Full Length Research Paper

ABSTRACT
The need of characterization of local ecotypes has emerged because of abandonment of traditional varieties and a loss of genetic resources, associated with increasing demography and environmental. Thus, a study was conducted at the Institute of Agricultural Research for Development (IRAD) of Njombé multipurpose station with the objective of characterizing ten local accessions of cassava both morphologically and qualitatively. The work was carried out on two plots following the device Block Full Randomized with 3 repetitions. The yield parameters of leaves, stems, tuberous roots and dry matter of cassava were analysed using analysis of variance (ANOVA) and Duncan test mean separation at 5% using R 3.3.1 software. The evaluation of the cyanogenic potential was made following the method of Picrate Test. During the process of processing into cassava chips, tuberous roots were peeled, washed, cut with a knife, dried in the sun for three days, then packaged in plastic bags and stored as chips. As a result of this analysis, there was a highly significant difference (P Ë‚ 5%) in accessions for yield parameters in leaves, stems, tuberous roots and dry matter content. Accession 68 had an excellent yield of leaves (4.08 t/ha), stems (16.91 t/ha), and tuberous roots (8.95 t/ha). The accession 66 had excellent matter content (40%). From the morphological characterization, it appears that the accessions are all different, even if similarities were observed. The evaluation of the cyanogenic potential revealed that the accessions were low in cyanide content. In view of the conversion into cassava chips, accession 46 with the high percentage (39.20%) of obtaining the chips would be the one to be popularized for the best yield in chips.
Key words: Characterization, morphology, quality, processing, cassava accession, chips.
INTRODUCTION
Cassava (Manihot esculenta Crantz) is a semi-perennial starchy tuber crop belonging to the Euphorbiaceae family (Ballot et al., 2016; Gmakouba et al., 2018). According to FAO's statistical division, cassava feeds nearly 700 million people worldwide and ranks fifth worldwide after wheat, rice, maize and potatoes (Amani et al., 2007; Von Grebmer et al., 2013). In Africa, in 2010, cassava production was estimated at 134 million tons, making it the continent's leading food resource (Von Grebmer et al., 2013). Cameroon ranks 17th in the world with an estimated production of 3,808,239 tons / year of cassava (Anonymous, 2012). Cassava is grown in four of Cameroon's five agro-ecological zones, on about 2 05 000 ha and is one of the most important food crops (Temegne et al., 2015). Despite the importance of cassava in Cameroon, domestic production remains low, and was estimated at 45 96 383 tons in 2013 and the total world cassava utilization is projected to reach 275 million tons in 2020 (Westby, 2008; Njukwe et al., 2014; Anonymous, 2015). This production is lower than that observed in Democratic Congo and Ghana, which are respectively the 2nd and 3rd African producers. Cassava is used for dominant or secondary self-consumption (welding food). It is marketed fresh or after several stages of processing into many products for human food (gari, attiéké, foufou) or animal food (chips, flour) is a source of income for farmers. In the industry, it is used for the manufacture of products such as starch, glues and dextrins (Kouadio et al., 2014).
Cassava cultivation is threatened by pests, weeds and edaphic agronomic and socio-economic factors which are responsible for the drop in yields at 6.4 tons per hectare compared to a world average of 8.8 tons per hectare. This results in the neglecting of local accessions in favor of improved accessions, leading to the erosion of local cassava genetic resources. However, safeguarding these local accessions is a major challenge for sustainable agricultural development (Djaha et al., 2017). Referring to these constraints related to cassava production (low productivity of local varieties), a good knowledge of the morpho-qualitative characteristics and processing skills of a variety can facilitate the choice of an accession to put in place, taking into account the cultural constraints it imposes and the requirements and objectives (quality and quantity of the leaves, stems and roots, etc ...) of the producer. In a context where climate change is leading to more frequent droughts and increasing population pressures to cultivate less and less fertile soils, the particular agronomic benefits of cassava make it a very important plant (McKey et al., 2012). Even though cassava is one of the least demanding plants and can tolerate marginal ecological conditions for agriculture, it is its management by farmers that depends on the sustainability of its culture. This management must take into account the specificities of cassava, which are related on the one hand to its particular adaptation, and on the other hand to the evolutionary constraints that cassava shares with other plants propagated by farmers (Hauser et al., 2014).
In addition to these constraints in production areas, varieties of cassava are designated by vernacular names related to the phenotype or the name of the place or provenance or in the name of the person who introduced it into a locality giving rise to confusion, since the same variety may have different names depending on the area of production (Djaha et al., 2017). In the course of domestication, these confusions have led the Amazonian Indians to practice skilful cassava management, but often misunderstood by scientists, hence the importance of characterizing local accessions of cassava in Cameroon.
The main objective of this work was to characterize ten accessions of cassava of the IRAD of Njombé, to identify those which have the interesting characteristics in relation to the parameters studied in order to popularize them with researchers and farmers.
MATERIALS AND METHODS
Study area
The study was conducted in the locality of Njombé, Njombé-Penja District, Moungo Department, Cameroon Littoral Region (Moist Rainforest Zone with Mono Modal Rainfall) (Figure 1).
The study was conducted on an Andosol, a generally black soil, with a clay fraction dominated by amorphous material and arable layers containing abundant organic matter. It is very porous and is an excellent rooting medium. It has a high phosphate absorption capacity and a strong pH-dependent cation exchange capacity (Beernaert and Bitondo, 1993).
