Full Length Research Paper

ABSTRACT
Aquatic organisms in the coastal regions are often exposed to many anthropogenic effluents. Kuwait Bay receives considerable quantities of sewage and industrial effluents, and thus, naturally, many endocrine disrupting chemicals (EDCs) find their way into the coastal waters. These EDCs could influence the physiology of the aquatic organisms inhabiting the coastal region. Response of two important local fish of aquaculture importance, bluefin bream (Sparidentax hasta) and yellowfin bream (Acanthopagrus latus) was evaluated by injecting a standard vitellogenin inducer, 17 β-estradiol (17 BEST). The inducer was used at 2, 4 and 6 µg/fish (injected intraperitoneally) to induce vitellogenin production in juvenile fish (150-200 g). There was a dose-dependent Vtg induction response as evidenced by a qualitative immune-dot assay. All levels significantly enhanced Vtg gene expression, as early as one-hour post injection. The expression was noticed to be the highest in liver tissue at 6 h post injection. The bluefin bream was used for amplification of the Vtg gene yielding 282 bp product which was sequenced. This indicated a 97% sequence similarity with the Vtg gene sequence of European seabream, S. aurata.
Key words: Induction, Vtg, EDCs, Kuwait, fish, effluent.
INTRODUCTION
Kuwait and other countries of the Gulf Cooperation Council (GCC) have many common natural exploitable aquatic resources. Bluefin bream, locally called sobaity, and yellowfin bream, called shaeim support a major fishery in the region. There has been a steady decline in the fishable stocks of commercially important species in the region and more so in Kuwait Bay. Several reasons have been attributed to be the cause of such a decline in the fishery in many parts of the world. Chen (2008) stated “whether overall global yield from marine fisheries will decline due to climate change remains unclear; however, regime shift within individual marine ecosystems and trends in fish landing for certain species will likely occur”. This statement fits well the ecosystem such as those of Kuwait Bay which has more anthropological influences and less oceanic influences. Previous studies conducted by the Kuwait Institute for Scientific Research (KISR) have revealed that the anthropological activities have immense influence on the marine ecosystem of the bay. A recent study has shown that the Kuwait Bay receives considerable quantities of endocrine disrupting compounds (EDCs) because of anthropological activities (Al-Jandal et al., 2016 and 2018). The EDCs behave as hormone mimics, estrogens affect the recruitment to fishery through altered reproduction (Oshima et al., 2003; Milla et al., 2011) and /or permanent or irreversible sex change (Kuhl and Brouwer, 2006).
The endocrine functions are one of the important physiological features that play an important role in reproduction and recruitment of a fish species akin to that of vertebrate reproduction. Vitellogenin (Vtg) is considered as an important physiological indicator of endocrine functional disruptions (Hutchinson et al., 2009; Burki et al., 2012; Wheeler et al., 2013). The process of reproductive disruption is the result of hormonal alterations that are due to either natural causes or due to anthropological activities. Anthropological activities have been recently causing increased awareness due to their role in enhancing endocrine disrupting chemicals (EDC) that directly alter reproductive hormonal dynamics and related behavioral defects (Kavlock et al., 1996; Leon-Olea et al., 2014; Dang 2014; 2016).
This study was a part of the project aimed at studying the chemical and biological effects of EDCs in Kuwait coastal waters. We studied the effect of experimental induction of Vtg and the gene expression pattern in two commercially important marine fish species (bluefin bream, S. hasta and yellowfin bream, A. latus) of the region. Also, isolation of the Vtg gene from S. hasta was carried out for a partial characterization and sequence analysis.
MATERIALS AND METHODS
The fish
The bluefin bream (S. hasta) and the yellowfin bream (A. latus) sub-adults in the size range of 100 - 150 g (as the smaller fish of less than 1 year-age are males) were provided by the aquaculture wing of Kuwait Institute for Scientific Research (KISR). The fish were treated with 17 β-estradiol (17 BEST, Sigma- USA) and the induction procedure as described as follows.
