ABSTRACT
Neuropeptides are mediators of neuronal signalling, controlling a wide range of physiological processes that cut across many organisms. GFSKLYFamide is a neuropeptide that belongs to the Echinoderm SALMFamide group of peptides. This study was aimed at determining the expression of GFSKLYFamide-like neuropeptide in the female gonad of the sea cucumber Holothuria scabra. Ten female H. scabra weighing between 62 and 175 g were collected at different times between May, 2009 and April, 2010 from the Andaman Sea at Koh Jum Island in Krabi province of Thailand for the study. Using GFSKLYFamide polyclonal antibody and Alexa 488-conjugated goat anti-rabbit IgG as primary and secondary antibodies, respectively, indirect immunofluorescence method and confocal microscope was used to localise GFSKLYFamide-like immunoreactivity in the female gonad of H. scabra. The results show GFSKLYFamide-like immunoreactivity being expressed in the sub-epithelial fibres of the coelomic epithelium. This is the first report providing evidence of GFSKLYFamide neuropeptide existence in the female gonad of H. scabra. It was suggested that further studies be carried out to investigate if there is similar existence of GFSKLYFamide neuropeptide in male gonad of H. scabra and also to know the structure or activity of the peptide(s) producing GFSKLYFamide-like immunoreactivity in H. scabra gonads.
Key words: GFSKLYFamide, SALMFamides, immunoreactivity, sea cucumber, echinoderm, gonad.
Neuropeptides have been known to play a crucial role in mediating several physiological processes in animals (Ajayi and Withyachumnarnkul, 2015; Semmens and Elphick, 2017). The neuropeptide, GFSKLYFamide (Gly-Phe-Ser-Lys-Leu-Tyr-Phe-NH2) is a heptapeptide first isolated in the sea cucumber Holothuria glaberrima (Diaz-Miranda et al., 1992). It belongs to the Echinoderm SALMFamide group of peptides and like many other
neuropeptides in this family, GFSKYLFamide is believed to play a crucial role in some physiological processes particularly those involving muscular relaxation (Melarange et al., 1999; Elphick and Melarange, 2001).
Echinoderms are of particular interest for studies on neuropeptides because as deuterostomes, they bridge the evolutionary gap between chordates and model protostomian invertebrates and provide a better understanding of the evolution of neuropeptide systems in the animal kingdom as whole (Semmens and Elphick, 2017). The sea cucumber, Holothuria scabra that is known as sandfish belongs to the Echinoderm phylum in the class of Holothuroidea, of the order Aspidochirotida and in the family of Holothuriidae. It is a tropical sea cucumber species that is overexploited due to its commercial value (Conand 1998; Purcell et al., 2012).
Research activities aimed at understanding the neurohormonal regulation of reproduction in this species has been done in our laboratory (Ajayi and Withyachumnarnkul, 2013). Efficient spawning induction in the animals with minimal stress is important in culture; hence, the need to understand the neurohormonal cues regulating gamete release, particularly in the female species, which would provide the needed eggs for fertilization. Moreover, there is no evidence to show that GFSKLYFamide-like neuropeptide, which has been reported in the gonads of H. glaberrima exists in H. scabra. The present study is therefore aimed at determining the expression of GFSKLYFamide-like neuropeptide in the female gonad of the sea cucumber H. scabra.
Animals
Ten female H. scabra weighing between 62 and 175 g were used in this study. The animals were collected at different times between May, 2009 and April, 2010. Samples were collected from the Andaman Sea at Koh Jum Island in Krabi province of Thailand with the following coordinates; 7.8119°N, 98.9770°E. They were maintained in filtered natural sea water within a temperature range of 28 to 31°C and salinity of about 32 ppt at the Shrimp Genetic Improvement Centre, Chiya, Suratthani before being transferred to the laboratory in Bangkok in oxygenated sealed plastic bags over a distance of 530 km by train.
Antibody
Polyclonal antibody against GFSKLYFamide was generously provided by Professor García-Arrarás (University of Puerto Rico, USA). The antibody was raised as described by Díaz-Miranda et al. (1995), using 63 g of synthetic GFSKLYFamide coupled to 15 mg BSA with 0.3% glutaraldehyde. The reaction was stopped by the addition of 1M glycine and the mixture dialyzed. Aliquot of the dialysate BSA-GFSKLYFamide conjugate was emulsified with complete Freund’s adjuvant and injected into two rabbits with half of the emulsion each, subcutaneously and intraperitoneally. Two boosters of the aliquot mixed with incomplete Freund’s adjuvant were given after the initial injection and sera was collected 7 and 14 days after each injection, preabsorbed with BSA and assayed by immunohistochemical reactivity on sections of sea cucumber gonad and by dot blot.