Raw materials
The plant materials consisted of ten local cassava accessions from the IRAD Njombé (A68, A22, A66, A98, A86, A30, A05, A49, A46, A02), with the collections made from different localities of the national territory.
Experimental design
The first plot on which there was characterization and harvesting operations (leaves, stems and tuberous roots) of the ten cassava accessions and was set up in 2015 by IRAD Njombé, following the logic of the bloc device, full randomized 3 repetitions (FRR). The second plot using the same logic, covering a total area of 308 m2, was for morphological characterization of the aerial part of cassava at the physiological stage two to three months after planting.
Collection of data
Morphological characters
The morphological characterization of the ten cassava accessions was carried out on the physico-morphological observation of the characters specific to each accession at different physiological stage (2 , 3 months, 2 years).
Morphological characters were determined through a cassava observation chart (Fukuda et al., 1998). It was the pubescence of the apical leaves, color of the apical leaves, shape of the leaves, color of the leaves, color of the petiole, color of the bark of the stem, the color of the epidermis of the stem, outer color of stem, type of branching, outer color of roots, color of cortex, color of pulp, texture of epidermis, root shape and length of root peduncles.
Performance evaluation
The assessment of fresh yield (leaves, stems, tuberous roots) and dry matter (tuberous roots) of ten local cassava accessions was performed on the first plot. It was done on 3 plants per experimental unit per accession, for a total of 9 plants per accession harvested.
In the field
The data (weight) of the different parts (leaves, stems, tuberous roots) were taken using a precision balance. The number of tuberous roots per plant was done by simple counting.
Laboratory procedures
Dry matter content (DMC)
This invoiced the evaluation of dry matter yield of cassava roots. Two to 3 tuberous roots per plant per accession were taken, peeled, cut into strips of 3 to 4 mm, weighed to a constant weight of 100 g and dried at 105°C for 24 h in an oven. The different samples were weighed using a precision balance. The dry matter content (DMC) was determined using the formula: DMC = (DW / FW) × 100 with DW: dry weight, FW: fresh weight.
Estimation of the cyanogenic potential
The tuberous root samples from the ten cassava accessions harvested were all estimated following the process for the determination of hydrocyanic acid, the Picrate Test (CNP Measurement by Picrate Method).
Operating mode
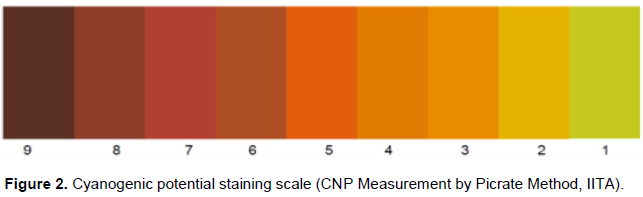
On the tuberous roots collected, a cross section in their middle was carried out. After this cut, cubes of 1 cm3 between the center and the bark of the root were obtained and inserted into test tubes. Five drops of toluene were introduced before closing the test tube. Filter paper (blotting paper) was soaked with an alkaline solution of sodium picrate and then suspended in a tightly closed test tube. After 10 to 12 h, the intensity of the staining was determined using a color scale (Figure 2).
Transformation of tuberous cassava roots into chips
The method used was that developed by IITA (Mück et al., 2000). The tuberous roots were peeled, washed, cut with a knife, dried in the sun for three days, packaged in plastic bags and stored as chips. The yield of cassava chips was presented in terms of rate of chips (RC) according to the formula below:
RC = (DW / FW) X100 with DW: dry weight of the chips; FW: fresh weight of the chips
Data analysis
The data collected were entered into Microsoft Office Excel 2013 spreadsheet. Only fresh yield data (leaves, stems, tuberous roots) and dry matter content (tuberous roots) were analyzed for variance (ANOVA 1), and a separation of means by the Duncan's multiple range test (DMRT) at the 5% level of significance using statistical analysis software R 3.3.1 was done.
RESULTS
Morphological characteristics of each accession
Morphological parameters relating to the aerial part of cassava
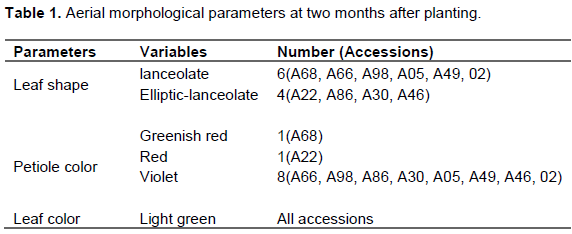
At two months after planting on the second plot, three morphological parameters of the aerial part of cassava were used. During the characterization it was observed that for the parameter forms of leaves, six accessions were lanceolate (A68, A66, A98, A05, A49, A02) and four elliptical-lanceolate (A22, A86, A30, A46). For the color of the petiole one accession was greenish red (A68), another red (A22) and eight others purple (A66, A98, A86, A30, A05, A49, A46, A02). For the leaf color parameter, only one variable was observed as light green (Table 1).