Each treatment consisted of 16 fish (16 x 3; for three levels of injection) and 16 as untreated control. At day 0 the fish were acclimatized in fiber glass reinforced plastic tanks (one-ton capacity). On day 3, first injection (2 µg/kg, T1; 4 µg mg/kg, T2; and 6 µg/kg, T3) of 17 BEST with olive oil (intraperitoneal injection). Treatment set, T4, was injected with oil only as a control. The fish were fed commercial seabream pellet feed (Aquamarine, Greece) three times a day. The commercial feed is certified to be free of any soya-based or ingredients of plant origin containing phytoestrogens.
Sampling
Fish serum samples were collected at 1, 6, 24h, 1 week, and day 10. Two fish were sacrificed for collecting different tissue samples (gills, heart, liver, and kidney) for RNA extraction. At 1-week post injection plasma samples for a qualitative dot blot assay were taken. Fish handling was carried out according to stipulated PETA procedures. The fish were anesthetized with 20 ppm of thyme oil during injection, blood sampling and final sacrifice.
On day 17, two fish were sacrificed for RNA extraction as explained above. Third injection for the remaining (4) fish was given at day 17. Day 24 –Two fish (S. hasta only) were sacrificed for collecting tissue samples for RNA extraction, amplicon analysis and sequencing. Plasma samples were collected for a qualitative dot blot assay.
Extraction of RNA and cDNA synthesis
A commercial RNA isolation kit was used for extracting total RNA from the tissue (2-5 mg) samples (gill, heart, liver and kidney) soon after collection or from RNA later (Sigma, USA)-stored samples according to the procedure described in the kit (Ambion Life Technologies, USA) with DNAse digestion to eliminate any interference by the genomic DNA. The extracted RNA was evaluated for its quality using Nanodrop spectrophotometer and visualized by running on 1.2% (wt/volume) agarose gel. Extracted RNA from different tissues was used for synthesizing cDNA (Agilent cDNA synthesis kit).
Amplification of Vtg gene
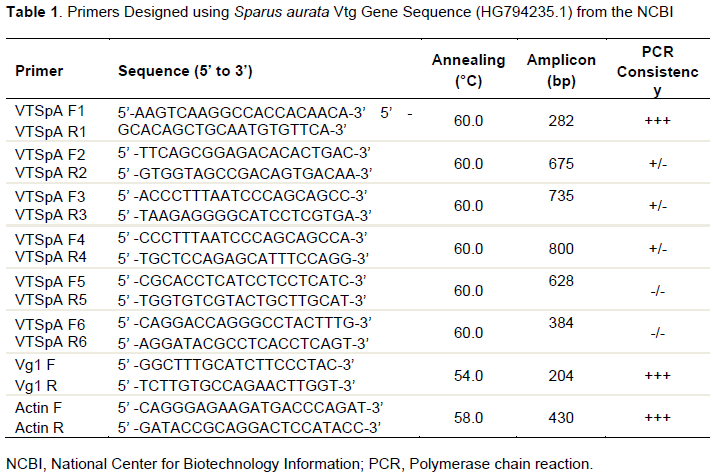
The bluefin and the yellowfin breams are close relative of the European seabream. Hence the Vtg gene sequence (HG794235.1) from the National Center for Biotechnology Information (NCBI) database was used to generate six random sets of primers (Table 1). Amplification of cDNA of the bluefin bream was carried out following the cycle conditions; initial denaturation at 95°C for 10 min followed by 35 cycles of 95°C for 1 min, 60°C for 45 s, 72°C for 30 s and final hold at 72°C for 5 min and 4°C store till electrophoresis. The amplified products were run on 1.2% agarose gel, 282 bp amplicon was excised from the gels and re-amplified to confirm the gene specificity, and then the gels were extracted to yield the amplicons using the gel DNA extraction kit (Qiagen Gel extraction kit). The extracted DNA amplicons were sent to a private molecular biology laboratory for sequencing. The primer set with a high consistency of amplification of samples (VTSpa F1R1) was used for obtaining the sequence information of the amplicon.
Immuno-Dot Assay (dot blot) was carried out with a fixed primary antibody dilution of 1:128, and varying dilutions of the test plasma (terminal sample of experimental Vtg induction) from 1:16 (24) to 1:512 (29) were used. The plasma of different dilutions was spotted (5 µl) on to a pre-wetted nitrocellulose paper (0.40 µl), incubated for 30 min to dry the spot, and the paper was blocked with blocking buffer (3% casein in PBS) at room temperature for 15 min. The paper was washed, incubated with the primary antibody for 1 h at room temperature, washed, and incubated for 1 h at 37°C with the secondary antibody (goat anti-rabbit conjugated with HRP) at a dilution of 1:200. The paper was washed again and treated with the substrate (diamino benzidine, DAB) for 15 to 20 min. The color development process was stopped by washing in distilled water. The color intensity was matched, and the highest dilutions with consistent color were taken as the titer.
