Full Length Research Paper

ABSTRACT
Three bivalves from the coast of Ghana: Crassostrea tulipa (n = 275), Anadara (Senilia) senilis (n = 310), from two ‘open’ lagoons (Benya and Ningo) and a ‘closed’ lagoon (Sakumo), and Perna perna (n = 155), from rocky shores adjacent to Benya and Sakumo, were analyzed for their total Cu, Zn, Fe, Mn, Cd and Hg concentrations and total body burden/load. The analyses were based on three-size class from two seasons. Average concentrations for oysters (C. tulipa) were: 40, 1197, 477, 16, 0.57 and 0.15 μg/g dw respectively. Cd and Hg levels in dry season samples were higher than those in wet season. While Zn and Fe dry season levels were lower than wet season with no variation in Cu and Mn, average total body burden were: 16, 451, 178, 6.6, 0.25 and 0.11 μg respectively. Geographical variations observed were mostly due to size distribution rather than the ecological differences between stations. Factors observed to influence metal concentrations and accumulation in these organisms include; size, metal bioavailability, season of sampling, hydrodynamics of the environment and reproductive cycle. Element concentrations in bivalves at the same location differ between different species and individuals due to species-specific ability/capacity to regulate or accumulate trace metals. Log-transformed regression analyses between body burden and size were used to compare the species and stations. In addition, the results were compared within class and among classes.
Key words: Trace elements, bivalves, bio-indicators, size class, lagoons, seasonal variation.
INTRODUCTION
The term 'trace element' is used in this text synonymously with 'heavy metal' and includes both the essential (e.g. Cu, Fe, Zn) and non-essential (Cd, Hg, Pb) ones. Elements known to be essential to animal life at trace concentrations are: I, Mo, Se, Cr, Sn Ni, Fe, Zn, Cu, Mn, Co, F, Si, and V. A further trace element, boron, is essential for plant life, in addition to the above. Not only these but also, 20 to 30 other elements occur, more or less constantly in low but variable concentrations in living tissues. The presence of these elements is believed to reflect the contact of the organism with its environment. Elements such as: Sn, V, Ni, Si and F, often referred to as the 'newer trace elements', only emerged as essential recently as an outcome of the use of purified diet and plastic isolator techniques (Otchere, 2019). A few trace elements are referred to as toxic because they are toxic at relatively low concentrations (close to the ppt level or ng/l; e.g. Pb EC50 is 370 µg/l (1.79 µmol./l)); however, this statement can be misleading because all the trace elements are potentially toxic if ingested at sufficiently high levels and for long enough periods. With some, notably F in man and Cu in sheep, the margins between beneficial and toxic intakes, that is, between needs and tolerance are small. With others such as Zn and Mn, tolerance is high Gand wide margins exist between minimum essential intake and those resulting in toxic effects. Concentrations of heavy metals in aquatic environments have two phases: dissolved and particulate. The comparison of metal levels between different years in the same environment or between different environments should be considered very carefully, taking into accounts the seasonal variability which can be very important in estuaries. In these environments concentrations may depend very much on salinity, apart from the seasonal variability (Laporte et al., 1997; Baeyens, 1998; Otchere, 2019).
Monitoring programs and research for metals in the environment have become widely established because of concerns over accumulation and toxic effects, particularly in aquatic organisms and to humans consuming these organisms. The criteria by which organisms are accepted as biological indicators for the assessment of contamination were proposed more than fifty years ago and remain unchanged (Phillips, 1990; Vaisman et al., 2005; Otchere, 2019). Bivalves are widely used as bio-indicators of heavy metal pollution in coastal areas because they are known to accumulate these elements, providing a time integrated indication of environmental contamination. In comparison to fish and crustaceans, bivalves have a very low level of activity of enzyme systems capable of metabolizing persistent organic pollutants (POPs), such as aromatic hydrocarbons and polychlorinated biphenyls. Therefore, contaminants concentrations in the tissues of bivalves more accurately reflect the magnitude of environmental contamination (Boyden and Phillips, 1981; Reinfelder et al., 1997; Otchere, 2005). Factors known to influence metal concentrations and accumulation in these organisms include; metal bioavailability, season of sampling, hydrodynamics of the environment, size, sex, changes in tissue composition and reproductive cycle. Seasonal variations have been related to a great extent to seasonal changes in flesh weight during the development of gonadic tissues. Element concentrations in mollusks at the same location differ between species and individuals due to species-specific ability/capacity to regulate or accumulate trace metals, hence the question: Do species matter in monitoring programs? (Joiris et al., 2000; Silva et al., 2003; Otchere, 2005).
