Full Length Research Paper

ABSTRACT
Maize is one of the most important cereal crop widely grown in the world. Maize crosses along with similar maturing checks were evaluated at Hawassa in 2015-2016 cropping season to understand the nature of gene action governing yield and its attributes through line × tester analysis and to study genetic contribution of line, tester and the interaction of line × tester to total variation. The experiment was done by using α-lattice design 6 × 11 arrangement. Cross L31×T2 showed the highest mean grain yield than both checks. The result showed that, lines played an important role towards days to anthesis, days to silking, ear length, number of rows per cob and number of kernels per row, indicating predominance of maternal lines. Based on analysis of genetic variance, traits variance due to specific combining ability (σ²SCA) was higher than variance due to general combining ability (σ²GCA) indicated, non-additive gene action was important than additive gene action in the inheritance of these traits. These best cross combinations could be effectively utilized in maize breeding for the improvement of yield components and thus their incorporation in further breeding program is suggested.
Key words: Additive, non-additive, cross, genotypes, line × tester, yield.
INTRODUCTION
Maize is one of the most important cereal crops in the world after wheat and rice. Maize is nutritionally, an important crop used as food and feed. It is a source of industrial materials for the production of fuel, oil, starch, syrup; gluten, alcohol, glucose, ethanol and many more products. Its cultivation extends over a wide range of geographical and environmental conditions ranging from 58°N to 40°S. Portuguese traders introduced maize to Ethiopia in 16th or 17th century (Haffnagel, 1961). Currently, in Ethiopia, maize is one of the most important cereal crops grown in almost all parts of the country. The popularity of maize in Ethiopia is partly because of its high value as a food, fodder and source of fuel for rural area. Approximately, 88% of maize produced in Ethiopia is consumed as food, both as green and dry grain (Abate et al., 2015). The total annual production and productivity of maize in Ethiopia exceeds all other cereal crops except Tef in area coverage (Mosisa et al., 2011).
Objective of maize breeding programs is evaluation of best yielding and adaptive. Improvement of varieties (genotypes) needs deep genetic information. Breeders conducted a genetic analysis for yield and yield related traits of genotypes. In fact, maize has been subjected to extensive genetic studies than any other crop (Hallauer and Miranda, 1988). Several biometrical techniques were used to study genetic analysis of quantitative traits. Among them, line × tester is suggested by Kempthorne (1957) and is used to breed both self and cross pollination plants. This method of efficient study on large number of lines provides reliable estimates of genetic components, estimates of specific combining ability (SCA), general combining ability (GCA) and gene action governing quantitative traits.
Therefore, the current study aimed to understand the nature of gene action governing yield and its attributes through line × tester analysis and to study genetic contribution of line, tester and the interaction of line × tester and total variation.
MATERIALS AND METHODS
Experimental materials and design
Hawassa is situated at 7°4′N and 38°31′E latitude and longitude, respectively, at an altitude of 1700 m.a.s.l. in the central rift valley of Ethiopia. Sixty-four crosses were developed by crossing 32 inbred lines with two testers in line × tester mating design (Kemperton, 1957). Inbred lines (32) were coded as L1, L2…L32 and two testers CML-144 (F2) -21-2-2-1- 1/CML144 and CML-144 (F2) -15-2-2-2-1/CML159 were coded as T1 and T2. Sixty-four crosses were planted along with similarly maturing checks BHQPY-545 and BH-546 at Hawassa. The experiment was planted by using α-lattice design 6×11 arrangement (Patterson and Williams, 1976) with two replications. Each block comprises of 11 units having 5.1 m long and 9.75 m width with the spacing of 0.75 m between rows and 0.30 m between plants. All cultural practices were performed as per required (Table 1a).


Data like days to maturity (DM), field weight, seed moisture content and thousand kernel weight (TKW) were collected plot bases while data like plant height (PH), ear height (EH), ear length (EL), number of rows per ear (NRPE), ear diameter (ED) and number of kernels per row (NCPR) were collected on plant bases. Biomass (BM) and grain yield (GY) was calculated by using the following formula:

Where: MC = moisture content of grain at harvest, 0.8 = shelling percentage, 85 = standard moisture content of grain, n = number of plants harvested, 17 = total number of plants in a plot, 10000 = area of hectare in square meters.
Data analysis
All data obtained were subjected to SAS computer software to test the significance genotypes (Gomez and Gomez, 1984). Genetic parameter analysis and proportional contribution of line tester and line × tester were done only for tr.
Genetic parameter analysis