Immunohistochemistry
Indirect immunofluorescence method was used to localise GFSKLYFamide-like immunoreactivity in the female gonad of H. scabra using frozen sections. Tissues were processed for immunohistochemical analysis, thus animals were anaesthetized in ice for about 30 min before dissection. Dissected tissues were fixed immediately in 4% paraformaldehyde for 5 to 24 h at 4°C, washed three times in PBS for 10 min each and cryoprotected in 30% sucrose overnight. Sections of 7 μm thickness were cut with a cryostat (LIECA CM 1850), mounted on poly-L-lysine-coated slides and permeabilized in PBS-Triton X-100 (0.1%) for 5 min before blocking with normal goat serum (1:50 in PBS) for 1 h. Sections were incubated overnight at room temperature with primary antibody (1:1000 in PBS) followed by three washes in PBS-Tween 20 (0.05%) for 10 min each. This was followed by 1 to 2 h incubation in Alexa 488-conjugated goat anti-rabbit IgG (1:500 in PBS) at room temperature. Sections were then washed 3 times, 10 min each, in PBS-Tween 20. Slides were incubated in TOPO-3 (1:500 in PBS) at room temperature for 1 h then rinsed in PBS-Tween 20 (0.05%) and mounted in buffered glycerol (pH 8.6). Tissues were examined and photographs taken with an Olympus Confocal laser scanning microscope (FV 1000). Images were processed using OLYMPUS FLOVIEW 1.7b viewer and Adobe Photoshop CS3. Positive and negative control tests were run on a nitrocellulose membrane using a Dot Immunoblot Assay (DIA) protocol previously developed by us. The DIA protocol involves spotting 0.5 μl of synthetic GFSKLYFamide neuropeptide on three 0.22 mm nitrocellulose membranes. The membranes were air-dried at room temperature for 30 min and baked at 100°C for 30 min. After cooling to room temperature, the membranes were washed for 10 min in a small volume of 0.1M Tris buffer (pH 7.4) with 0.05% Tween 20 (v/v; Buffer 1). After washing, the membranes were incubated for 60 min in a blocking solution containing 3% BSA in 0.1M Tris buffer (pH 7.4). Following blocking, the membranes were incubated overnight in GFSKLYFamide primary antibody (1:10,000) diluted in Buffer 1. After the removal of primary antibody, the membranes were washed 3 times for 5 min each in Buffer 1 and incubated in AP-conjugated goat anti-rabbit antibody (1:500 in Buffer 1) for 1 h. Membranes were again washed 3 times for 5 min each in Buffer 1, developed with NBT/BCIP (1:100 in development buffer) and then washed in distilled water, air-dried and photographed. Preabsorption control was done by substituting the primary antibody with preabsorbed antibody or PBS. Working dilutions of GFSKLYFamide antibody (1: 1000 in PBS) were preabsorbed overnight at 4°C with 100, 50 and 10 ng/μl of synthetic GFSKLYFamide neuropeptide.
The polyclonal antibody used in this has been characterized and used extensively in previous studies. It was reported to show high specificity for GFSKLYFamide neuropeptide and does not react against CCK, galanin, proctolin, and CARP. It was also reported that in preabsorption controls, GFSKLYFamide at 1 and 10 pg/ml eliminated the observed immunoreactivity from the digestive system (Diaz-Miranda et al., 1995).
A matured gonad of H. scabra is shown in Figure 1. Results from our positive and negative control test using DIA show that GFSKLYFamide antibody, acting as primary antibody, reacted with synthetic GFSKLYFamide neuropeptide while preabsorbed GFSKLYFamide antibody, acting as primary antibody, did not elicit any reaction with synthetic GFSKLYFamide neuropeptide (Figure 2). Also, substituting the primary antibody with PBS failed to elicit any reactions. This shows that GFSKLYFamide polyclonal antibody that was used is specific for GFSKLYFamide neuropeptide.
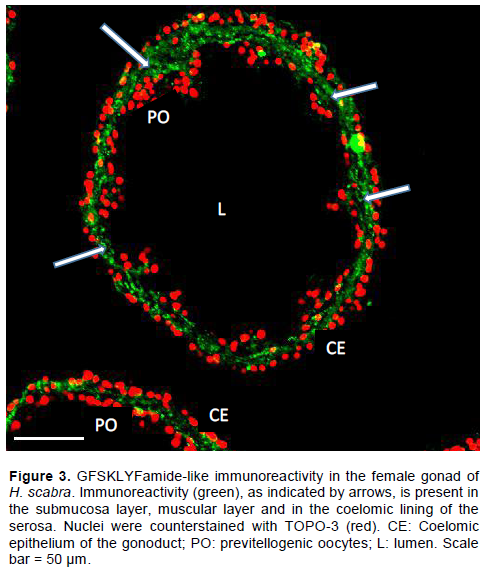
Results also show the localization of GFSKLYFamide-like immune-reactivity in the female gonad of the sea cucumber H. scabra. GFSKLYFamide-like immune-reactivity was expressed in the sub-epithelial fibres of the coelomic epithelium (CE). Also from the result, it was observed that the lumen (L) of the gonad was almost empty except for few previtellogenic oocytes (PO) lying within the germinal epithelium of the gonad (Figure 3).