RESULTS AND DISCUSSION
Total RNA isolation, cDNA synthesis and gene amplification
The RNA isolation resulted in quality RNA yield as visualized on the agarose gel. The cDNA synthesized produced consistent amplification of Vtg gene and the highest consistency, as visualized on gel electrophoresis, was obtained with VTSpA primers with an expected amplicon size of 282 bp (Figure 1a and b). Sequence similarity analysis revealed 97% similarity (Figure 1c) with the Vtg A gene of European seabream (S. aurata).
Gene expression pattern
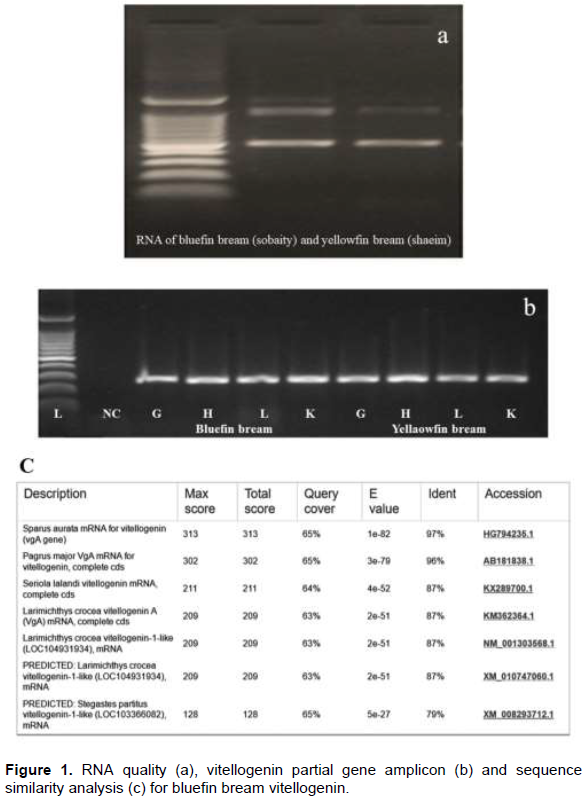
The gene expression pattern with different doses of estrogen (compared with that of the respective tissues from the control fish) was evident from 1 h post injection. The highest levels of gene expression were seen in the kidney tissue followed by liver, heart, and gills. Among various doses, 6 µg/fish produced the highest expression. Expression of Vtg gene in treated fish in different tissues was similar compared to two reference expressions, the actin gene in respective samples and the Vtg gene in untreated control fish.
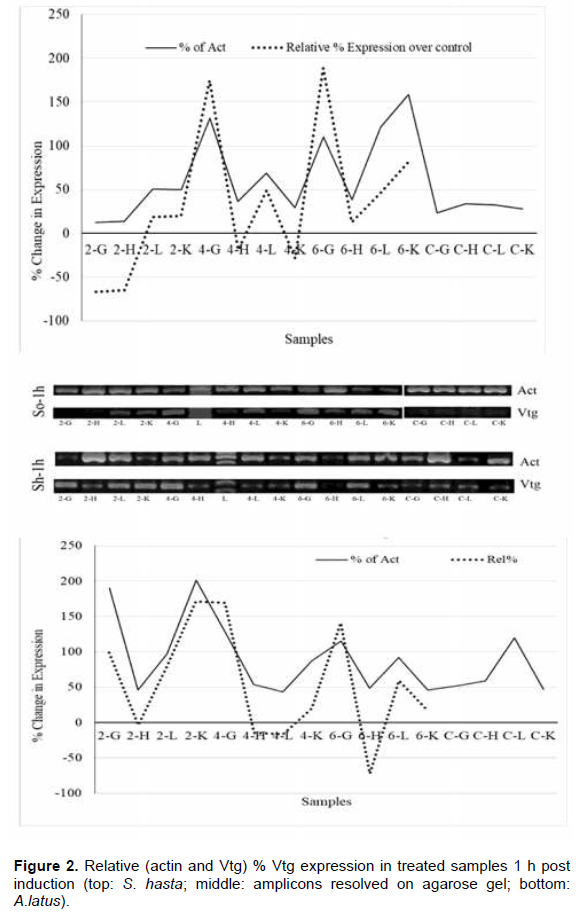
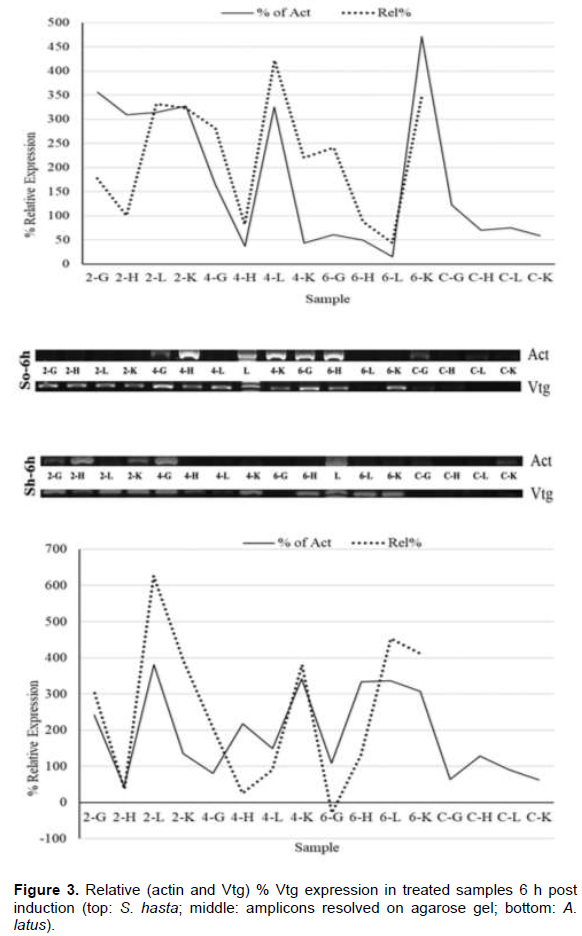
Soon after the injection, at 1 h, all the tissue showed higher VTG expression in comparison to their respective actin genes (Figure 2), while, the lower doses had down regulation in gill and heart tissues. Liver, however, showed upregulation in all the dose regimes. The pattern of expression was also similar between the species. During the entire period of one week, both the species showed an identical pattern of Vtg expression. The highest relative Vtg gene expression was noticed at 6 h post induction in both the breams (Figure 3). Except the liver samples other tissues showed lower or down regulated relative expression at 1d (Figure 4) while, all tissues showed up regulation of Vtg at 1-week post induction (Figure 5). The gene expression results were supported by the results of immune-dot assay (Figure 6) conducted on the plasma samples at the end of the induction experiment.
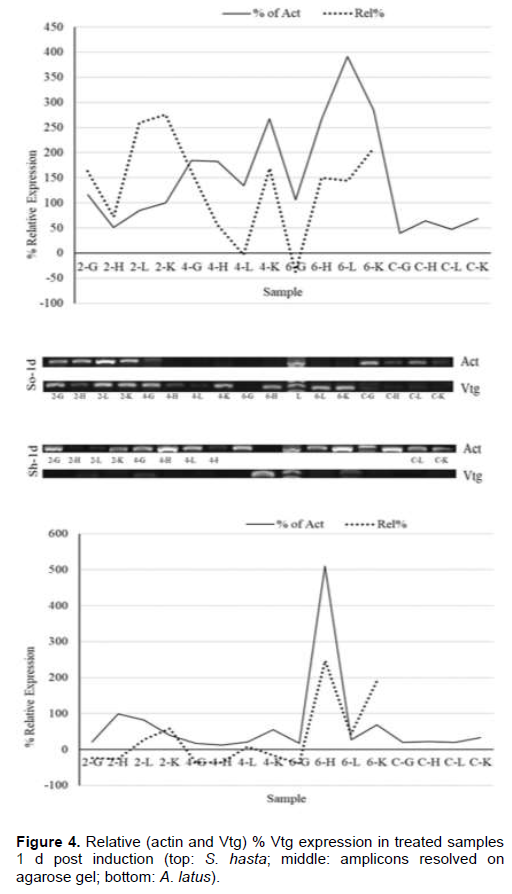
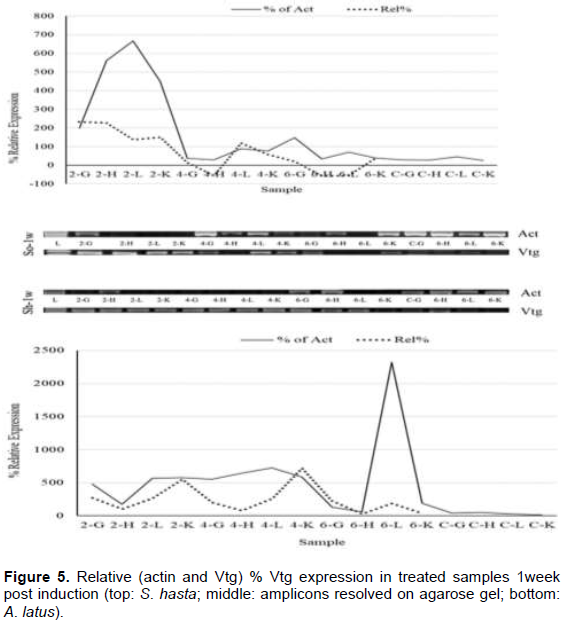
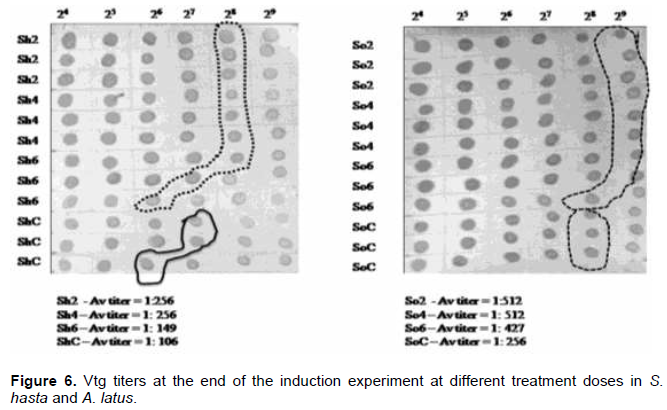
