Full Length Research Paper

ABSTRACT
Intestinal schistosomiasis remains a major cause of morbidity and mortality in most parts of the world, despite decades of widespread chemotherapy use. The present study determined the intermediate host proteins expressing the immunogenic properties against the parasite by determining the efficacy. Digestive gland and foot parts were isolated by dissection from the Biomphalaria pfeifferi, processed and the concentration was determined. The proteins were used to immunize the experimental groups of BALB/c mice which were then challenged with the parasite to determine the efficacy. The results show the proteins were efficacious with foot and digestive gland soluble proteins having 59.1 and 40.9% worm reduction, respectively. They also stimulated the production of effective cytokines, immunoglobulins and reduced hepatic pathology.
Key words: Schistosomiasis, immunogenic, Biomphalaria pfeifferi, BALB/c mice.
INTRODUCTION
Schistosomiasis is a parasitic illness caused by trematodes (blood flukes) of the Schistosoma genus (Jauréguiberry et al., 2010). Schistosomiasis is the third most distressing illnesses globally, after malaria and intestinal helminthiasis. It is a source of mortality and morbidity in developing nations: Asia, the Middle East, the Caribbean, South America, and Africa. Above 207 million individuals, 85% of those that reside in Africa are usually infected with schistosomiasis (Góes et al., 2012). Current methods used for controlling intestinal schistosomiasis which includes chemotherapy and molluscicides do not give lasting protection. A vaccine is, therefore, an ideal cost-effective alternative method for the control of schistosomiasis. Despite the discovery and publication of numerous potentially promising vaccine candidates from Schistosoma mansoni and, to a lesser extent, Schistosoma haematobium, only one vaccine, namely, BILHVAX, or the 28-kDa GST from S. haematobium, entered clinical trials (Reynolds et al., 1994).
Common antigens between different species of Schistosoma and their intermediate host have been reported (Khattab et al., 2010) providing a reason why the parasite is able to develop in this host. Determination of immunogenic proteins can therefore be used as an ideal method for vaccine candidate, which can be used to control schistosomiasis as an alternative to mass drug treatment.
Schistosomes are not able to proliferate inside their mammalian hosts. As a result, non-sterilizing vaccine or naturally-acquired immunity could considerably reduce disease transmission and human pathology. Vaccination that would be developed against schistosomes would be directed towards averting the infection and or decreasing the fecundity of the parasite (Fong, 2013). A decrease in the numbers of the worms is the ‘gold principal’ for the vaccine development against schistosomes, with the migrating phase of the schistosomules being the probable target of major vaccine of protective immune responses.
Nevertheless, because schistosome eggs are responsible for transmission and pathology, a vaccine that is directed at egg viability and parasite fecundity also seems entirely fitting. While the standard induce 50 to 70% (above 90% in some scenarios) protection experienced in experimental animals and further immunizations amplifies this degree further, it might turn out to be premature to go after irradiated schistosome vaccines for use by humans, although their production for veterinary use is possible (Ziniel et al., 2015). The concept is proven, and most of the requisite approaches, even though they need up scaling and refining are published.
MATERIALS AND METHODS
Host and parasite
BALB/c mice were maintained at laboratories of Institute of Primate Research (IPR) in Kenya. The animals were kept in standard environmental condition of 25.0±2°C temperature, 55±10% relative humidity and 12 h light/dark cycle. They were fed on rodent pellets and water ad libitum. Biomphalaria pfeifferi, the S. mansoni intermediate host, were obtained from Mwea, Kirinyaga County and maintained at the Malacology room at Institute of Primate Research IPR, at 22 to 25°C, 12 h light/dark and were fed on dried lettuce. Five weeks post infection (pi), of the snails with five to eight miracidia, shedding of cercariae was done on snails in a beaker with 20 ml of water from the snail tanks by exposing them to light (100 watts bulb shielded with glass to safeguard the snails from the heat) for 3 h to shed the cercariae. Cercarial concentration in well stirred suspension was estimated by counting three 50 µl aliquots under the dissecting microscope. An average of cercarial number was taken and used to calculate the concentration of cercariae per milliliter. This was used to determine the volume of the suspension that was required to infect each mouse with 250 cercariae. This was done according to Smithers and Terry (1965).
Experimental design
BALB/c mice were divided into three experimental groups vaccinated with digestive gland (DG), with foot protein (FT), and an infected control (IC). DG and FT groups were immunized and given two boosters of their specified proteins. All the three groups were challenged with 250 S. mansoni cercariae after the final boost. Blood was collected at the beginning of the study and after a 2 week interval thereafter for 6 weeks. Serum was used for the determination of cytokine and IgG using ELISA. Perfusion, gross and histo-pathology were carried out at week 6 pi.