Three months after planting on the second plot
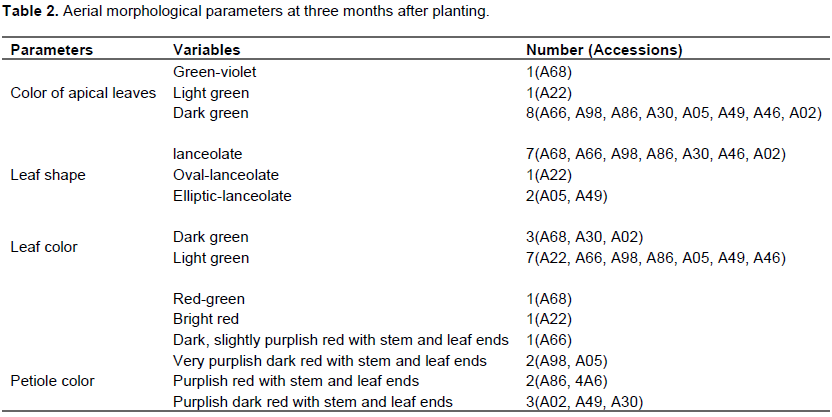
Three months after planting on the second plot, five parameters were used. Absence of pubescence was observed in all accessions. However, for parameters such as apical leaf color, eight accessions were dark green (A66, A98, A86, A30, A05, A49, A46, A02), one accession was purplish green (A68), and another green lightning (A22). For the leaf shape seven accessions were lanceolate (A68, A66, A98, A86, A30, A46, A02), one oval-lanceolate (A22) and two elliptical-lanceolate (A05, A49). For leaf color, two variables were observed, dark green in three accessions (A68, A30, A02) and light green in seven accessions (A22, A66, A98, A86, A05, A49, A46). Regarding the petiole color parameter, a color variation in the red domain was observed. Thus, a red-greenish accession (A68), a bright red (A22), a dark, slightly purplish red at the stem and leaf ends (A66), two very purplish-dark red at the stem and leaf ends (A98, A05), two purplish red at the stem and leaf ends (86, 46) and three purplish-dark red at the stem and leaf ends (A02, A49, A30); (Table 2).
Two years after planting on the first plot
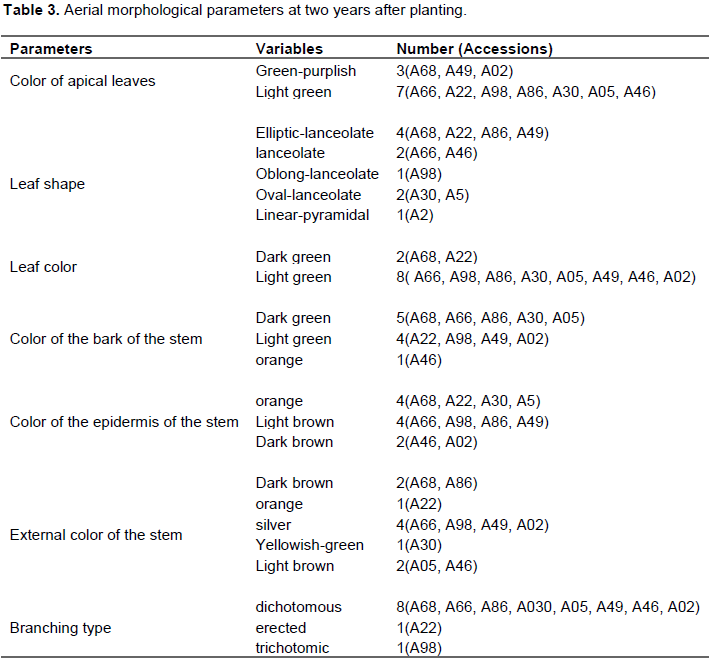
Seven morphological parameters of the aerial parts were used to observe the variations (characters) within ten local accessions of cassava at two years after planting. During this characterization, two variables were observed for the apical leaf color parameter, including the purplish green found in three accessions (A69, A49, A02) and the light green in seven accessions (A66, A22, A98, A86, A30, A05, A46); leaf shape with five variables including elliptical-lanceolate observed in four accessions (A68, A22, A86, A49), lanceolate in two accessions (A66, A46), oblong-lanceolate in one accession (A98), oval-lanceolate in two accessions (A30, A05) and linear-pyramidal in an accession (A02). For the color of the bark of the stem, they were three variables, the dark green of which is observed in five accessions (A68, A66, A86, A30 and A05), the light green observed in four accessions (A22, A98, A49, A02) and orange in an accession (A46). With regards to the type of branching, they were three variables, the dichotomous type observed in eight accessions (A68, A66, A86, A30, A05, A49, A46, A02), accession (A22) and trichotomous in an accession (A98) (Table 3).