The work of the present investigation formed a part of the project which was aimed at investigating the endocrine disrupting chemicals (EDCs) in the coastal waters of Kuwait (Al-Jandal et al., 2018). The major objective of the present experiment was to trace the influence of estrogen, the major endocrine disrupting chemical (EDC), on the expression of Vtg in different organs of two commercially important protandrous breams. The study was also to partially characterize the Vtg gene in bluefin bream, a commercially important marine fish of the region. The dot-blot assay to evaluate the effect of induction on the Vtg production indicated the highest titers in 2 and 4 µg/fish dose levels. These results agree with the results on Vtg gene expression assays in this study. Even the lowest level of injected dose 2 µg/fish produced elevated gene expression in both the species, S. hasta and A. latus. Similar results were obtained in the case of rainbow fish (Melanotaenia fluviatilis) where minimum level of estradiol required was 3 µg/fish (Shanthanagouda, 2011). Receptor mediated uptake of Vtg by the oocytes after being transported through blood from the liver, the site of synthesis of Vtg, is well-known. Since the estrogen was injected, even doses of 2 µg/fish were enough to induce Vtg production. Male fish do not produce Vtg, however it was established that they respond to exogenous estradiol like that was noticed in oviparous vertebrates (Carnevali et al., 1995) by increased production and secretion of Vtg from the liver (Le Guellec et al., 1988; Bidwell et al., 1991). Most teleost studies showed that the Vtg gene expression is predominantly seen in liver tissue; however, they stated that organs such as gill, fin, heart, kidney, spleen and intestine also show Vtg gene expression at varying intensities (Wang et al., 2005, Yin et al., 2009, Zhou et al., 2009, Ma et al., 2009; Zhong et al., 2014). Enhanced expression of Vtg genes in kidney at 1-week post induction and significant relative (to Vtg levels of un-induced fish) expression in all the tissue examined is probably indicative of immune modulatory function of vitellogenin. Similar views of immune modulatory effects such as anti-viral and anti-bacterial properties of vitellogenin have been reported (Li et al., 2008; Soffker and Tyler, 2012, Zhong et al., 2014). Significant response of both yellowfin and bluefin breams to all doses of 17-β estradiol are like the response recorded in tilapia with a single dose of 5 µg/fish (Davis et al., 2007) of the hormone. The inflow and