Preparation of immunizing agent
B. pfeifferi snails were used to prepare soluble protein extracts. DG and FT were obtained from the snail. Foot and digestive gland were obtained from the snail under a dissecting microscope. Using a scarpel, the FT and DG were incised and placed in labeled Eppendorf tubes containing phosphate buffered saline (PBS ×1) as described in our laboratory (Kuria et al., 2012). The two tissues were homogenized separately in a glass mortar and pestle. The homogenate was then centrifuged for 1 h at 14,000 g at 4°C. The supernatant was obtained and assayed for protein concentration using the microtitre plate technique. The protein concentrations were read using ELISA reader at wavelength of 595 nm (Yole et al., 1996). The foot and digestive gland soluble proteins were both 1.44 mg/ml, respectively. The protein extracts were stored at -20°C until required.
Vaccination and challenge
Both DG group receiving soluble protein from the snail digestive gland and FT receiving soluble protein from the foot protein were immunized with 50 μg of the specified soluble protein extract in 100 μl of complete Freund’s adjuvant intraperitoneally. Each of the groups received 2 boosters at weeks 2 and 4 of 25 μg of specified protein extract in 100 μl of incomplete Freund’s adjuvant intraperitoneally. One week after the final vaccination, the two groups of immunized mice, DG and FT, and a control IC, were challenged with cercariae. The mice were shaved on the stomach wall using a surgical blade and anaesthetized with ketamine/xylazine mixture (20:1 made by adding 0.5 ml of xylazine to 10 ml of ketamine). Anaesthesia was delivered as 0.02 ml per 30 g mouse body weight using a syringe and needle intraperitoneally. Once unconscious, the mice were arranged on a wooden infecting rack. Cotton wool, dipped in water, was used to clean and wet the shaven area to allow easy penetration of the cercariae. Metal rings, 1 cm diameter, were placed on the shaven area and cercaria suspension was dispensed into the metal ring using a 1000 µl micro pipette and allowed to penetrate for 30 min (Smithers and Terry, 1965).
Preparation of serum
At the 2nd, 4th and 6th weeks post-challenge, blood was obtained from anaesthetized (mixture of Rompun and Ketamine in ratio of 20:1) mice from all the groups (DG, FT, IC) by heart puncture. Serum was prepared from the blood and stored at -20°C before use in cytokine and IgG enzyme linked immunosorbent assay (ELISA).
Immunological assays
ELISA for Schistosome specific IgG
Nunc-ImmunoTM (MaxiSorpTMSurface) ELISA plates were coated overnight with 50 µl of 10 µg/ml soluble adult worm antigen preparation (SWAP) and 0 to 3 h release protein (This contains the proteins released by penetrating schistosomula between 0 and 3 h of penetration), diluted in bicarbonate buffer, pH 7.4 and incubated overnight at 4°C. The proteins were prepared in the laboratory according to Yole et al. (1996). The plates were washed six times using wash buffer (0.5% Tween 20 in PBS). This was followed by blocking of the non-specific binding sites with 100 µl 3% BSA in PBS for 1 h at 37°C and washing off unbound BSA six times with washing buffer. Fifty microliters of diluted (1:200) serum samples were dispensed into each well in duplicates and incubated for 2 h at 37°C, and then washed. After washing the unbound serum, 50 µl of 1:2000 peroxidase conjugated goat anti-mouse IgG was dispensed into the wells and incubated for 1 h at 37°C. The unbound conjugate was washed off as before and 50 µl TMB microwell peroxide substrate added. The plates were incubated at 37°C in the dark for 30 min. The optical density was read at 630 nm in an ELISA microplate BioTekR reader.
Cytokine profiles (Th1 and Th2) using flow cytometry
The reagents used were obtained from BDTM Biosciences (Califonia, USA) BD cytometric bead array (CBA) mouse Th1/Th2 cytokine kit. This contained all the necessary reagents and capture antibodies specific for Interleukin-2 (IL-2), IL-4, IL-5, Interferon-γ (IFN-γ), and tumor necrosis factor (TNF) in a single serum sample. The kit performance was optimized for analysis of physiologically relevant concentrations (pg/ml levels) of specific cytokine proteins in serum samples. The serum samples were retrieved and 50 µl of each added to labelled acquisition tube. 50 µl of mice Th1/Th2 PE detection reagent was added to all the tubes (that is, Th1/Th2 cytokine standard and serum samples). The mixture was incubated in darkness for 2 h at room temperature. 1 ml of wash buffer was then added to each tube and the tubes were centrifuged at 200 g for 5 min. The supernatant from each tube was carefully aspirated and discarded. 300 µl of the wash buffer was then added in each tube to resuspend the bead pellet after which the suspension was loaded on BD FACS CaliburTM flow cytometer. The standards were acquired from the lowest (0 pg/ml) to the highest (Top Standard) concentration, followed by the test samples. The data was captured using BD FACS CaliburTM flow cytometer, BD CellQuestTMPro version 5.2.1 and FCAP Array Software version 1 was used in the analysis of cytokine profiles.