Neuropeptides are signalling molecules that are produced by neurons and they often play the role of neurotransmitters and neuromodulators in several physiological systems (Semmens and Elphick, 2017). In this study, GFSKLYFamide-like immunireactivity was localized in the female gonad of the sea cucumber H. scabra. The GFSKLYFamide-like immunoreactivity was specifically expressed in the sub-epithelial fibres of the coelomic epithelium of female H. scabra. This result is similar to an earlier report in H. glaberrima where it was observed that the coelomic epithelium and sub-epithelial fibres of both male and female gonads of H. glaberrima expressed GFSKYLFamide-like immunoreactivity (Diaz-Miranda et al., 1995). Although a major difference between this report and that of the H. glaberrima study is the fact that the current study was limited to only the female species of H. scabra and GFSKLYFamide-like immunoreactivity is exclusively expressed in the sub-epithelial fibres of female H. scabra.
GFSKLYFamide neuropeptide could be a key signalling molecule in Echinoderms owing to the localization of immunoreactivity against this neuropeptide in the second sea cucumber species, as demonstrated in this study. It has already been suggested that GFSKLYFamide plays a significant role in the control of multiple action systems including digestion, respiration, circulation, locomotion and even reproduction (Diaz-Miranda et al., 1995). The present study confirms earlier reports that GFSKLYFamide-like immunoreactivity occurs throughout the nervous system of the sea cucumber (Diaz-Miranda et al., 1995; Elphick, 2014; Ajayi and Withyachumnarnkul, 2015).
H. scabra is a tropical species, found in warm shallow waters particularly in the indo-pacific region (Purcell et al., 2012). This species of sea cucumber is commonly eaten due to its medicinal properties; hence, its great commercial value. As a result of its overexploitation, it is liable to becoming an endangered species. The sea cucumber, H. scabra have separate sexes that are difficult to distinguish unless at spawning. Reproduction in H. scabra occurs through a process that involves broadcast spawning of gametes into the seawater where sperm make direct contact with eggs to begin the process of fertilization (Hamel et al., 2001). The reproductive cycle of H. scabra comprises a resting (resorption), growing maturity, spawning, post-spawning stages (Yoichi and Michiyasu, 2014). The estimated size of sexually matured H. scabra is 210 mm in females and 213 mm in males (James and James, 1994; Abdul-reza et al., 2012). The gonad of female H. scabra are in various stages of maturity for example in immature gonad, tubules are single and short, in maturing gonad the tubules are long, yellowish and germinal cells are visible (Baskar, 1994). In adult female gonad, the tubule, in addition to being long is yellowish in colour as well as branched (Baskar, 1994). The average weight of female gonad is 31 g while in male is 24 g (Conand, 1993). Content of the gonads in H. scabra include filamentous tubules which form a tuft that is attached to the dorsal mesentery in the coelomic cavity (Conand, 1993). Proceeding into the dorsal mesentery is the gonoduct which is lined by ciliated epithelium (Purcell et al., 2012). The gonoduct in female H. scabra is lined by ciliated epithelia cells that are directed towards the lumen. The gonoduct also consist of connective tissue and coelomic epithelium lining it externally. Embedded in the germinal epithelium are numerous oogogonial cells (oocytes) that have nuclei which can be seen when lightly stained. The oocytes of H. scabra varies in shape from pyramidal to elongated and club-shaped (Hamel et al., 2001). In this study, it is shown that the lumen of the gonad was almost empty except for few previtellogenic oocytes lying within the germinal epithelium of the gonad. This suggests that female H. scabra used may have spawned just before it was obtained and used for the experiment. Female gonadal tubules of H. scabra have been reported to be shorter and wider, containing few oocytes after spawning ((Hamel et al., 2010).
This study has shown that GFSKLYFamide-like immunoreactivity is present in the female gonad of the sea cucumber, H. scabra. This, to the best of our knowledge, is the first report providing evidence for the existence of GFSKLYFamide neuropeptide in the female gonad of H. scabra. It is however suggested that further studies be carried out to determine the structure of the peptide(s) producing this immunoreactivity in H. scabra, as well as an investigation into the existence of GFSKLYFamide-like immunoreactivity in the male gonad of H. scabra.