outflow from sewage treatment plans (STPs) in Kuwait were found to have significant levels of EDCs with estrogens contributing to an extent of 30 and 368 ng/l, respectively (Saeed et al., 2012). The results of the present study indicating high Vtg expressions in liver and other organs have supporting evidence from the earlier works (Al-Jandal et al., 2018) who recorded significant enlargement of liver of fish from the natural exposure experiments where the STP discharges were found to contain considerable levels of EDCs. Thus, it will be interesting to know about the dynamics of immune genes in fish exposed to EDCs for a longer duration and examine their disease susceptibility levels under such conditions. The aggregation or otherwise of fish near the nutrient/EDC-rich discharge sites might provide an opportunity for the EDCs to significantly influence the Vtg and immune gene functions. Future studies aimed at combination exposure of fish to different EDCs, found to occur near the STP discharge point, could provide information on the complex interactions and the immunity status.
ACKNOWLEDGEMENT
We are thankful to the partial funding support (P214-42WE-01) for the study by the Kuwait Foundation for the Advancement of Sciences (KFAS), the approval of the project (EM059C) and support by the Management of Kuwait Institute for Scientiï¬c Research.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Al-Jandal N, Saeed T, Azad IS, Abusam A (2016). Chemical and biological assessment of endocrine disrupting chemicals in wastewater and coastal waters in Kuwait. Project Report, KISR 13929, 108 p. |
|
|
Al-Jandal N, Saeed S, Azad IS, Al-Subiai S, Al-Zekri W, Hussain S, Al-Hasan E (2018). Impact of endocrine disrupting compounds in sewage impacted coastal area on seabream. Ecotoxicology Environmental Safety 150:280-288. |
|
|
Bidwell CA, Kroll KJ, Severud E, Doroshov SI, Carlson DM. (1991). Identification and preliminary characterization of white sturgeon (Acipenser transmontanus) vitellogenin mRNA. General and Comparative Endocrinology 83:415-424. |
|
|
Burki R, Krasnov A, Bettge K, Rexroad CE III, Afanasyev S, Antikainen M, Burkhardt-Holm P, Wahli T, Segner H (2012). Pathogenic infection confounds induction of the estrogenic biomarker vitellogenin in rainbow trout. Environmental Toxicology and Chemistry 31:2318-2323. |
|
|