Gross pathology, perfusion and worm recovery
At the 6th week pI, the abdominal cavities of mice in group DG, FT and IC were opened. Gross pathology examination focused on the general and overt appearance of the liver. The observations considered included inflammation, adhesions and presence of granulomas. Granulomas appear as numerous raised pinheads sized foci distributed over the surface of the liver. The grading of the granulomas was as follows; none; 0, few; 1 - 3, moderate; 3 - 10 and severe; more than 10 granulomas per lobe. The inflammation was graded as follows: normal, slightly inflamed (>normal), inflamed (>slightly inflamed); adhesions were either present or absent (Yole et al., 1996). The perfusate was collected in a 20 cm glass Petri-dish. The perfusate containing the recovered worms were transferred into urine jar and topped with phosphate buffered saline (PBS). After the worms had settled, the supernatant was sucked out, and the settling procedure repeated three times. When the supernatant was clear, the worms were then placed on a Petri dish containing PBS and then counted. Any worms still left in the mesenteries/liver were recovered by soaking the mouse in a Petri dish containing PBS for at least 30 min, to allow the worms to crawl out. Percentage worm recovery and reduction for each group was calculated as shown in the formulae:

Histopathology
After perfusion and recovery of worms from each mouse, the liver was fixed in 10% formalin for 2 weeks. A representative portion was obtained from each tube and placed in holding tissue cassettes and washed in running water overnight to remove excess formalin. The tissues were then processed using an automatic tissue processor which dehydrated sequentially in increasing concentrations of alcohols of 85, 95, and 100% at hourly stepped intervals to achieve optimum dehydration. The tissues were then cleared off alcohol twice in two changes of toluene. Infiltration with hot paraffin wax was done for 3 h. The tissues were then embedded in fresh molten paraffin wax (Sherwood Medical Co. USA) and allowed to dry. The embedded tissues were sectioned at 6 microns thicknesses with a rotary microtome (Leitz, Germany). The thin tissue sections were mounted on glass slides and stained with haematoxylin and eosin. The tissue sections were observed under light microscope for any pathological changes. Granulomas sizes were measured. The mean size was calculated from average of width and length (Farah et al., 2000).
Statistical analysis
Data were analyzed using Statistical Package for Social Sciences (SPSS). Statistical significance between the groups was tested using analysis of variance. The results were presented as mean±standard deviation (SD) and P value was considered significant if less than 0.05.
RESULTS
Worm recovery
The mean worm burden of IC was 22±3.78 which represents 8.8% maturation of penetrant cercariae. Worm burden of FT immunized group was 9±1.53, which represents 40.9% worm recovery which was significantly different (p<0.05) from the infected control. The DG immunized group had a mean worm burden of 13±2.28, representing 59.1% worm recovery and was significantly different (p<0.05) from the infected control. There was no significant difference between FT and DG (p>0.05). The group immunized with FT had the highest worm reduction at 59.1% as compared to 40.9% in the group immunized with DG.
Cellular responses
Serum of mice from each group was assayed for IL-2, IL-5, IFN-γ, IL-4 and TNF for the different time points following challenge with S. mansoni.
IL-2 level in serum after challenge with S. mansoni
Figure 1 shows the concentration of IL-2 at different time points from weeks 0 to 6. At challenge (WK 0), the concentration in DG (8.80±1.13 pg/ml), was significantly higher than IC (6.14±0.16 pg/ml) (p<0.05). The concentration rose sharply to a peak at week 2 (16.69±2.94 pg/ml) followed by a steadily decline to the lowest concentration at week 4 (8.45±0.70 pg/ml) and week 6 (3.15±0.59 pg/ml). The concentration in FT at week 0 (5.90±0.13 pg/ml) was not significantly different (p>0.05) from that of IC (6.14±0.16 pg/ml). There was no change in the concentration at week 2 (6.51±0.23 pg/ml) post challenge. However, at the 4th week, the concentration rose to 12.73±2.71 pg/ml followed by a drop to 6.01±0.15 pg/ml at week 6.