The ratio of σ²GCA to σ²SCA was expressed as 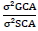
Additive variance
σ²l=[Ms(l)-Mse]/rt=1/2σ²A σ²A=2σ²l
σ²t=[Ms(t)-Mse]/rl=1/2σ²A σ²A=2σ²t
σ²A=[2σ²l+2σ²t]/2=σ²l+2σ²t
σ²D=dominance variance
σ²lt=[Ms(l×t)-Mse]/r= σ²D
Average degree of dominance (Ä) was calculated according to the following equation:

if Ä = 0 no dominance
if Ä = <1> 0 partial dominance
if Ä = 1 complete dominance
if Ä > 1 over dominance
Where: σ²GCA = variance of general combining ability, σ²SCA = variance of specific combining ability, MSl = mean square of line, MSt = mean square of tester, MSl×t = mean square of line × tester, l = line, t = tester, r = replications.
Proportional contribution of line, tester and line × tester to total variation in hybrid combinations
The percentage contribution of lines (females), testers (males) and line × tester to the hybrids were calculated according to Abuali et al. (2016):

Where, SS (c) = sum square of cross, SS (l) = sum square line, SS (t) = sum square tester, SS (L×T) = sum square of line × tester, l = line, t = tester, r = replications.
RESULTS AND DISCUSSION
Genetic parameters analysis
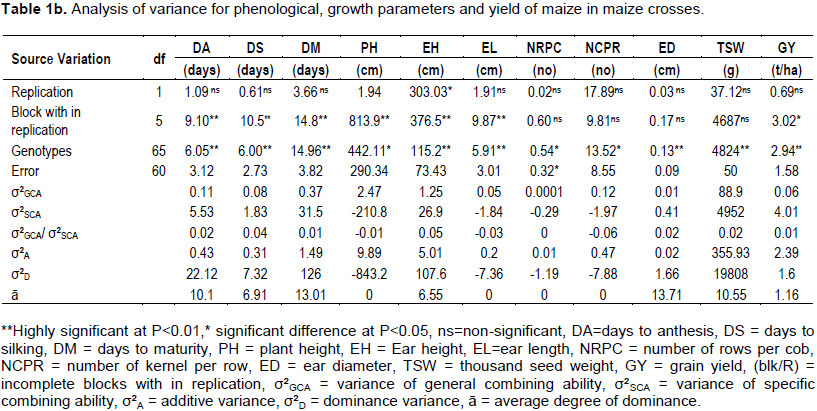
The analysis of variance showed that, there was a significance difference between genotypes for all traits. The analysis of variance indicated that, sufficient genetic variability is present among genotypes for all characters (Table 1b). Variance due to SCA (σ²SCA) was higher than variance due to general combining ability (σ²GCA) and the ratio of σ²GCA to σ²SCA was less than one for traits like days to anthesis, days to silking, days to maturity, ear diameter, thousand kernel weight, grain yield and cob per plant which indicate, non-additive gene action was more important than additive gene action in the inheritance of these traits (Table 1). Non additive gene action is not easily fixable, implies that best hybrids were not easily identified for the following traits. Similarly, Atanaw et al. (2010) reported that, non-additive gene effects were important than additive gene effect for grain yield. Also, Kamara et al. (2014) found similar result for ear diameter and thousand kernel weight. σ²GCA was larger than σ²SCA in plant height, ear length, number of rows per cob, number of kernels per row and biomass which indicates the additive gene action played the great role in governing the inheritance of these traits than non-additive gene action (Table 1). Additive gene action is easily fixable, implying that, best hybrids were easily identified for the following traits. Alamnie et al. (2007) and Panhwar et al. (2008) reported that, additive gene effects were more important than non-additive gene effects for plant height and number of kernels per row. The result was pact with that of Sharma et al. (2004) who found preponderance of additive genetic effects in the control of traits like plant height, ear length, number of rows per cob, number of kernels per row and biomass. Similar result has been reported by different researchers (Irshad-El-Haq et al., 2010; El-Badawy, 2012; Aminu et al., 2014) for grain yield.