Morphological parameters relating to the underground part of cassava
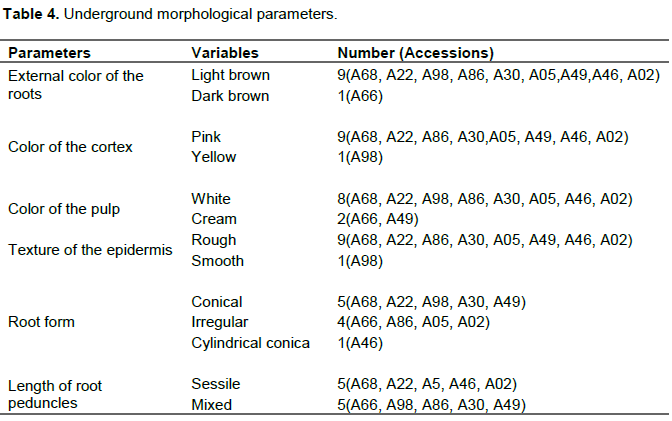
Six parameters were used to characterize the underground part of the ten cassava accessions. This characterization revealed that for the external color parameter of the roots, nine accessions were light brown (A68, A22, A98, A86, A30, A05,.A49,A46, A02) and one dark brown (A66); for the color of the cortex, nine accessions were pink (A68, A22, A86, A30, A05, A49, A46, A02) and one yellow (A98); for the color of the pulp, eight were white (A68, A22, A98, A86, A30, A05, A46, A02) and two cream (A66, A49); for the texture of the epidermis, nine were rough (A68, A22, A86, A30, A05, A49, A46, A02) and one smooth (A98); root shape conical for five (A68, A22, A98, A30, A49), four had irregular shape (A66, A86, A05, A02) and one was cylindrical conical (A46); for the length of the root peduncles, five were sessile (A68, A22, A05, A46, A02) and five mixed (A66, A98, A86, A30, A49) (Table 4).
Evaluation of the performance of each accession
Yield by weight of leaves of each accession
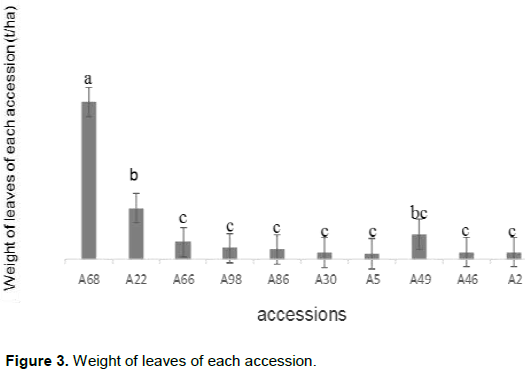
Analysis of the variance showed a highly significant difference in leaf weight per plant (P < 5%). This difference in cassava accessions in terms of leaf weight per plant is illustrated in Figure 3 and showed that accession A68 was different from all accessions while accessions A22 and A49 were not very different but differ significantly from accessions A66, A98, A86, A30, A05, A46 and A02.
Thus, this difference is presented by separating letter accessions (Duncan's multiple range test) where accessions with the same letters are not significantly different.
Weight yield of the stems of each accession
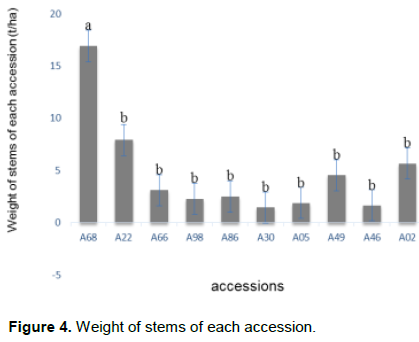
From the analysis of the variance, there is a highly significant difference between accessions in terms of stem weight per plant (P < 5%). This difference is illustrated in Figure 4. Accession A68 had the highest stem weight followed by accessions A22, A02, A49 (which were not significantly different). By concordance, a letter separation of cassava accessions was made using the Duncan's multiple range test.
Fresh tuberous yield of ten accessions
Yield by weight of tuberous roots per plant
Analysis of variance showed that there was a significant difference between the weight of tuberous roots per plant (P < 5%). Figure 5 presents this difference between the accessions. Thus accession A68 had the highest weight followed by accessions A22 and A66.
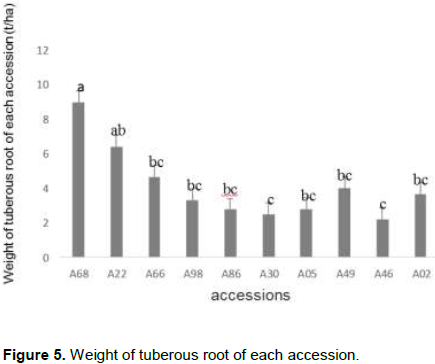
Yield in number of tuberous roots per plant
Analysis of variance of the effect of accessions showed no significant difference in the number of tuberous roots per plant. Accession A66 had the highest mean number of tuberous roots per plant (4.78) followed by accessions A98, A86, A49, A05 and accession A46 with the lowest value (2. 89) (Figure 6).
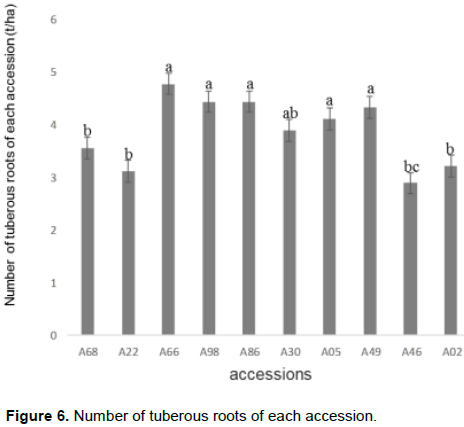
Yield in number of rotten tuberous roots per plant
Analysis of variance showed a highly significant difference in the number of rotten tuberous roots per plant (P < 5%). A separation of these accessions by the Duncan's multiple range test was done where the accessions with the same letters are not significantly different. The highest number of decayed tuberous roots (3.56) was observed in accession A68 and was therefore different from all others. It was followed by accession A49 with an average of 2 rotten tuberous roots per plant and accessions A22, A66, A86; accession A02 had the lowest average (Figure 7).