IL-4 production in serum after challenge with S. mansoni
After the cytokine assay for IL-4, it was observed that there was no detectable or significant production of the cytokine. This could be supported by Mountford et al. (2001)where they pointed out that IL-4 was not an important cytokine component in the immunity of schistosome in mice
IL-5 production in serum after challenge with S. mansoni
Figure 2 shows the concentration of IL-5 at different time points from week 0 to week 6. At challenge (WK 0), the concentration of IL-5 in DG (4.19±0.24 pg/ml) was not significantly different from IC (4.53±0.21 pg/ml) (p>0.05). The concentration rose steadily at week 2 (5.93±0.24 pg/ml), week 4 (6.97±0.34 pg/ml) and reached peak at week 6 (7.77±0.86 pg/ml). The concentration of IL-5 in FT at week 0 (9.35±0.78 pg/ml) was at its peak and significantly different from DG and IC. The concentration declined significantly at week 2 (6.53±0.29 pg/ml) followed by a significant rise at week 4 (7.89±0.48 pg/ml). At week 6 (5.16±0.53 pg/ml), the concentration had reduced significantly compared to DG and IC.

IFN-γ concentration in serum after challenge with S. mansoni
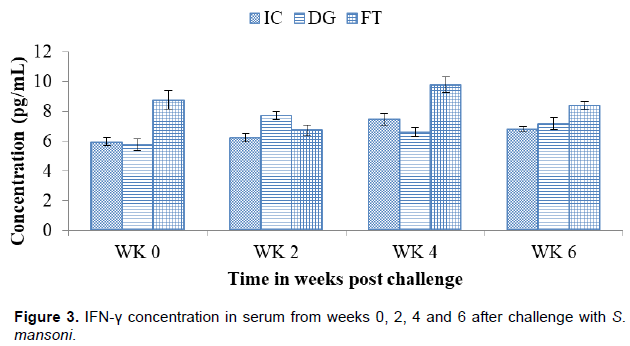
Figure 3 shows the concentration of IFN-γ at different time points from week 0 to week 6. At challenge (WK 0), DG (5.79±0.38 pg/ml) and IC (5.97±0.28 pg/ml) had no significant difference in their concentrations. The level rose to its peak at week 2 (7.74±0.28 pg/ml) then declined significantly at week 4 (6.61±0.29 pg/ml). At week 6, the concentration (7.19±0.43 pg/ml) rose with no significant difference. The concentration in FT at week 0 (8.77±0.61 pg/ml) was significantly higher than IC. The concentration declined at week 2 (6.74±0.36 pg/ml), followed by an increase and a decrease at the 4th (9.79±0.51 pg/ml) and 6th (5.97±0.28 pg/ml) weeks, respectively.
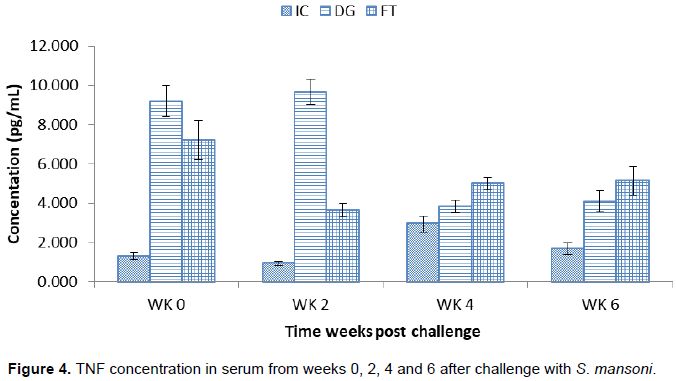
TNF concentration in serum after challenge with S. mansoni
Figure 4 shows the concentration of TNF at different time points from week 0 to week 6. The concentration in DG at challenge (9.18±0.06 pg/ml) was significantly higher than IC (1.31±0.17 pg/ml). The concentration rose to peak at week 2 (9.66±0.05 pg/ml) followed by a significant decline at week 4 (3.85±0.05 pg/ml). At week 6, the concentration (4.09±0.07 pg/ml) rose slightly with no significant difference. The concentration in FT at week 0 (7.22±1.01 pg/ml) was significantly higher than IC which then declined at week 2 (3.64±0.35 pg/ml) significantly. At week 4 (5.01±0.33 pg/ml) and week 6 (5.14±0.72 pg/ml), the concentration increased steadily.
Schistosome-specific IgG antibody concentration for 0 to 3 h antigen
Figure 5 shows the responses of IgG for 0 to 3 h antigen at different time points from week 0 to week 6. The concentration in DG at challenge (0.15±0.02 OD) was significantly higher than IC (0.5±0.01 OD) significantly. The concentration rose steadily at week 2 (0.49±0.01 OD) and reached peak at week 4 (0.71±0.02 OD) with significant difference. At week 6 (0.42±0.02 OD) the concentration declined with significant difference. FT at week 0 (0.57±0.01 OD) was significantly higher than IC. The level rose to peak at week 2 (0.67±0.02 OD) significantly which then declined steadily at week 4 (0.61±0.02 OD) and week 6 (0.51±0.02 OD) significantly. The levels were higher than the background significantly.