This paper presents data on the total metal concentration, body burden based on size class for six elements and the effect of season on these bivalves. In addition, it also highlights that different animals in the same community at the same trophic level could accumulate pollutants differently due to differences in animal physiology, habitat/niche's physical and chemical properties.
MATERIALS AND METHODS
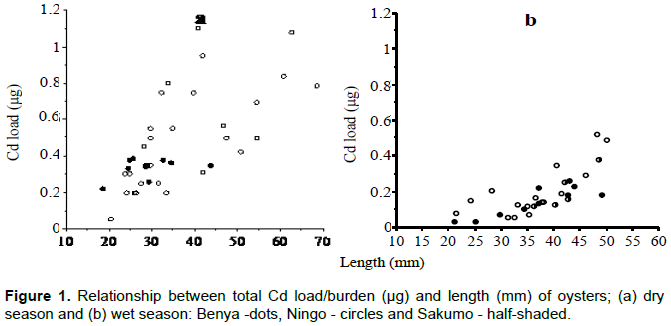
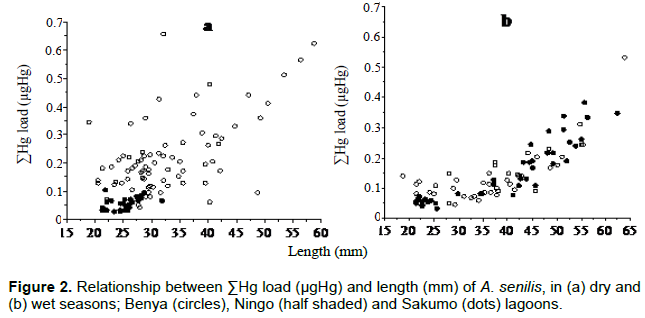
Three species of bivalves were collected from the coast of Ghana: with approximately 590 km long coastline, which stretches from 3°W to 1° 10'E and lies between 4°5' and 6°6'N. About 50 lagoons occur on the coast. The lagoons are of two main types: 'open' and 'closed' lagoons. The open lagoons are in contact with the sea throughout the year and therefore partly under tidal influence. Temperature and salinity ranges are: 24 to 32°C and 10 to 40 psu, respectively. The closed lagoons are cut off from the adjacent sea by a sand bar (about 40 m wide) for the greater part of the year. Temperature and salinity ranges are: 27 to 34°C and 27 to 70 psu, respectively. Hyperhaline condition results from evaporation during the dry season (Yankson and Obodai, 1999; Otchere, 2020). Samples of Crassostrea tulipa (oysters; n = 275), Anadara (Senilia) senilis (cockles; n = 310) from three lagoons and Perna perna (mussels; n = 155) from two rocky shores adjacent to these lagoons were picked. Samples were randomly collected in October 2016 and February 2017 (wet and dry seasons, respectively). The analytical procedure used to measure the metals: Cd, Cu, Fe, Mn and Zn in the bivalves was based on UNEP/FAO/IAEA (1982) with modification as follows: sub-sample (dried) tissue (0.2 g) was heated with 10 ml of concentrated nitric acid (70 to 90°C) till all tissue had been digested. The temperature was then gradually increased to 135°C and drops of H2O2 added for further oxidation. After cooling, solutions were diluted to 50 ml with double distilled water and filtered with 1.6 μm fiberglass filter paper (GF/A). Samples were then stored at room temperature in 50-ml volumetric flask until they were analyzed. Analyses were carried out with flame Atomic Absorption Spectrophotometer (AAS) (Perkin Elmer 3110 system 2) with electrode discharge lamps (EDL) and hollow cathode lamp (HCL). Wavelengths, detection limits and quality control/assurance have been described in detail by Otchere (2003). Method for the determination of Hg has been described in Joiris et al. (1998). The size classes were based on shell length (less than 30 mm as small, 30-40 mm as medium and greater than 40 mm as large) for cockles and mussels; while shell length of (less than 40 mm as small, 40-50 mm as medium and greater than 50 mm as large) for oysters. The data were expressed as total body concentration and total body burden. The results were compared within class (from different locations and species) and among classes using Mann-Whitney-U and Kruskal-Wallis test of significance as the distributions were non-normal and skewed. The species were compared using log transformed regression analysis. Significance levels were fixed at 0.05 as significant and 0.01 as highly significant; also, median values (min - max) were presented.