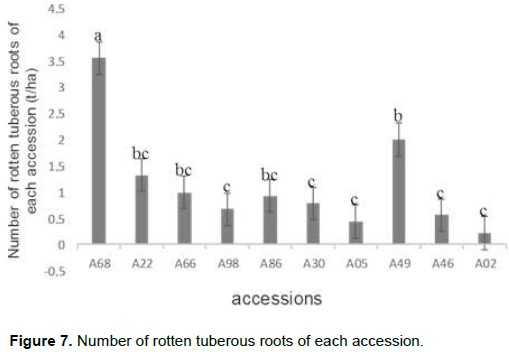
Evaluation of the dry matter content
Analysis of variance revealed a highly significant difference in dry matter per accession (P < 5%). This difference between the accessions is illustrated in Figure 8 where a separation of accessions by the Duncan test was made. Figure 8 also presents the average of each accession in terms of dry matter content and a variation in value between accessions reflecting this highly significant difference. Thus, it was observed that accession A66 had the highest value followed by the accessions A86, A30, A05 and A49. In contrast, accession A68 was the one having the lowest level of dry matter (Figure 8).
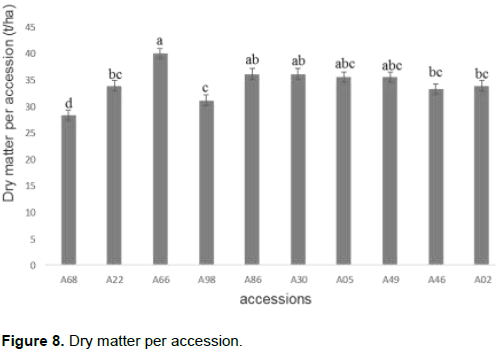
Estimation of the cyanogenic potential of the ten cassava accessions
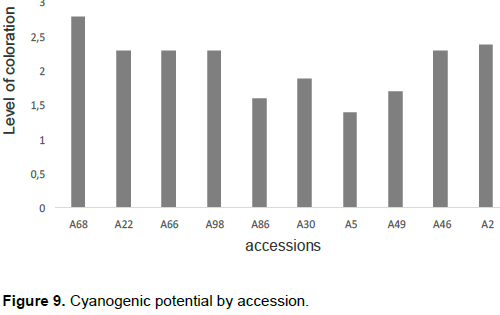
The cyanogenic potential of the accessions was estimated by colorimetric assay, after which a staining threshold was obtained using the Excel 2013 spreadsheet. This accession coloring threshold is presented in Figure 9 where accession A68 had the highest potential followed by accessions A02, A46, A22, A66 and A98 while accession A05 had the weakest potential.
Determination of the yield of chips
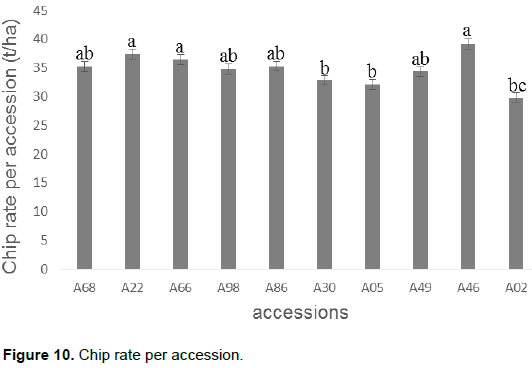
A chip rate was used to determine the chip yield of the ten cassava accessions. In general, all accessions had a chip rate greater than 30%. However, accession A45 had the best rate (39.20%) followed by accessions A22, A66, A86, A68. Accession A02, on the other hand, recorded the lowest rate of smut (29.81%) (Figure 10).
DISCUSSION
Morphological characteristics of each accession
The morphological characters of each accession showed differences amongst the ten accessions of cassava studied, reflecting a great morphological variability within these accessions. These results are similar to those obtained by Djaha et al. (2017) in the same plant. According to these authors, this high variability of accessions in Côte d'Ivoire could be explained by the cultural practice based on the use of several cultivars in the same field; on the other hand, by continuous exchange of planting material with interesting agronomic characters between farmers of different localities. Nevertheless, this varietal richness of cassava could also be due to an excess of heterozygotes within the varieties (accessions). Indeed the variability of the characters could be explained by the result of the crossing between two different individuals by a certain number of characters which are re-associated in a descending individual (Kouadio et al., 2014).
Moreover, there were links of similarities between the accessions. High fractions of the accessions in relation to certain traits are observed and could suggest similarities at the genetic and therefore phenotypic level. This was supported by Kouadio et al. (2014), who started that the similarity between the varieties can find an explanation in the existence of genetic proximity group. This proximity shows the links with gene exchange during sexual reproduction. These differences and similarities between accessions could be attributed to the environment that in the current context of climate change would affect the phenotype of plants. This finding was illustrated by the characterization of these accessions at different physiological stages (two months, three months, 2 years) and is reinforced by the results of Kouadio et al. (2014), which stipulate that the physical characters depend on the varieties and ecological conditions. Thus for the same accession one can have different characters depending on the culture area and time (here relative to the seasons).
Otherwise, the characterization of colors will enable researchers and farmers to recognize an accession according to the criteria that one would like to have such as yield tuber, chips, the degree of acidity of tubers and taste for consumption.