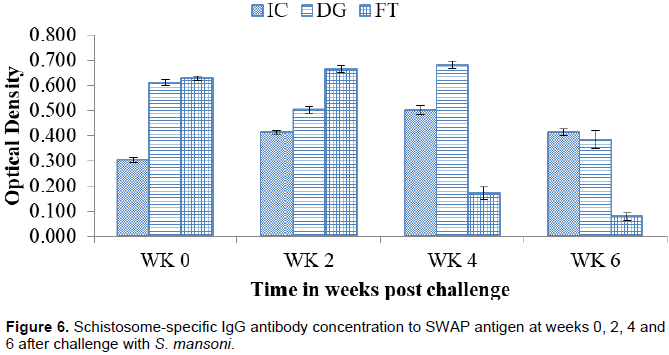
Schistosome-specific IgG antibody responses to SWAP antigen
Figure 6 shows the responses of IgG to SWAP antigen at different time points from week 0 to week 6. The concentration of DG at challenge (0.73±0.01 OD) was higher than IC (0.30±0.01 OD) significantly. The concentration declined at week 2 (0.50±0.01 OD) but rose to peak significantly at week 4 (0.68±0.01 OD). At week 6 (0.26±0.04 OD) the concentration declined significantly. FT at week 0 (0.63±0.01 OD) was significantly higher than IC. The concentration rose to peak significantly at week 2 (0.66±0.02 OD). The concentration declined steadily at week 4 (0.17±0.03 OD) and week 6 (0.36±0.02 OD) significantly.
Pathology
Gross pathology
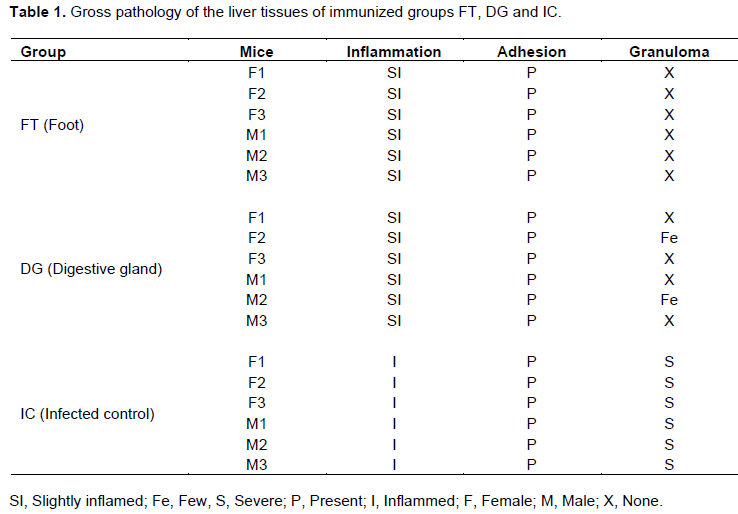
Gross pathology is as shown in Table 1. The liver tissues of group FT appeared normal with slight inflammation. All the liver tissues had adhesions. Granulomas were not observed in the entire group. In group DG, all the liver tissues were slightly inflamed (Figure 7). All liver tissues had adhesions and two of the liver tissues from two mice had few granulomas, while the rest had none. In group IC, all liver tissues were inflammed and had adhesions (Figure 8). The granulomas were severe in all the mice. The grading of the granulomas was as follows: none, 0; few, 1 - 3; moderate, 3 - 10; and severe, more than 10 granulomas, per lobe. The inflammation was graded as follows: Normal, slightly inflamed (> normal), inflamed (> slightly inflamed); adhesions were either present or absent (Yole et al., 1996).