RESULTS
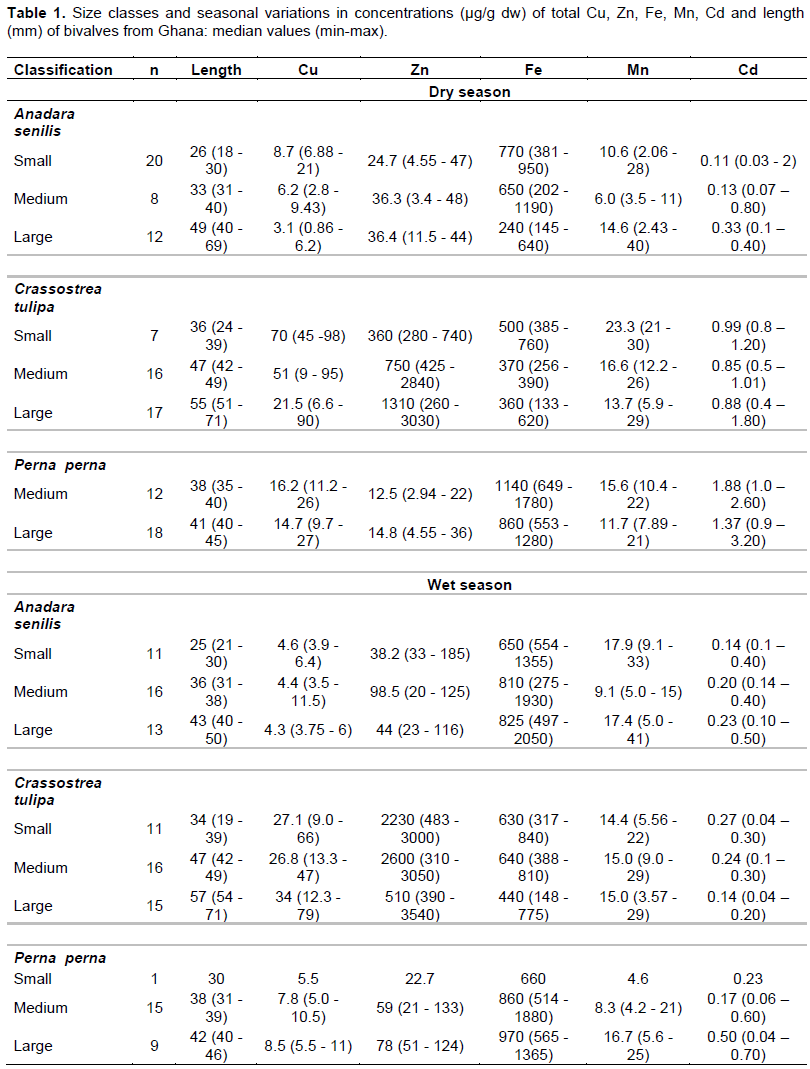
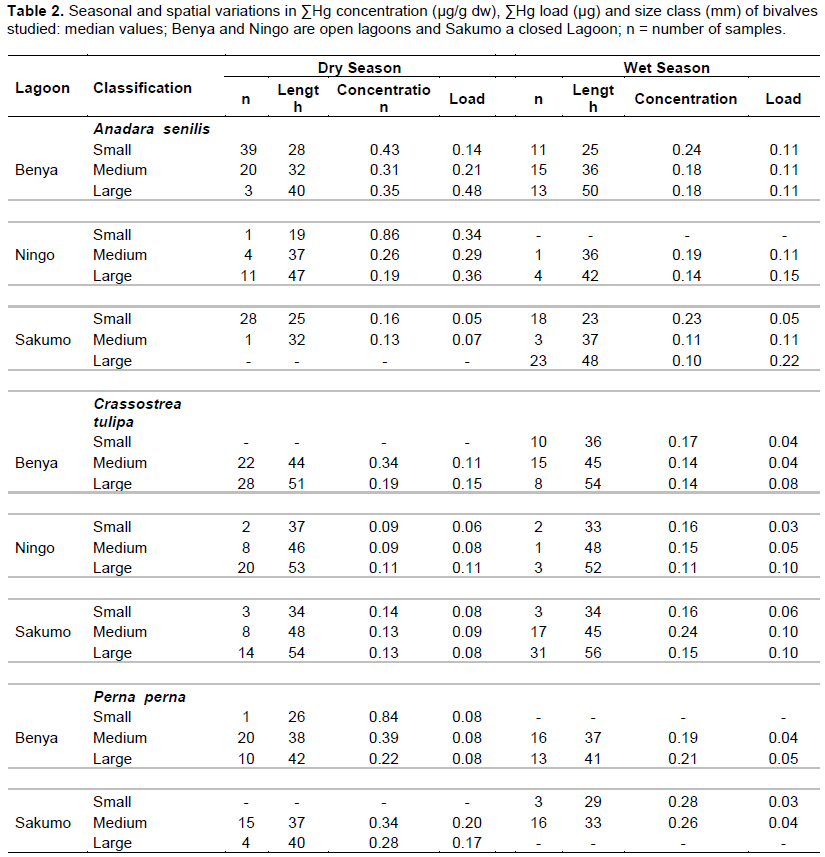
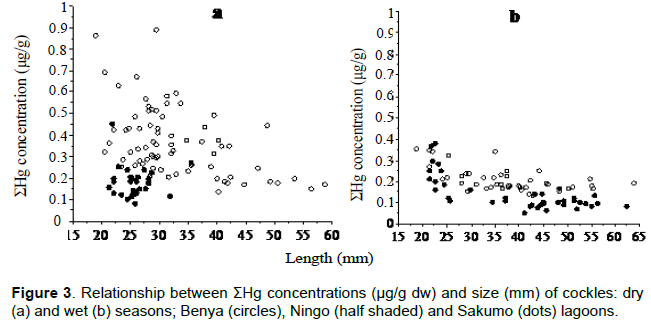
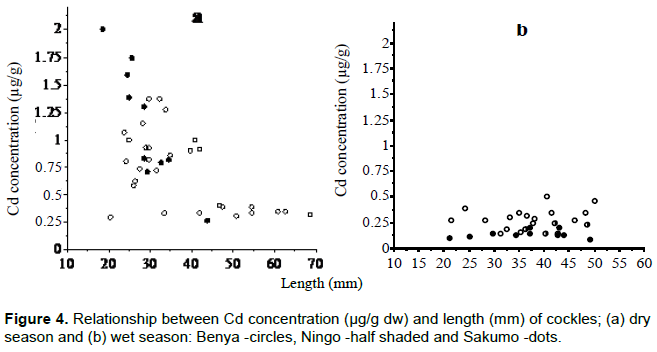
Size class values for body concentrations and body burden (load) for both seasons are shown in Table 1 for Cd, Cu, Fe, Mn, Zn and Table 2 for Hg. One important factor influencing these results is size. For example, Hg concentration in the dry season (cockles) decreases with size as a "growth dilution" effect but stable in the wet season (Figure 3). In this case, regional differences in Hg could be entirely due to size effect. Likewise, the seasonal difference in Cd (Figure 4) and Cu dry season concentrations were also apparently higher than those in the wet season. A closer look at the total body burden data with respect to Hg (Table 2), reveals that seasonal differences could be detected. Comparing Benya and Sakumo wet season body burden (with larger cockle sizes; 37 mm) to dry season (29 mm), suggest a lower metal availability: Cd (Figure 1) and Hg (Figure 2) at the stations during the wet season. Thus, the above seasonal differences in Hg and Cd concentrations (Figures 3 and 4) are real. Both body burden and concentrations in these metals showed no geographical difference during the seasons and any seeming difference was due to size/age, whereas Zn, Fe and Mn dry season concentrations were lower than wet season. Again considering the body burden for Hg, the influence of