Carnevali O, Sabbieti MG, Mosconi G, Polzonetti-Magni AM (1995). Multihormonal control of vitellogenin mRNA expression in liver of frog, Rana esculenta. Molecular and Cellular Endocrinology 114:19-25. |
|
|
Chen CTA (2008). Effects of Climate Change on Marine Ecosystems. In: K. Tsukamoto, T. Kawamura, T. Takeuchi, T. D. Beard, Jr. and M. J. Kaiser, eds. Fisheries for Global Welfare and Environment, 5th World Fisheries Congress 2008, pp. 307-316. |
|
|
Dang ZC (2014). Fish biomarkers for regulatory identification of endocrine disrupting chemicals. Environmental Pollution 185:266-270. |
|
|
Dang ZC (2016). Interpretation of fish biomarker data for identification, classification, risk assessment and endocrine disrupting chemicals. Environment International 92-93:422-441. |
|
|
Davis LK, Hiramatsu N, Hiramatsu K, Reading BJ, Matsubara T, Hara A, Sullivan CV, Pierce A L, Hirano T, Garu EG (2007). Induction of there vitellogenins by 17 beta-estradiol with concurrent inhibition of growth-insulin-like growth factor 1 axis in a euryhaline teleost, the tilapia (Oreochromis mossambicus). Biology of Reproduction 77:614-624. |
|
|
Hutchinson TH, Bög C, Winte, MJ, Owens JW (2009). Benefits of the maximum tolerated dose (MTD) and maximum tolerated concentration (MTC) concept in aquatic toxicology. Aquatic Toxicology 91:197-202. |
|
|
Kavlock RJ, Daston GP, DeRosa C, Fenner-Crisp P, Gray LE, Kaattari S, Lucier G, Luster M, Mac MJ, Maczka C, Miller R, Moore J, Rolland R, Scott G, Sheehan DM, Sinks T, Tilson HA (1996). Research needs for the risk assessment of health and environmental effects of endocrine disruptors: a report of the U.S. EPA-sponsored workshop. Environmental health perspectives 104(suppl 4):715-740. |
|
|
Kuhl AJ, Brouwer M (2006). Antiestrogens inhibit Xenoestrogen induced brain aromatase activity but do not prevent xenoestrogen-induced feminization in Japanese medaka (Oryzias latipes). Environmental Health Perspectives 114:500-506. |
|
|
Le Guellec K, Lawless K, Valotaire Y, Kress M, Tenniswood M (1988). Vitellogenin gene expression in male rainbow trout (Salmo gairdneri). General and Comparative Endocrinology 71(3):359-371. |
|
|
Leon-Olea, M, Martyniuk CJ, Orlando EF, Ottinger MA, Rosenfeld CS, Wolstenholme JT, Trudeau VL (2014) Current concepts in neuroendocrine disruption. General and Comparative Endocrinology 203:158-173. |
|
|