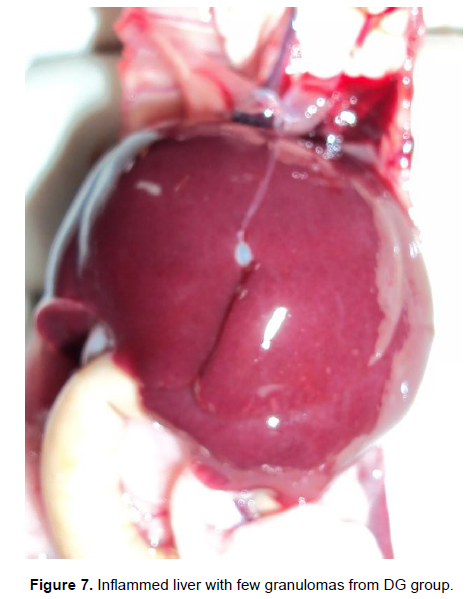
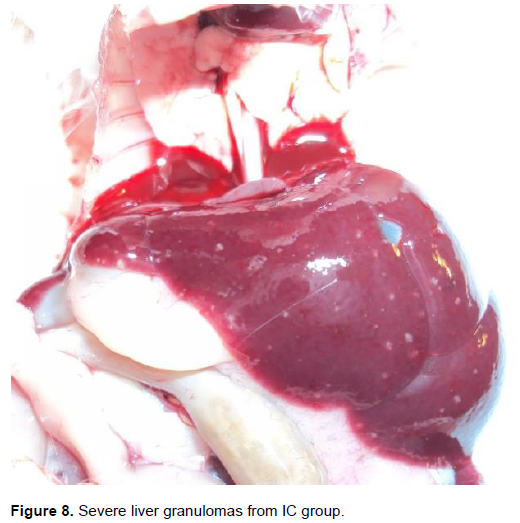
Histopathology
Granuloma sizes were determined using a microscope fitted with a micrometer. The sizes were calculated as mean of the length and width of the granuloma (Farah et al., 2000)as shown in Table 2. In the immunized groups, FT and DG, no granulomas were observed. Group IC had granulomas which measured 1.85±0.34 (Figure 9). When the immunized, FT and DG groups were compared, there was no significant difference (p>0.05) between them. When the immunized groups were compared to IC, there was significant difference (p<0.05).

DISCUSSION
The worm maturation in BALB/c mice was 8.8%. This maturation value observed in the study was different from the range previously reported for rodents (laboratory mouse) 20 to 50% (Yole et al., 1996). BALB/c mice have been found to be a good model for parasitology and immunology since they are permissive definitive hosts and support the full growth, development and sexual reproduction of the parasite (Ross et al., 2002).
The results show that the immunizing agents, FT and DG, were able to invoke protection in BALB/c mice leading to high worm reduction. Worm reduction of both FT and DG exceeded the World Health Organization goal of 40% (WHO, 1999).
Speculation is that S. mansoni infections induce an immune response when TH1 versus TH2 pathways correlate with resistance to infection and lies in the source of different cytokines involved. TH1 responses are typically characterized by secretion of IFN-γ, IL-2 and TNF, whereas TH2 responses are characterized by secretion of IL-4 and IL-5 (Wynn et al., 2004). The immunization induced both TH1 and TH2 type responses which plays an important role in anti-schistosome infection by producing cytokines. The production of the cytokine shows that FT and DG stimulated the TH1 cellular response to S. mansoni in mice which leads to resistance of infection.
The trend of IFN-γ in the study agree with those of Flamme et al. (2001)where it was pointed out that during acute infection, mice have been shown to produce elevated levels of IFN-γ. Also, it is important to note that the levels of the IFN-γ depicted in serum in this study are lower than the levels found in other studies which determined the levels in culture supernatants from lymphocyte cells.
The results of this study in the assay for IL-2 are consistent with results found by Pearce et al. (1991)where they sought to investigate the down regulation of TH1 cytokine production accompanying induction of TH2 responses in schistosomiasis. The results indicated that before the onset of egg laying (the prepatent period), T cells from mice secreted a similar pattern of TH1 type cytokines. It is also pointed out that the role of IL-2 might be to promote selected outgrowth of cells to differentiate to the TH2. IL-2 may also be directly involved in TH2 differentiation process since it also mediates a gene activation program (Leonard and Lin, 2000).
IL-5 induces terminal differentiation of activated B cells into antibody-forming cells in mice and enhances proliferation and differentiation of eosinophils precursors into mature eosinophils in mice and humans. Resistance to reinfection has been associated with these responses (Colley and Secor, 2014). IL-5 is a TH2 type cytokine which could be active in facilitating schistosome egg excretion which is mediated by eosinophil production (Fallon and Dunne, 1999). The results are supported by the fact that IL-5 expression decreases as the worm changes from schistosomula to adult worms. This is explained that as adult worms mature, the immune response shifts TH1 towards TH2 due to egg deposition which was not the case in this study.
In this study, after the cytokine assay for IL-4, it was observed that there was no detectable or significant production of the cytokine. This could be supported by Mountford et al. (2001)where it was pointed out that IL-4 was not an important cytokine component in the immunity of schistosome in mice exposed to irradiated cercariae. Similar results were found by Farias et al. (2010)where no IL-4 production was detected despite higher levels of IFN-γ. This could be further supported by the role of IL-4 in the induction of immunoglobulin class switching to immunoglobulin E, which is responsible for mediating allergic reaction (Gilmour and Lavender, 2008)which is a parasitic infection.