Evaluation of the performance of each accession
The evaluation of the yield of leaves, stems, tuberous roots and dry matter of each accession illustrates the variability of the means of accessions by weight of leaves per plant (WLP), weight of stems per plant (WSP), weight of tuberous roots by plant (WTRP), number of tuberous roots per plant (NTRP), number of rotten tuber roots per plant (NRTRP) and dry matter content (DMC). This variation of the averages observed can be explained by a genetic heterogeneity of the ten accessions of cassava. According to Kouadio et al. (2014), physical characteristics such as diameter, length, number of tubers per plant and individual fresh weight of tubers depends on the varieties and ecological conditions.
Accessions A68, A22, A49, A66, A98 had the highest values by weight of the leaves per plant. These values are justified by the fact that they are branched except accession A22 which is erect. According to Bakayoko et al. (2013); cassava varieties that produce a lot of branching can quickly solve the problem of availability of planting material. On the other hand, these high leaf weight values per plant of the accessions would have their capacity to store a maximum of nutritive reserves necessary for tuberization and may be of interest in the localities where they are consumed as a vegetable.
Accessions A68, A22, A02, A49 had the highest stem weights. These accessions can be profitable for the needs of production and multiplication of plant propagation material. Moreover, the fact that they are branched out with the exception of accession A22, corroborates the results obtained by Bakayoko et al. (2013) in the same plant. However, these high stem weight values ​​per plant may also be related to the number of stems per plant, stem length, and the proportion of mature parts of the stem. This could explain the high value of accession A22 and thus induce a character specific to each accession.
Accessions A68, A22, A66, A49 and A02 had the highest yields of tuberous roots per plant. By retrospective leaf weights per plant, it could be seen that only accessions A02 and A98 did not support the relationship between the leaf area index and the tuberous root yield mentioned above. First accession A98 had a high leaf weight but a low tuberous root (3.29) and accession A02, on the other hand had a low value of (0.20) leaf but a high root, which leads us to make several hypotheses about this result. Given the age (24 months) of the harvested plot, we can therefore think of a continuous growth of accessions, that is to say that, the accessions were able to reach their end of cycle of tuberization and without harvesting, the roots got rotten and the plant continued its cycle. This result could explain the poor performance of other accessions. One can still think of a competition and an inhibition of the development of the other over time. Since in the plot inter-accessions intermingled with one another have been encountered either by lodging or by the fact that one of them has a crawling development and over time comes to prevent the development of the neighboring accession. This might have affected the leaf area index and led to low leaf and tuberous root weight. Such could be the case for accessions A86, A30, A05 and A46. For accession 02, the fact that it had a higher tuberous root yield and a lower leaf weight value could be explained by the phenomenon of verse that would reverse accessions on one another and thus causing a decrease in the leaf index of these.
Nevertheless, the difference in yield of tuberous roots had several explanations: according to Bakayoko et al. (2012); by the growth of storage roots and by the number of roots per plant which is in this case mostly between 3 and 4. According to Segnou (2000), the number of tuberous roots at initiation is 3 per plant and remains constant over time for a local clone. This difference in yield is also reflected in the number of rotten tuber roots that: according to Segnou (2002) when the tuberous roots are left standing beyond the harvest period, waterlogging and starch concentration causes bursting in the soil and resulting cracks are gateways to micro-organisms responsible for rot. According to Kouadio et al. (2014), cassava varieties rot little or not at 12 months after planting since they are not related to any physical character of the plant. This is illustrated by the number of rotten tuber roots of accessions with high or low yields (A68, A22, A66, A49 and A86). According to Bakayoko et al. (2012) the harvest period significantly influences average yields. They observed that at 15 months the yield of fresh tuberous roots was 45 t/ha compared to 39 t/ha at 18 months (loss of 6 t/ha in 3 months for improved varieties). This variation in yield may also be due not only to the varieties, the ecological conditions and the leaf area, but also to the low yield of the local varieties because of their difficult adaptation to the climatic changes and the effects of the diseases and pests: "the tolerant local varieties would not resist any more to the same diseases after some time” (Segnou, 2002). Accessions A66, A86, A30, A05 and A49 are those with the highest solid levels. Accession A66 had the highest dry matter (40%) and accession A68 the lowest. Accession A68 had a good yield in both leaves and tuberous roots and with a number of tuberous roots equivalent to the number of rotten tuber roots. This result is consistent with those obtained by Bakayoko et al. (2012) who have shown that clones with very high yields have a low dry matter content and therefore tend to accumulate a lot of water. Apart from this observation, there is also a variation in the dry matter content between the accessions. This difference in dry matter content between accessions can be explained according to Bakayoko et al. (2012) or by the genotypic properties of the varieties. The production of the dry matter follows a similar evolution to that of the leaf area index and the leaf index most favorable to the development of cassava roots should be between 3 and 5 by the distribution of the dry matter between the various organs of the plant, which undergoes many changes during the cycle: the growth and development of cassava can be influenced by several ecological factors. For example, for long days that lead to a significant drop in yields, temperature drops that can significantly delay tuberization and drought that could speed up the decline of the leaf area index. They assert that the foliar production is greatly reduced at the beginning of the dry season, in order to limit transpiration. Raffaillac and Akakpo (1996) showed that the increase in soil moisture is accompanied by a drop in the dry matter content of cassava roots and noted a correlation between soil moisture and dry matter content in the soil tuberous roots. This could also be a cause of variation in the rate of dry matter (rainy season crossed by accessions). This confirms the results of Segnou (2002), which state that the drop in the concentration of dry matter in the tuberous roots at harvest, 12 months after planting, is due to the return of the rainy season (in a wetted soil, the roots are more easily engorged with water). According to this author, the difference in yield of dry matter results from the volume of the leaf mass, the interception of sunlight, the photosynthetic activity and the translocation speed of the nutritive reserves of the leaves to the tuberous roots during the vegetative cycle. However, these statements on the water content of the roots and the environment suggest that the dry season is not as bad as that to obtain a good dry matter yield even if it causes a reduction of the index foliage of the plant since a low water content in the roots leads to a high concentration of dry matter. This corroborates the works of Segnou (2002) who finds that the high concentration in dry matter is related to the dry season which occurred in the experimental locality, which could probably be the case observed in the results obtained since the harvest of these ten accessions occurred between the months of April and May.
