules. Sulphur is important for nitrogenase activity because the smaller of the two oxygen-sensitive non-heme iron proteins contains a single Fe4S4. Besides nitrogenase, the activities of other important enzymes involved in N2 fixation, such as phosphoenolpyruvate (PEP)-carboxylase, malate dehydrogenase or glutamate synthase, are also reduced when S supply is inadequate.
MICRONUTRIENTS
Molybdenum
Molybdenum is required to the Rhizobium bacteria for proper function of nitrogenase enzyme which involved in nitrogen ï¬xation. Again molybdenum is the cofactor for the enzyme nitrate reductase which involved in nitrogen assimilation (Alam et al., 2015; Chatterjee and Bandyopadhyay, 2015). Therefore, plant N metabolism is closely related to the Mo concentration in soil, especially for leguminous plants (Alam et al., 2015). The application of molybdenum in deï¬cient soil encouraged nitrogen ï¬xation and nodule formation (Rahman et al., 2008). Due to this relationship, Mo-deï¬cient legumes show an unusual proliferation of nodules, which in turn leads to N deï¬ciency (Marschner, 2011). In acid soils, Mo bioavailability is reduced by the interaction of molybdate (MoO42-) with Fe oxides and organic matter (OM) (Makoi et al., 2013). Supplying Mo will not only correct the Mo deficiency manifested in such soils, but also will improve plant growth by enhancing nodulation and N2 fixation. At maturity, beans receiving mineral N had a higher shoot but lower pod dry weight compared with N2-fixing plants supplied with Mo. Therefore, Mo is one of deficient plant nutrients under acidic soil conditions. No study was conducted on the response of legumes to Mo application in western Ethiopia. Thus, there is a need to study the role of Mo on nodulation and nitrogen fixation, particularly for western Ethiopia.
Iron
Legumes, which develop a symbiosis with nitrogen-ï¬xing bacteria, have an increased demand for iron. Iron is required for the synthesis of iron-containing proteins in the host, including the highly abundant leghemoglobin, and in bacteroids for nitrogenase and cytochromes of the electron transport chain (Brear et al., 2013). Iron is an essential nutrient for both legume and its root nodules. Nitrogen fixation it is a component of several enzymes such as nitrogenase, the electron carrier ferredoxin, leghemoglobin and several hydrogenases. The heme component of leghemoglobin has a particularly high Fe requirement (Brear et al., 2013). Therefore, Fe is needed in greater amounts for nodule formation than for host plant growth. Although Fe deficiency does not significantly affect shoot growth in peanut, it severely decreases nodule mass, leghemoglobin concentration, number of bacteroids and nitrogenase activity, compared with plants treated with foliar Fe.
Boron
Legume crops required more amount of boron compared to most ï¬eld crops as boron plays vital role in proper development of reproductive organs. Its deï¬ciency leads to sterility in plants by malformation of reproductive tissues affecting pollen germination, resulting in increased flower drop and reduced fruit set. Again, the boron deï¬ciency could hamper the response to applied nitrogen because nitrogen aggravates the boron demand of the crop (Chatterjee and Bandyopadhyay, 2015). Particularly, legume crops require B in relatively high concentrations for nodule development. The B concentration in nodules is about four to five times higher than in roots. Boron is required for the development of infection threads and nodule cell invasion. In absence of B, the binding of rhizobial cell surfaces to the infection thread wall is inhibited; bacteria cannot progress through the infection thread (Noor and Hossain, 2007). Boron deficiency also causes abortion of infection threads as well as degeneration of cell walls and the membranes surrounding the intracellular bacteroids. These impairments to nodule development result in decreased N2 fixation and necrosis.
Cobalt
Severe Co deficiency reduces infection and retards nodule formation whereas nodule growth rate is not affected by Co supply. Moreover, Co is required for the synthesis of leghemoglobin. Cobalt deficiency affects nodule development and function in various ways. For example, in lupins relying on symbiotic N2 fixation, Co deficiency depresses host plant growth but not nodule mass, which even increases. The most sensitive indicator of Co deficiency is the bacteroid content of nodules. Similarly, Saha et al. (2017) indicated, organisms making use of N2 must have cobalt available to them, although a lesser concentration of this element may be required for growth on combined nitrogen.