TNF is also responsible to hosts response to infectious agents and recruitment of leukocytes at the inflammation sites (El-Sayed et al., 2016). In this study, the levels of TNF were present throughout the experiment period. This is an indication that the immunizing agents stimulated its secretion. This could explain the low recovery of worms due to the fact that it is responsible for recruitment of leukocytes and the presence of IFN-γ. It could also explain why there was no granuloma observed in the experimental groups since it is responsible for down regulation of granuloma formation as pointed out by El-Sayed et al. (2016) that the cytokine has been implicated in the up-regulation of adhesion molecule expression of vascular endothelial which aggregates leukocytes that block parasite migration.
Protective immunity against schistosomiasis involves both cellular and humoral immune responses. In this study, the immunized mice produced significant levels of IgG antibody both in 0 to 3 h and SWAP antigens. The worm recovery agrees with this finding as it was less in the immunized groups since the IgG produced in the 0 to 3 h antigen could have eliminated more parasites at that stage. In SWAP antigen, the production of IgG decreases at week 6, which could be explained by the low worm recovery in immunized groups, hence the low number of adult worm and the decrease in IgG.
The trends observed in the study correlates very well with the presence and production of IL-5 in immunized mice. IL-5 is responsible for production of antibody by inducing terminal differentiation of activated B cells into antibody-forming cells in mice (Takatsu et al., 2009). IFN-γ stimulates the expression of immunoglobulin of the G type (IgG). This explains the levels of IgG in the experimental groups which could have been as a result of higher levels of IFN-γ modulating IL-5.
Granuloma is a tiny pinhead sized foci on the surface of the liver lobe (Yole et al., 1996). This indicates cellular infiltration consisting mainly of eosinophils, macrophages, fibroblast and lymphocytes surrounding a schistosome tissue trapped egg. To the naked eyes, they appear as pin-sized white/cream spots giving the surface a rough texture. They serve as essential host protective function. In FT and DG, all the liver tissues were slightly inflammed and adhesions were present. This was positive indication for an encounter with an infection process. In mice immunized with FT, all had no granulomas. In group immunized with DG, two of the mice had their liver with few granulomas. This could be an indication that most of the parasites did not develop to maturity for egg deposition to cause granuloma formation. On the other hand, the role played by TH1 cellular cytokines such a tumor necrosis factor (TNF) and IFN-γ in regulating cells in granuloma formation being elucidated (Hams et al., 2013). The presence of the few and or moderate granulomas could be attributed to immune evasion mechanisms of the parasites and hence reach maturity to lay eggs. However, both FT and DG were better in reducing pathology than IC. In the positive control group IC, the liver tissues were inflammed, had adhesions and the granulomas were severe indicating that the parasites developed to maturity for egg deposition to take place and cause granuloma formation. This could again be explained in terms of increased immune responses and is supported by the presence of IFN-γ and TNF production which play role in recruitment of the effector cells (Hams et al., 2013).
CONCLUSION
Conclusively, the two immunizing soluble proteins FT and DG were efficacious since they exceeded WHO criteria. The FT is better than DG in worm reduction. Both FT and DG reduce the gross pathology remarkably compared to IC with FT reducing more than DG. In terms of immune responses, FT has more pronounced responses than DG. It is recommended that future studies could assess the synergistic effect of both soluble protein extracts when used together and allow sampling extended beyond week 6 after immunization and ascertain if the protection is long lived.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGMENTS
The authors would like to thank Ngundi Collins, Sammy Kisara, and Esther Kagasi for their technical assistance. Appreciation also goes to National Council for Science Technology and Innovation for the gracious financial support and Institute of Primate Research for allowing the research to be conducted in their facilities.