The authors have not declared any conflict of interests.
Authors appreciate support from the Center of Excellence for Shrimp Molecular Biology and Biotechnology (CENTEX Shrimp), Faculty of Science, Mahidol University, Thailand as well as by Professor García-Arrarás (University of Puerto Rico, USA) for the generosity in providing antibody.
REFERENCES
|
Abdul-reza D, Musa K, Dariush M, Majid A, Seyyed AN (2012). Holothuria scabra (Holothuroidea: Aspidochirotida): ï¬rst record of a highly valued sea cucumber, in the Persian Gulf, Iran. Available
View [accessed Nov 28 2017].
|
|
|
|
Ajayi A, Withyachumnarnkul B (2013). Presence and distribution of FMRFamide-like immunoreactivity in the sea cucumber Holothuria scabra (Jaeger, 1833). Zoomorphology 132(3):285-300.
Crossref
|
|
|
|
|
Ajayi A, Withyachumnarnkul B (2015). Expression of GFSKLYFamide-like neuropeptide in the digestive system of the sea cucumber Holothuria scabra (Echinodermata). African Journal of Biotechnology 14(25):2124-2129.
Crossref
|
|
|
|
|
Baskar BK (1994). Systematics, biology, ecology and zoogeography of holothurians: Some observations on the biology of the holothurian Holothuria (metriatyla) scabra (Jaeger). CMFRI Bulletin 46:39-43.
|
|
|
|
|
Conand C (1993). Reproductive biology of the holothurians from the major communities of the New Caledonian Lagoon. Marine Biology 116(3):439-450.
Crossref
|
|
|
|
|
Conand C (1998). Overexploitaion of the present world sea cucumber fisheries and perspective in mariculture. In "9th international Echinoderm conference, San Francisco" (R.Mooi and M. Telford, eds.). pp. 449-454.
|
|
|
|
|
Díaz-Miranda L, Blanco RE, García-Arrarás JE (1995). Localization of the heptapeptide GFSKLYFamide in the sea cucumber Holothuria glaberrima (Echinodermata): a light and electron microscopic study. Journal of Comparative Neurology 352(4):626-640.
Crossref
|
|
|
|
|
Díaz-Miranda L, Price DA, Greenberg MJ, Lee TD, Doble KE, García-Arrarás JE (1992). Characterization of two neuropeptides from the sea cucumber Holothuria glaberrima. The Biological Bulletin 182(2):241-247.
Crossref
|
|
|
|
|
Elphick MR, Melarange R (2001). Neural control of muscle relaxation in echinoderms. Journal of Experimental Biology 204(5):875-885.
|
|
|
|
|
Elphick M (2014). SALMFamide salmagundi: The biology of a neuropeptide family in echinoderms. General and Comparative Endocrinology 205:23-35.
Crossref
|
|
|
|
|
Hamel JF, Conand C, Pawson DL, Mercier A (2001). The sea cucumber Holothuria Scabra (Holothuroidea: Echinodermata): Its biology and exploitation as beche-de-mer. Advances in Marine Biology 41:129-223.
Crossref
|
|
|
|
|
Hamel JF, Mercier A, Conand C, Purcell S, Toral-Granda TG, Gamboa R (2010). Holothuria scabra. IUCN Red List of Threatened Species. International Union for Conservation of Nature. Retrieved 1 June 2017.
|
|
|
|
|
James PR, James DB (1994). Management of Ber-der-mer industry in India. In "proceedings of the natural workshop on Beche-der-mer" (K. Rengarajan and D.B James, eds.) pp. 17-22.
|
|
|
|
|
Melarange R, Potton DJ, Thorndyke MC, Elphick MR (1999). SALMFamide neuropeptides cause relaxation and eversion of the cardiac stomach in starfish. Proceedings of the Royal Society of London B: Biological Sciences, 266(1430):1785-1789.
Crossref
|
|
|
|
|
Purcell SW, Samyn Y, Conand C (2012). Commercially important sea cucumbers of the world. FAO species catalogue for fishery purpose, Rome. pp. 80-82. Available at:
View
|
|
|
|
|
Semmens DC, Elphick MR (2017). The evolution of neuropeptide signalling: insights from echinoderms. Oxford University Press, pp. 1-11.
|
|
|
|
|
Yoichi M, Michiyasu Y (2014). The Reproductive Cycle of the Sea Cucumber Holothuria scabra around Shioya Fishing Port, Okinawa Prefecture. Aquaculture Science 62(3):235-242.
|
|