Nickel
Nickel (Ni) is an essential micronutrient required for plants’ metabolism due to its role as a structural component of urease and hydrogenase, which in turn perform nitrogen (N) metabolism in many legume species. Seed treatment with cobalt, molybdenum and Bradyrhizobium strains has been widely practiced to improve crops (Lavres et al., 2016). Additionally, seed treatment together with Ni fertilization of soybean might improve the efï¬ciency of BNF, boosting seed dry matter yield, and N content. Seed dry matter yield, aerial part dry matter yield and BNF increased, respectively, by 84, 51 and 15% in relation to the control plants at 45 mg kg-1 Ni via seed treatment (Lavres et al., 2016).
BIOTIC FACTORS
Quality of inoculants
The rhizobia involved in nodulation can influence the percentage and amount of nitrogen fixed by the legume/Rhizobium symbiosis. The effectiveness of BNF depends on the management of other inputs such as nutrient availability, population of rhizobia and soil pH (Keyser and Li, 1992; Ukovich et al., 2008). The process of BNF by legume nodules requires large amounts of P, and its availability is a primary constraint to N2 fixation (Danso, 1992). Deficiencies of soil nutrients, especially P may restrict the development of a population of free-living rhizobia in the rhizosphere, limit the growth of the host plant, restrict nodulation itself, and cause an important nodule function (Danso, 1992). Moreover, limitation of N mineral in the soil tends to enhance fixation by legumes including soybean (Ukovich et al., 2008). The population and activity of rhizobia is highly influenced in acid soils, affecting directly N fixation (Jones and Giddens, 1985). Nodulation of legumes is also affected by the native rhizobial population in the soil. Lack of effective rhizobial strains in the soil restricts nodulation and proper growth of legumes. Currently, in Western Ethiopia the effectiveness of legume inoculants are very poor. This might be due to the poor quality of inoculants, poor survival during storage and death on the legume seeds after inoculation.
Competitiveness of native rhizobia
The proportion of the nodules formed on a particular host is influenced by the competitive ability of an inoculated Rhizobium strain in comparison to indigenous strains, which may vary in their effectiveness. The introduction of effective strains of rhizobia depends on the competition for nodules' sites between the introduced strains and the native population of rhizobia. Thus, a key property of an inoculum strain must be the ability to outcompete the indigenous soil bacteria. Triplett (1990) indicates that a high competitiveness of inoculum strains in comparison with native rhizobia strains is as important as the effectiveness of symbiotic nitrogen fixation itself. At field site, well-adapted native strains, with the low nitrogen-fixing ability and/or high nodulation ability, may dominate the highly efficient, laboratory-tested inoculated strains (Martensson and Gustafsson, 1985). Montañez (2000) reported that inoculation attempts failed to improve legume productivity because the indigenous strains occupied the root nodules rather than the inoculum strains. Thus, nodulation competitiveness is the ability of a given strain to dominate nodulation in the presence of other strains of the same species. There is lack of information on competitiveness of legume inoculants with native rhizobia in western Ethiopia. Therefore, there is a need to conduct research on the competitiveness of major legume inoculants that are elite over the indigenous one is important for western Ethiopia.
STRATEGIES TO ENHANCE BNF PARTICULARLY FOR WESTERN ETHIOPIA
Selection of inoculants for tolerance to soil acidity
In Ethiopia, inoculation of food legumes with rhizobial inoculants is not common practice but could provide an option to increase seed yields in low nitrogen (N) acidic soils. In these acid soils, the selection of acid tolerant rhizobia is one strategy that may increase the performance of legume crops (Abera et al., 2016). However, knowledge about the diversity and symbiotic efficiency of rhizobia nodulating major food legumes in western Ethiopian soils is scanty. Therefore, evaluation of rhizobia strains isolated from Ethiopian soils for their acid tolerance and symbiotic N fixation efficiency is an alternative option for increasing legume yields in western Ethiopia.