REFERENCES
|
Colley DG, Secor WE (2014). Immunology of human schistosomiasis. Parasite Immunol. 36(8):347-357. |
|
|
El-Sayed NM, Fathy GM, Sara AR, El-Shafei MA (2016). Cytokine patterns in experimental schistosomiasis mansoni mice treated with silymarin. J. Parasit. Dis. 40(3):922-929. |
|
|
Fallon PG, Dunne DW (1999). Tolerization of Mice to Schistosoma mansoni Egg Antigens Causes Elevated Type 1 and Diminished Type 2 Cytokine Responses and Increased Mortality in Acute Infection. J. Immunol. 162(7):4122-4132. |
|
|
Farah IO, Nyindo M, King CL, Hau J (2000). Hepatic granulomatous response to Schistosoma mansoni eggs in BALB/c mice and olive baboons (Papio cynocephalus anubis). J. Comparative Pathol. 123(1):7-14. |
|
|
Farias LP, Cardoso FC, Miyasato PA, Montoya BO, Tararam CA, Roffato HK, Leite LCC (2010). Schistosoma mansoni Stomatin like protein-2 is located in the tegument and induces partial protection against challenge infection. PLoS Neglected Trop. Dis. 4(2):e597. |
|
|
Flamme ACL, Patton EA, Pearce EJ (2001). Role of Gamma Interferon in the Pathogenesis of Severe Schistosomiasis in Interleukin-4-Deficient Mice. Infect. Immunity 69(12):7445-7452. |
|
|
Fong IW (2013). Challenges in Infectious Diseases. New York, NY: Springer New York. |
|
|
Gilmour J, Lavender P (2008). Control of IL-4 expression in T helper 1 and 2 cells. Immunology 124(4):437-444. |
|
|
Góes VC, Neves RH, Gomes DC, Machado-Silva JR (2012). Effects of high-fat chow on heart tissue in acute and chronic experimental murine schistosomiasis mansoni. Parasitology 139(11):1462–1470. |
|
|
Hams E, Aviello G, Fallon PG (2013). The Schistosoma Granuloma: Friend or Foe? Frontiers in Immunology. P. 4. |
|
|
Jauréguiberry S, Paris L, Caumes E (2010). Acute schistosomiasis, a diagnostic and therapeutic challenge. Clin. Microbiol. Infect. 16(3):225-231. |
|
|
Khattab HH, Soliman AA, El-Bahy MM, El-Sayed NM (2010). Value of purified Schistosoma snails antigens in diagnosis of schistosomiasis. JASMR 5(2):153-158. |
|
|
Kuria KM, Waihenya R, Kutima H, Njoki P, Yole DS (2012). Cellular responses against Schistosoma mansoni in immunized BALB/c mice with soluble proteins from intermediate host, Biomphalaria pfeifferi. Tanzania J. Health Res. 14(4):280-287. |
|
|
Leonard WJ, Lin JX (2000). Cytokine receptor signaling pathways. J. Allergy Clin. Immunol. 105(5):877-888. |
|
|
Mountford AP, Hogg KG, Coulson PS, Brombacher F (2001). Signaling via Interleukin-4 Receptor Chain Is Required for Successful Vaccination against Schistosomiasis in BALB/c Mice. Infect. Immunity 69(1):228-236. |
|
|
Pearce EJ, Caspar P, Grzych JM, Lewis FA, Sher A (1991). Downregulation of Th1 cytokine production accompanies induction of Th2 responses by a parasitic helminth, Schistosoma mansoni. J. Exp. Med. 173(1):159-166. |
|
|
Reynolds SR, Dahl CE, Harn DA (1994). T and B epitope determination and analysis of multiple antigenic peptides for the Schistosoma mansoni experimental vaccine triose-phosphate isomerase. J. Immunol. (Baltimore Md 1950) 152(1):193-200. |
|
|
Ross AGP, Bartley PB, Sleigh AC, Olds GR, Li Y, Williams GM, McManus DP (2002). Schistosomiasis. New England J. Med. 346(16):1212-1220. |
|
|
Smithers SR, Terry RJ (1965). The infection of laboratory hosts with cercariae of Schistosoma mansoni and the recovery of the adult worms. Parasitology 55(4):695-700. |
|
|
Takatsu K, Kouro T, Nagai Y (2009). Interleukin 5 in the link between the innate and acquired immune response. Adv. Immunol. 101:191-36. |
|
|
World health Organization (WHO) (1999). Report of the WHO informal consultation on Schistosomiasis control, Geneva, Document WHO/CDS/CPC/SIP/99.2. |
|
|
Wynn TA, Thompson RW, Cheever AW, Mentink-Kane MM (2004). Immunopathogenesis of schistosomiasis. Immunol. Rev. 201:156-167. |
|
|
Yole DS, Pemberton R, Reid GD, Wilson RA (1996). Protective immunity to Schistosoma mansoni induced in the olive baboon Papio anubis by the irradiated cercaria vaccine. Parasitology 112(1):37-46. |
|
|
Ziniel PD, Karumudi B, Barnard AH, Fisher EMS, Thatcher GRJ, Podust LM, Williams DL (2015). The Schistosoma mansoni Cytochrome P450 (CYP3050A1) Is Essential for Worm Survival and Egg Development. PLoS Neglected Trop. Dis. 9(12):e0004279. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