Estimation of the cyanogenic potential of the ten cassava accessions
In light of the cyanogenic potential estimate, there was a variation in the threshold of staining for accessions with A68 having the highest staining threshold (2.8) and the lowest in accession (1.4). This could possibly be due to the varieties and would suggest a genetic character specific to each accession. However, this variation in the cyanogenic potential of accessions suggests the influence of certain factors such as the age of the plant (two years) but also ecological conditions that in this current context of climate change greatly influences agriculture and cultural conditions. This is supported by the results of Atibu (2004), who states that the concentration of cyanide varies widely between varieties and ecological and cultural conditions. These results also corroborate the results of Delange et al. (1982) who found that the concentration of hydrocyanic acid in roots varies considerably among the varieties studied and that, in addition to differences between varieties, the hydrocyanic acid content in cassava tissues appears to be affected by several environmental factors such as soils and temperatures. They also specified that this content of hydrocyanic acid in cassava tissues grown under the same identical edaphoclimatic conditions differ, at a given age. Like these authors, Amani et al. (2007) have shown that the content of hydrocyanic acid depends on the climatic conditions of development of the plant. Indeed, in the case of a severe dry season the content of hydrocyanic acid increases. The generalized reading of the results makes it possible to observe a low staining threshold and a small variation of the cyanogenic potential of the accessions. This would allow later to consider a classification of these accessions in varieties with high or low potential cyanogen because according to Delange et al. (1982); cassava cultivars are classified as "sweet" or "bitter" depending on their hydrocyanic acid content in the fresh pulp. In 2004, Atibu presented two bases for classifying cassava varieties based on the content of hydrocyanic acid.
Determination of the yield of chips
The determination of the chip yield of each accession shows a variation in the rate of chips between accessions (29.81 to 39.20%). Accession A46 has the highest value followed by accessions A22, A66, A86 with 37.42, 36.45 and 35% respectively. This difference in the rate of chips between accessions is mainly due to a variation in the moisture content and gene pool of each accession. These results could help the scientist re-orientate the selection program in cassava by crossing accessions according to desired characteristics. These results are in agreement with those obtained by Gbemenou et al. (2011) on the same plant. In addition, the dry matter content varies from one accession to another. This leads us to re-implicate the hypotheses and thoughts about the ability of each accession to store the nutrients from photosynthesis. However, given the age of the plants, the roots have matured for a long time, left in the ground these roots have continued their maturation until the stage of withering characterized by a cracking and an increase of fibers in the tuberous roots causing thus a decline in reserves.
CONCLUSION
From this study, there is clear variability of the morphological and qualitative characters between the accessions. Regarding the morphological characters (above and below ground), all cassava accessions were different from each other, even if they had some similarities. However, accessions with ramifications are presented as a solution in obtaining the planting material. Regarding yield, accession A49 was capable of meeting all expectations and is the only accession with a good yield of leaves, stems, tuberous roots and dry matter. Nevertheless, accession A68 is capable of answering the problem of nutrition of populations with fresh tuberous roots and leaves as a vegetable. For the dry matter content conversion, accession A66 is the most suitable for meeting this requirement, and for the conversion to chips, accession A46 is the most appropriate. Moreover, in view of the evaluation of their cyanogenic potential, all these accessions can be classified as sweet cassava since they all have a potential of less than 3 on the color scale.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Amani G, Nindjin C, N'zue B, Tscchannen A, Aka D (2007). Potentialités à la transformation du manioc (Manihot esculenta Crantz) en Afrique de l'Ouest. Actes de l'atelier internationnal, Abidjan, Côte d'Ivoire 341 p. |
|
|
Anonymous (2012). Annuaire des statistiques du secteur agricole exercice 2009/2010 n°17. Division des études et projets agricoles, Cellule des enquêtes et statistiques. Ministère de l'agriculture. Yaoundé, Cameroun 123 p. |
|
|