Selection of legume genotypes
Genetic stability, especially under conditions of stress factor is essential for any strain recommended for use in commercial inoculants (Hungria and Vargas, 2000). The amount of nitrogen fixed by legumes varies widely with host genotype, Rhizobium efficiency, soil and climatic conditions and, of course, the methodology used in assessing fixation (Montañez, 2000). The effectiveness of various legume species and their micro-symbionts has been provided in several publications. As a result, selection of suitable legume genotypes that has an ability to form symbiotic relationships with rhizobia is very important for major legumes that are produced in western Ethiopia
CONCLUSION
Nowadays, a diversity of legumes including soybean, common bean, groundnut, pigeon pea and cowpea are the major food legume crops that have been produced in sub-humid tropics of western Ethiopia. Biological nitrogen fixation is an important aspect of sustainable and environmentally friendly food production and long-term crop productivity. However, if BNF is to be utilized, it must be optimized. This review focused on the stress factors that limit biological BNF in leguminous crops. Environmental factors affecting nitrogen fixation include temperature, moisture, acidity and several chemical components of the soil such as nitrogen, phosphorus, calcium, molybdenum content and agronomic management practices. In addition to environmental factors, the competitiveness of the rhizobia in forming nodules and the effectiveness of the Rhizobium-host plant to fix N2 exert a control on N2, fixation. It is often difficult to isolate the effect of the aforementioned factors on inoculation success from their influence on symbiosis and nitrogen fixation. In the near future, particularly in developing countries, tremendous opportunities exist for enhancing the BNF capacity of legumes. Therefore, enhancing BNF by optimizing biotic and abiotic factors that limit BNF is the best option in promoting legume yield in highly depleted soils of western Ethiopia.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Abdenna D, Negassa C, Tilahun G (2007). Inventory of Soil Acidity Status in Crop Lands of Central and Western Ethiopia. "Utilisation of diversity in land use systems: Sustainable and organic approaches to meet human needs" |
|
|
Abdul-Jabbar BKA, Saud HM (2012). Effects of Phosphorus on biological nitrogen fixation in soybean under irrigation using saline water. Global Journal of Science Frontier Research Agriculture and Biology, 12(1):65-72. |
|
|
Abebe Z (2017). On-farm Yield variability and Responses of Common bean (Phaseolus vulgaris L.) Varieties to Rhizobium Inoculation with Inorganic Fertilizer Rates. Journal of Animal and Plant Sciences, 32(2):5120-5133. |
|
|
Abera Y, Assefa F, Moses T, Masso C (2016). Preliminary Characterization of Soybean Nodulating Rhizobia in Ethiopian soils. View. |
|
|
Abubakari F, Tetteh FM, Abubakari F, Tuffour HO, Aduwu A (2016). Strategies for Improving Nodulation and Nitrogen Fixation of Leguminous Crops to Enhance Production in Smallholder Farming Systems. Journal of Global Agriculture and Ecology, 4(4):185-190. |
|
|
Adamovich A, Klasens V (2001). Symbiotically fixed nitrogen in forage legume–grass mixture. Grassland Science in Europe, 6:12. |
|
|
Ahmad I, Akhtar MJ, Asghar HN, Khalid M (2013). Influence of Rhizobium Applied in Combination with Micronutrients on Mungbean. Pakistan Journal of Life and Social Sciences, 11(1):53-59. |
|
|
Alam F, Kim TY, Kim SY, Alam SS, Pramanik P, Kim PJ, Lee YB (2015). Effect of molybdenum on nodulation, plant yield and nitrogen uptake in hairy vetch (Vicia villosa Roth). Soil Science and Plant Nutrition, 61(4):664-675. |
|
|
Al-Falih AMK (2002). Factors affecting the efficiency of symbiotic nitrogen fixation by Rhizobium. Pakistan Journal of Biological Sciences, 5(11):1277-1293. |
|
|
Araujo RS, Cruz SP, Souchie EL, Martin TN, Nakatani AS, Nogueira MA, Hungria M (2017). Preinoculation of Soybean Seeds Treated with Agrichemicals up to 30 Days before Sowing: Technological Innovation for Large-Scale Agriculture. International Journal of Microbiology, 2017. |