Li Z, Zhang S, Liu Q (2008). Vitellogenin functions as a multivalent pattern recognition receptor with an opsonic activity. PLoS One 3(4):e1940. |
|
|
Ma L, Li D, Wang J, He J, Yin Z (2009). Effects of adrenergic agonists on the extrahepatic expression of vitellogenin Ao1 in heart and brain of the Chinese rare minnow (Gobiocypris rarus). Aquatic Toxicology 91(1):19-25. |
|
|
Milla S, Depiereux S, Kestemont P (2011). The effects of estrogenic and androgenic endocrine disruptors on the immune system of fish: a review. Ecotoxicology 20:305-319. |
|
|
Oshima Y, Kang IJ, Kobayashi M, Nakayama K, Imada N, Honjo T (2003). Suppression of sexual behaviour in male Japanese medaka (Oryzias latipes) exposed to 17β-estradiol. Chemosphere 50:429-436. |
|
|
Saeed T, Al-Bloushi A, Abdullah HI, Al-Khabbaz A, Jamal Z (2012). Preliminary assessment of sewage contamination in coastal sediments of Kuwait following a major pumping station failure using fecal sterol as markers. Aquatic Ecosystems and Health 15:25-32. |
|
|
Shanthanagouda AH (2011). Molecular responses of the Murray River rainbowfish, Melanotaenia fluviatilis exposed to endocrine disrupting chemicals. PhD Thesis submitted to School of Applied Sciences Science, Engineering and Technology Portfolio, RMIT University P 228. |
|
|
Soffker M, Tyler C (2012). Endocrine Disrupting Chemicals and Sexual Behaviors in Fish - A Critical Review on Effects and Possible Consequences. Critical Reviews in Toxicology 42(8):653-668. |
|
|
Wang H, Tan JT, Emelyanov A, Korzh V, Gong Z (2005). Hepatic and extrahepatic expression of vitellogenin genes in the zebrafish, Danio rerio. Gene 356:91-100. |
|
|
Wheeler JR, Panter GH, Weltje L, Thorpe KL (2013). Test concentration setting for fish in vivo endocrine screening assays. Chemosphere 92:1067-1076. |
|
|
Yin N, Jin X, He J, Yin Z (2009). Effects of adrenergic agents on the expression of zebrafish (Danio rerio) vitellogenin Ao1. Toxicology and Applied Pharmacology 238(1):20-26. |
|
|
Zhong L, Yuan L, Rao Y, Li Z, Zhang X, Liao T, Dai H (2014). Distribution of vitellogenin in zebrafish (Danio rerio) tissues for biomarker analysis. Aquatic Toxicology 149:1-7. |
|
|
Zhou J, Zhu XS, Cai ZH (2009). Endocrine disruptors: an overview and discussion on issues surrounding their impact on marine animals. Journal of Marine Animals and their Ecology 2:7-17. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