The value of additive gene effects was more than the value of dominance gene effect for plant height, ear length, number of kernels per row, number of rows per cob, while the value of dominance gene effects was higher than the value of additive gene effects for days to anthesis, days to silking, days to maturity, ear height, ear diameter, thousand seed weight and grain yield (Table 1). The average degree of dominance was more than one for days to anthesis, days to silking, days to maturity, ear height, ear diameter, thousand seed weight and grain yield, indicating these traits were under control of the over dominance gene effect, whereas the average dominance was zero for traits like plant height, ear length, number of kernels per row, number of row per cob indicate there was no dominance for the traits (Table 1).
Mean performance of genotypes (crosses and check)
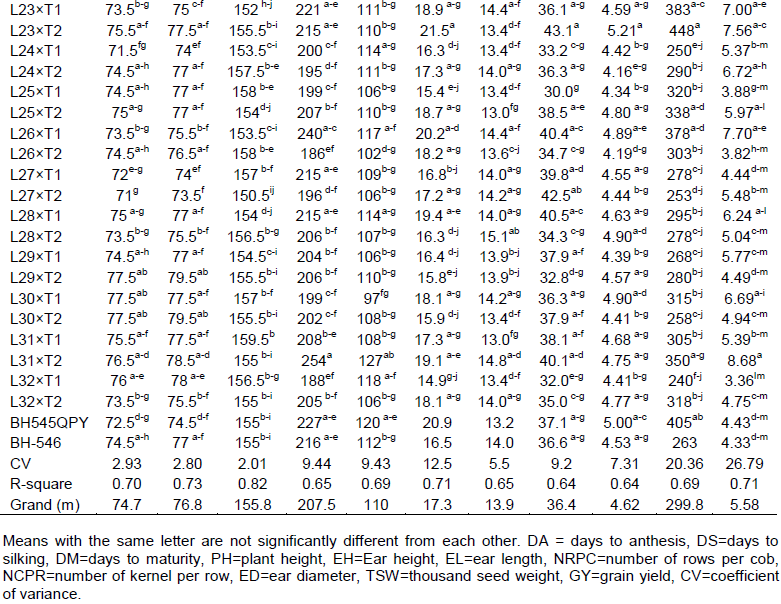
The mean performances of genotypes are presented in Table 2. Top ten high yielding crosses relative to both checks were L31×T2 (8.68 t ha -1), L8×T1 (8.25 t ha -1), L26×T1 (7.70 t ha-1), L23×T2 (7.56 t ha-1), L12×T2 (7.04 t ha-1), L16×T1 (7.00 t ha-1), L23×T1 (7.00 t ha-1), L21×T1 (6.93 t ha-1), L21×T2 (6.91 t ha-1) and L8×T2 (6.82 t ha-1) (Table 2). The mean grain yield of check BHQPY-545 and BH-546 were 4.4 t ha-1 and 4.3 t ha-1, respectively. Low yielding crosses in relation to checks were L2×T2 (4.24 t ha-1), L6×T2 (3.59 t ha-1), L7×T1 (4.37 t ha-1), L9×T2 (4.03 t ha-1), L11×T2 (3.81 t ha-1), L14×T2 (3.67 t ha-1), L17×T2 (2.88 t ha-1), L20×T2 (3.93 t ha-1), L25×T1 (3.88 t ha-1), L26×T2 (3.82 t ha-1) and L32×T1 (3.36 t ha-1) (Table 2). The grain yield ranged from 2.88 t ha-1 (L17×T2) to 8.68 t ha-1 (L31×T2) with an overall mean of 5.55 t ha-1 (Table 1). Those crosses that had better mean values over standard checks indicate the possibility of obtaining promising hybrid variety with desirable traits. Highest mean number of days to maturity was obtained for cross L11×T1 (166 days), while the lowest mean number of days to maturity was recorded for cross L12×T2 (149.5 days). Three crosses L12×T2 (-5.5 days), L21×T2 (-4.5) and L27×T2 (-4.5 days) were early as compared to both standard checks BHQPY-545 and BH-546 (Appendix 2). Plant height for genotypes ranges from 165 (L2×T1) to 254 cm (L31×T2) with the mean 207.5 cm. Ear height for genotypes ranges from 95 (L7×T2) to 134 cm (L19×T2) with the mean of 110 cm (Table 2).