|
|
Bambara S, Ndakidemi P A (2010). Phaseolus vulgaris response to Rhizobium inoculation, lime and molybdenum in selected low pH soil in Western Cape, South Africa. African Journal of Agricultural Research, 5(14):1804-1811. |
|
|
Bekere W, Kebede T, Dawud J (2013). Growth and Nodulation Response of Soybean (Glycine max L.) to Lime, Bradyrhizobium japonicum and Nitrogen Fertilizer in Acid Soil at Melko, South Western Ethiopia. International Journal of Soil Science, 8(1):25-31. |
|
|
Bordeleau LM, Prevost D (1994). Nodulation and nitrogen fixation in extreme environments. In Symbiotic Nitrogen Fixation. Springer Netherlands. pp. 115-125. |
|
|
Brear EM, Day DA, Smith PM (2013). Iron: an essential micronutrient for the legume-rhizobium symbiosis. Frontiers in Plant Science, P. 4. |
|
|
Bruijn FJ (2016). Biological Nitrogen Fixation Book Summary. Advances in Microbiology, 6:407-411. http://dx.doi.org/10.4236/aim.2016.66040 |
|
|
Chatterjee R, Bandyopadhyay S (2015). Effect of boron, molybdenum and biofertilizers on growth and yield of cowpea (Vigna unguiculata L. Walp.) in acid soil of eastern Himalayan region. Journal of the Saudi Society of Agricultural Sciences, 21(3):112-117. |
|
|
Coskan A, Dogan K (2011). Symbiotic Nitrogen Fixation in Soybean. INTECH Open Access Publisher. |
|
|
Crawford JTW, Singh U, Breman H (2008). Solving problems related to soil acidity in Central Africa's Geat Lakes Region. International Center for Soil Fertility and Agricultural Development (IFDC) - USA. |
|
|
Danso SKA (1992). Biological Nitrogen fixation in tropical agrosystems: Twenty years of biological nitrogen fixation research in Africa. In Biological nitrogen fixation and sustainability of tropical agriculture: proceeding of the 4th International Conference of the African Association for Biological Nitrogen Fixation, held at the International Institute of Tropical Agriculture, Nigeria, 24-28 Sept 1990. Chichester. View. |
|
|
Food and Agriculture Organization (FAO) (2006). Guidelines for soil description (4th Ed.). Food and Agriculture Organization of the United Nations. Rome. www.fao.org/3/a-a0541e.pdf. |
|
|
Ferreira TC, Aguilar JV, Souza LA, Justino GC, Aguiar LF, Camargos LS (2016). pH effects on nodulation and biological nitrogen fixation in Calopogonium mucunoides. Brazilian Journal of Botany, 39(4): 1015-1020. |
|
|
Foy CD (1984). Physiological effects of hydrogen, aluminium, and manganese toxicities in acid soil. In: Soil acidity and liming, 2nd edition, F. Adams, Ed., Madison, WI: ASA, CSSSA and SSSA. pp. 57-97. |
|
|
Giller KE (2001). Nitrogen fixation in tropical cropping systems. Wallingford, United Kingdom, CAB International. 423 pp. |
|
|
Goi SR, Sprent JI, James EK, Jacob-Neto J (1993). Influence of nitrogen form and concentration on the nitrogen fixation of Acacia auriculiformis. Symbiosis, 14:115-122. |
|
|
Hansen AP (1994). Symbiotic N fixation of crop legumes: Achievements and perspectives. Centre for Agriculture in the Tropics and Subtropics, University of Hohenheim, Germany. MargrafVerlag, Weikersheim, Germany. View. |
|
|
Herridge DF, Roughley RJ, Brockwell J (1984). Effects of rhizobium and soil nitrate on establishment and functioning of the soybean symbiosis in the field. Australian Journal of Agricultural Research 35:146-161. |
|
|
Hirpa L, Niguse D, Setegn G, Geremew B, Firew M (2013). Response to Soil Acidity of Common Bean Genotypes (Phaseolus vulgaris L.) Under Field Conditions at Nedjo, Western Ethiopia. STAR Journal, 2(3):03-15. |
|
|
Hungria M, Franco AA (1993). Effects of high temperature on nodulation and nitrogen fixation by Phaseolus vulgaris L. Plant and Soil, 149(1):95-102. |
|
|
Hungria M, Vargas MA (2000). Environmental factors affecting N2 fixation in grain legumes in the tropics, with an emphasis on Brazil. Field Crops Research, 65(2):151-164. |
|
|
Jones RA, Giddens JE (1985). Tolerance of soybean rhizobia to soil acidity. Pertanika, 8(3):311-315. |
|
|
Kanyanjua SM, Ireri L, Wambua S, Nandw SM (2002). Acidic soils in Kenya: Constraints and remedial options. KARI Technical Note No. 11. Nairobi, Kenya. |
|
|