Anonymous (2015). Chapiter 14: Agriculture ; Institut National de la Statistique, Annuaire Statistique du Cameroun 24 p. |
|
|
Atibu KE (2004). Cinétique de l'élimination du cyanure dans le manioc. Université de Kinshasa - Licence en Sciences groupe Chimie 36 p. |
|
|
Bakayoko S, Kouadio KKH, Soro D, Tschannen A, Nindjin C, Dao D, Girardin O (2012). Rendements en tubercules frais et teneurs en matière sèche de soixante-dix nouvelles variétés de manioc (Manihot esculenta Crantz) cultivées dans le centre de la Côte d'Ivoire. Journal of Animal and Plant Sciences 14(2):1961-1977. |
|
|
Bakayoko S, Soro D, N'dri B, Kouadio KKH, Tschannen A, Nindjin C, Dao D, Girardin O (2013). Etude de l'architecture végétale de 14 variétés améliorées de manioc (Manihot esculenta Crantz) dans le centre de la Côte d'Ivoire. Journal of Applied Biosciences 61:4471-4477. |
|
|
Ballot C, Wango S, Atakpama W, Semballa S, Zinga I, Batawila K, Akpagana K (2016). Amélioration des rendements de la culture du manioc (Manihot esculenta Crantz, Euphorbiales, Euphorbiaceae) par les terres de termitières dans la zone de savane de Damara en République Centrafricaine. Revue Marocaine des Sciences Agronomiques et Vétérinaires 4(2):40-53. |
|
|
Beernaert F, Bitondo D (1993). Land Evaluation Manual. Department of Soil Science Centre Universitaire de Dschang, Cameroun. Cassava Biotechnology Network (6, 2004, Cali, Colombia) 396 p. |
|
|
Delange F, Ahluwalia R (1982). Cassava toxicity and thyroid. Proceedings of a workshop held in Ottawa, Canada. IDRC. s.l.: OMS 17 p. |
|
|
Djaha EK, Kouabenan A, Bonny BS, Tchoa K, Amouakon WJL, Kone D, Kone M (2017). Caractérisation agro-morphologique de 44 accessions de manioc (Manihot esculenta Crantz) cultivés en Côte d'Ivoire. International Journal of Biological and Chemical Sciences 11(1):174-184. |
|
|
Fukuda WMG, Guevara CL, Kawuki R, Ferguson ME (1998). Selected morphological and agronomic descriptors for the characterization of cassava. IITA 19 p. |
|
|
Gbemenou J, Gnonlonfin B, Koudande DO, Sanni A, Brimer L (2011). Farmers' perceptions on characteristics of cassava (Manihot esculenta Crantz) varieties used for chips production in rural areas in Benin, West Africa. International Journal of Biological and Chemical Sciences 5(3):870-879. |
|
|
Gmakouba T, Koussaos, Traore ER, Kpemoua KE, Zongo JD (2018). Analyse de la diversité agromorphologique d'une collection de manioc (Manihot esculenta Crantz) du Burkina Faso. International Journal of Biological and Chemical Sciences 12(1):402-421. |
|
|
Hauser S, Wairegi L, Asadu CLA, Asawalam DO, Jokthan G, Utiang U (2014). Guide to growing cassava. Africa Soil Health Consortium 48 p. |
|
|
Kouadio KKH, Ettien DJB, Bakayoko S, Soro D, Girardin O (2014). Variabilité physico-morphologique des racines tubéreuses de manioc (Manihot esculenta Crantz) cultivées sur ferralsol en zone de forêt d'Afrique de l'Ouest. Journal of Applied Biosciences 82:7316-7325. |
|
|
McKey D, Elias M, Pujol B, Duputié A, Delêtre M, Renard D (2012). Maintien du potentiel adaptatif chez les plantes domestiquées à propagation clonale. Leçons de gestion par les cultivateurs de manioc amérindiens. Revue d'ethnoécologie 1:1-27. |
|
|
Mück O, Schuler B, Bell A (2000). les richesses du sol, Chapitre 2 : le manioc, un aliment pour tout le monde. Deutsche stiftung für internationale entwicklung 330 p. |
|
|
Njukwe E, Hanna R, Sarr SP, Shigeru A, Kirscht H, Mbairanodji A, Ngue-Bissa T, Tenkouano A (2014). Cassava value chain development through partnership and stakeholders' platform in Cameroon. International Journal of Agricultural Policy and Research 2(11):383-392. |
|
|
Raffaillac JP, Akakpo K (1996). Matière sèche des racines de manioc et aptitude à la transformation en foufou au Togo. Cahiers Agricultures 5(3):185-188. |
|
|
Segnou M (2000). Caractéristiques morpho-physiologiques liées à la précocité du rendement chez le manioc. Cahiers Agricultures 9:77-79. |
|
|
Segnou M (2002). Développement végétatif et potentiel de rendement chez le manioc. Tropicultura 20(4):161-164. |
|
|
Temegne NC, Ngome AF, Kuate AF (2015). Influence de la composition chimique du sol sur la teneur en éléments nutritifs et le rendement du manioc (Manihot esculenta Crantz, Euphorbiaceae) dans deux zones agro-écologiques du Cameroun. International Journal of Biological and Chemical Sciences 9(6):2776-2788. |
|
|
Von Grebmer K, Headey D, Bene C, Haddad L, Olofibiyi T, Wiesmann D, Fritschel H, Yin S, Yohannes Y, Foley C (2013). Indice de la faim dans le monde : Le def de la faim: Construire la résilience pour une sécurité alimentaire et nutritionnelle durable 66 p. |
|
|
Westby A (2008). Cassava Utilization, Storage and Smallscale Processing. In R. Hillock, J. Thresh, & A. C. Bellotti, eds., Cassava Biology, Production and Utilization. CABI Publishing pp. 281-300. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