Proportional contribution of line, tester and line × tester
The proportional contribution of lines, testers and the interaction of line ×tester to the total variances are presented in 3. The result showed that, lines played an important role in days to anthesis, days to silking, ear length, number of rows per cob and number of kernels per row, indicating that predominant of maternal (lines) influence these traits and higher estimates of variance due to GCA (Table 3). The contribution of testers was low for all traits, which indicates higher estimates of variances due to SCA. The contribution of line × tester interactions played an important role in days to maturity, plant height, ear height, ear diameter, thousand kernel weight and grain yield, which indicate higher estimates of variances due to non-additive genetic effects and the importance of SCA. Shams et al. (2010) observed higher estimates of SCA variance due to line × tester. Aminu et al. (2014) also found the proportional contribution of line × tester was greater than tester for grain yield, plant height, ear height, thousand kernels weight and ear length of their study on combining ability and heterosis for phenologic and agronomic traits in maize (Zea mays L.) under drought conditions. In contrast, Shams et al. (2010) found proportional contribution of line × tester interaction was greater than line and tester for number of kernels per row and proportional contribution of tester was greater than line and the interaction line × tester in number of rows per cob in maize using line × tester method.

CONCLUSION
The analysis of variance showed sufficient genetic variability among genotypes for all characters. σ²SCA was greater than σ²GCA for traits like days to anthesis, days to silking, days to maturity, ear diameter, thousand kernel weight, grain yield. σ²GCA was larger than σ²SCA in plant height, ear length, number of rows per cob, number of kernels per row and biomass. The proportional contribution of line is greater than tester and the interaction of line × tester for traits like days to anthesis, days to silking, ear length, number of rows per cob and number of kernels per row. The proportional contribution of line × tester is greater than line and tester for days to maturity, plant height, ear height, ear diameter, thousand kernel weight and grain yield.
CONFLICT OF INTERESTS
The author declares that there is no conflict of interest.
ACKNOWLEDGEMENTS
The author thanks Oromia Agriculture Research Institute (OARI) for the funding, and also express gratitude to Yabello Pastoral and Dry Land Agricultural Research Center for facilitating financial assistance during the study. And sincerely appreciate and thank Hawassa Maize Research staffs for their support in planting, trial management and data collection.
REFERENCES
|
Abate T, Shiferaw B, Menkir A, Wegary D, Kebede Y, Tesfaye K, Kassie M, Bogale G, Tadesse B, Keno T (2015). Factors that transformed maize productivity in Ethiopia. Food Security 1; 7(5):965-81. |
|
|
Abuali AI, Khalafalla MM, Abdelmula AA, Idris AE, Osman AM (2016). Combining ability and heterosis for yield and yield components in maize (Zea mays L.). |
|
|
Alamnie A, Wali MC, Salimath PM, Jagadeesha BC (2006). Combining ability and heterosis for grain yield and ear characters in maize. Karantaka Journal of Agricultural Science, 19:13-16. |
|
|
Atanaw A, Nayakar NY, Wali MC (2010). Combining ability, heterosis and per se performance of height characters in maize. Karnataka Journal of Agricultural Science, 29(1):16. |
|
|
Aminu D, Garba M, Muhammad AS (2014). Combining ability and heterosis for phenologic and agronomic traits in maize (Zea mays L.) under drought conditions in the Northern Guinea Savanna of Borno State, Nigeria. African Journal of Biotechnology, 13(24). |
|
|
El-Badawy ME (2012). Estimation of genetic parameters in three maize crosses for yield and its attributes. Asian Journal of Crop Science, 4(4):127. |
|
|
Gomez KA, Gomez AA (1984). Statistical procedures for agricultural research. John Wiley & Sons; Feb 17. |
|
|
Haffnagel HP (1961). Agriculture in Ethiopia. FAO, Rome P 420. |
|
|
Hallauer AR, Miranda Filho JB (1988). Quantitative genetics in maize breeding, 2nd edn (Iowa State University Press: Ames, IA). |
|
|
Irshad-El-Haq M, Ajmal SU, Munir M, Gulfaraz M (2010). Gene action studies of different quantitative traits in maize. Pakistan Journal of Botany, 42(2):1021-30. |
|
|
Kamara MM, El-Degwy IS, Koyama H (2014). Estimation combining ability of some maize inbred lines using line × tester mating design under two nitrogen levels. Australian Journal of Crop Science, 8(9):1336. |
|
|
Kempthorne O (1957). An introduction to genetic statistics. John Wiley And Sons, Inc.; New York. |
|
|
Mosisa W, Legesse W, Berhanu T, Girma D, Girum A, Wende A, Tolera K, Gezahegn B, Dagne W, Solomon A, Habtamu Z (2011). Status and future direction of maize research and production in Ethiopia. In Meeting the Challenges of Global Climate Change and Food Security through Innovative Maize Research, P 17. |
|
|
Panhwar SA, Baloch MJ, Jatoi WA, Veesar NF, Majeedano MS (2008). Combining ability estimates from line × tester mating design in upland cotton. Pakistan Academy of Sciences, 45:69-74. |
|
|
Patterson HD, Williams ER (1976). A new class of resolvable incomplete block designs. Biometrika 1:83-92. |
|
|
Shams MA, Choukan RA, Majidi ES, Darvish FA (2010). Estimation of combining ability and gene action in maize using line× tester method under three irrigation regimes. Journal of Agricultural Science, 6(1):19-28. |
|
|
Sharma S, Narwal, MS, Kumar R. Dass S (2004). Line × Tester analysis in maize. Forage Research, 5(30):28-30. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