Keyser HH, Li F (1992). Potential for increasing biological nitrogen fixation in soybean. Plant and Soil, 141(1):119 -135. |
|
|
Lavres J, Castro F G, Sousa CGM (2016). Soybean seed treatment with nickel improves biological nitrogen fixation and urease activity. Frontiers in Environmental Science, 4:37. |
|
|
Mahamood J, Abayomi YA, Aduloju MO (2009). Comparative growth and grain yield responses of soybean genotypes to phosphorous fertilizer application. African Journal of Biotechnology 8(6):1030-1036. |
|
|
Makoi JHJR, Bambara S, Ndakidemi PA (2013). Rhizobium inoculation and the supply of molybdenum and lime affect the uptake of macroelements in common bean (P. vulgaris L.) plants. Australian Journal of Crop Science, 7(6):784-793. |
|
|
Marschner H (2011). Mineral Nutrition of Higher Plants, Academic press, London. P. 672. |
|
|
Masarirambi MT, Mbokazi BM, Wahome PK, Oseni TO (2012). Effects of kraal manure, chicken manure and inorganic fertilizer on growth and yield of lettuce (Lactuca sativa L. var Commander) in a semi-arid Environment. Asian Journal of Agricultural Sciences 4(1):58-64. |
|
|
Massawe PI, Mtei KM, Munishi LK, Ndakidemi PA (2016). Effect of Rhizobium and Intercropping Systems on Soil Nutrients and Biological Nitrogen Fixation as Influenced by Legumes (Phaseolus vulgaris and Dolichos lablab). International Journal of Current Microbiology and Applied Sciences, 5(10):135-149. |
|
|
Mesfin K, Belay Y, Abera H (2014a). Effects of Liming and Phosphorus Levels on Yield and Yield Components of Haricot Bean (Phaseolus vulgaris L.) Varieties on Nitosols of at Wolaita Zone, Ethiopia. Asian Journal of Crop Science, 6(3):245-253. |
|
|
Mesfin K, Belay Y, Abera H (2014b). Liming Effects on Yield and Yield Components of Haricot Bean (Phaseolus vulgaris L.) Varieties Grown in Acidic Soil at Damot Sore District, Southern Ethiopia. International Journal of Soil Science, 2(4):76-81. |
|
|
Mhango WG, Snapp S, Kanyama-Phiri GY (2017). Biological nitrogen fixation and yield of pigeonpea and groundnut: Quantifying response on smallholder farms in northern Malawi. African Journal of Agricultural Research, 12(16):1385-1394. |
|
|
Mmbaga GW, Mtei KM, Ndakidemi PA (2014). Extrapolations on the use of rhizobium inoculants supplemented with phosphorus (P) and potassium (K) on growth and nutrition of legumes. Agricultural Sciences, 5(12):1207. |
|
|
Mohammadi K, Sohrabi Y, Heidar G, Khalesro S, Majidi M (2012). Effective factors on biological nitrogen fixation. African Journal of Agricultural Research, 7(12):1782-1788. |
|
|
Monta-ez A (2000). Overview and case studies on biological nitrogen fixation: Perspectives and limitations. FAO, Case Study, 2:1-11. |
|
|
Munns DN (1986). Acid soils tolerance in legumes and rhizobia. Advances in Plant Nutrition, 2:63-91. |
|
|
Ndakidemi PA, Dakora FD, Nkonya EM, Ringo D, Mansoor H (2006). Yield and economic benefits of common bean (Phaseolus vulgaris) and soybean (Glycine max L) inoculation in northern Tanzania. Australian Journal of Experimental Agriculture, 46(4):571-577. |
|
|
Niste M, Vidican R, Pop R, Rotar I (2013). Stress factors affecting symbiosis activity and nitrogen fixation by Rhizobium cultured in vitro. Pro-Environment/ProMediu, 6(13):42-45. |
|
|
Noor SS, Hossain MA (2007). Effects of boron and molybdenum on the yield of chickpea. Journal of Agriculture and Rural Development, 5(1,2):17-24. |
|
|
Nyoki D, Ndakidemi PA (2016). Intercropping System, Rhizobia Inoculation, Phosphorus and Potassium Fertilization: A Strategy of Soil Replenishment for Improved Crop Yield. International Journal of Current Microbiology and Applied Sciences, 5(10):504-522. |
|
|
Panchali K (2011). Symbiotic nitrogen fixation and seed development of genetically modified soybean in relation to Bradyrhizobium inoculation and nitrogen use under acidic and saline dykeland and soil conditions. MSc thesis, Dalhousie University, Halifax, Nova Scotia. |
|
|
Peoples MB, Ladha JK, Herridge DF (1995). Enhancing legume N2 fixation through plant and soil management. In Management of Biological Nitrogen Fixation for the Development of More Productive and Sustainable Agricultural Systems. Springer Netherlands. pp. 83-101. |
|
|
Rahman MM, Yamamoto M, Niimi M, Kawamura O (2008). Effect of nitrogen fertilization on oxalate content in Rhodesgrass, Guineagrass and Sudangrass. Asian-Australasian Journal of Animal Sciences, 21(2):214. |
|
|
Ramos MLG, Parsns R, Sprent JI, James EK (2003). Effect of water stress on nitrogen fixation and nodule structure of common bean. Pesquisa Agropecuiria Brasileira 38(3):339-347. |
|
|
Rampim L, Lana MDC, Lima PR, Rosset JS, Klein J, Richart A, Guimarães VF, Sarto MVM, Frandoloso JF, Danielle MD, Molin PD, Koppo J, Diel PS, Souza AKP (2015). Inoculation of Bradyrhizobium with cellular additives and micronutrients in soybean seeds cultivated in Oxisol under no-tillage system. African Journal of Microbiology Research, 9(24):1613-1621. |
|
|
Ronner E, Franke AC, Vanlauwe B, Dianda M, Edeh E, Ukem B, Bala A, Van Heerwaarden J, Giller KE (2016). Understanding variability in soybean yield and response to P-fertilizer and Rhizobium inoculants on farmers' fields in northern Nigeria. Field Crops Research, 186:133-145. |
|
|
Saha B, Saha S, Das A, Bhattacharyya PK, Basak N, Sinha AK, Poddar P (2017). Biological nitrogen fixation for sustainable agriculture. In Agriculturally Important Microbes for Sustainable Agriculture (81-128). Springer, Singapore. |
|
|
Saturno DF, Cerezini P, Silva PM, Oliveira AB, Hungria M, Nogueira MA (2017). Mineral nitrogen impairs the biological nitrogen fixation in soybean of determinate and indeterminate growth types. Journal of Plant Nutrition, 40(12):1690-1701. |
|
|
Sogut T, Ozturk F, Temiz MG, Toncer O, Onat BZ (2013). The effects of rhizobial inoculation and nitrogen fertilizer application nodulation, yield and yield components of groundnut (Arachis hypogaea L.). Global Journal on Advances in Pure and Applied Sciences, 01:158-167. |
|
|
Solomon T, Pant LM, Angaw T (2012). Effects of Inoculation by Bradyrhizobium japonicum Strains on Nodulation, Nitrogen Fixation and Yield of Soybean (Glycine max L. Merill) Varieties on Nitisols of Bako, Western Ethiopia. International Scholarly Research Notices, 12:1-8. |
|
|
Tabatabai MA (1986). Sulfur in Agriculture. American Society of Agronomy: Madison, Wisconsin. View. |
|
|
Triplett EW (1990). Construction of a symbiotically effective strain of Rhizobium leguminosarum bv. trifolii with increased nodulation competitiveness. Applied and Environmental Microbiology, 56(1):98-103. |
|
|
TSO (2010). Fertilizer manual (8th Ed.). Department for Environment, Food and Rural Affairs. United Kingdom (UK). |
|
|
Uchida RS (2000). Essential nutrients for plant growth: nutrient functions and deficiency symptoms. In: Silva JA Uchida RS (Eds.), Plant nutrient management in Hawaii soils. Manoa College of Tropical Agriculture and Human Resources, University of Hawaii at Manoa pp. 31-55. |
|
|
Weisany W, Raei Y, Allahverdipoor KH (2013). Role of some of mineral nutrients in biological nitrogen fixation. Bulletin of Environment, Pharmacology and Life Sciences, 2(4):77-84. |
|
|
Workneh B (2013). Liming effects on yield and yield attributes of nitrogen fertilized and bradyrhizobia inoculated soybean (Glycine max L.) grown in acidic soil at Jimma, South Western Ethiopia. Journal of Biology, Agriculture and Healthcare, 3(7):139-143. |
|
|
Wilcox JR, Shibles RM (2001). Interrelationships among seed quality attributes in Soybean. Crop Sciences 41(1):11-14. |
|
|
World Bank (2014). Raise agricultural productivity. View. |
|
|
Zahran HH (1999). Rhizobium-legume symbiosis and nitrogen fixation under severe conditions and in an arid climate. Microbiology and Molecular Biology Reviews, 63(4):968-989. |
|
|
Zerihun A, Alemayo D, Wolde-Meskel E (2015). On farm yield responses of soybean [Glycine max L. (Merrill)] to fertilizer sources under different soil acidity status in Gobu Sayo District, Western Ethiopia. Journal of Agronomy 14(1):30-36. |
|
|
Zerihun A, Tolera A (2014). Yield response of Faba bean to fertilizer rate, Rhizobium inoculation and lime rate at Gedo Highland, Western Ethiopia. Global Journal of Crop, Soil Science and Plant Breeding, 2(1):134-139. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