size is obvious from Table 2; and Mn did not show any difference both in location and season. Any detected differences in concentration were apparent. On the other hand, seasonal differences could be found in Fe and Zn concentrations. For example, Ningo dry season burden (with higher biomass) was lower than wet season, an indication of a higher contamination of these metals in the wet season. Geographical differences could be detected in other species, such as higher Zn concentration in oysters from Benya during both seasons. Seasonal variations on the other hand, could be detected mainly due to hydrological differences like the effect of rain or differences between open and closed lagoons. Elevated levels in Table 1: Zn in oysters (260 - 3540 μg/g dw), Fe in cockles (150- 2050 μg/g dw) and in mussels (510 - 1880 μg/g dw) reflect the inherent variability of metals in bivalves dependence on the particular species-metal pair considered, and also on the degree of contamination involved (Otchere, 2003).
DISCUSSION
Where it is difficult to assess whether observed differences in tissue level/burden between these bivalves reflect real differences in environmental trace metal constitution, or are merely due to variations in body size; plots of μg metal per individual (total burden versus size) could be carried out. Generally, the total metal burden is related to body weight/size by the metabolic power function: Y = aWb, where Y is the burden, W the dry weight, a the intercept and b the slope co-efficient (Otchere, 2019). Coefficients greater than two (b > 2) of the body weight as shown by Cd, Zn and Mn in cockles and Cu, Fe, Cd and Mn in mussels (Table 3) can best be explained as being due to removal of these elements from body circulation and accumulated within specific tissues, possibly as a result of some exceptional affinity. In the case of the oysters (Table 3), all the relationships were approximately equal to one (b = 1), implying that element concentrations were independent of size. The absolute amount of the metal is not dictated by the amount of such binding compound/storage mechanisms in the tissues.
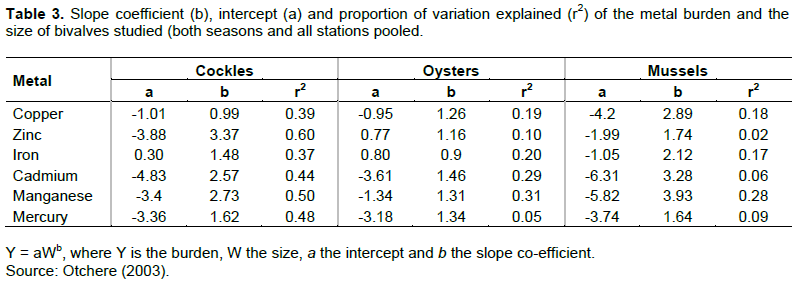
The tendency to decrease the metal concentrations with an increase in body weight is not significant in all cases. However, for the essential elements; Cu and Fe (in cockles and oysters), the size of the animal could be mainly responsible for the metal concentration variability. The slope value of about 2 in cockle populations examined: Benya, Ningo and Sakumo (Table 4) may well be due to the unusually elevated environmental Fe concentration in the lagoons. An increase in metal burden with size/weight of the individuals is expected and it is indeed noted for all metals, except for Zn and Cd in mussels. Information about the factors responsible for the observed metal burden-size can be obtained from the slope of the regression lines (Table 4).
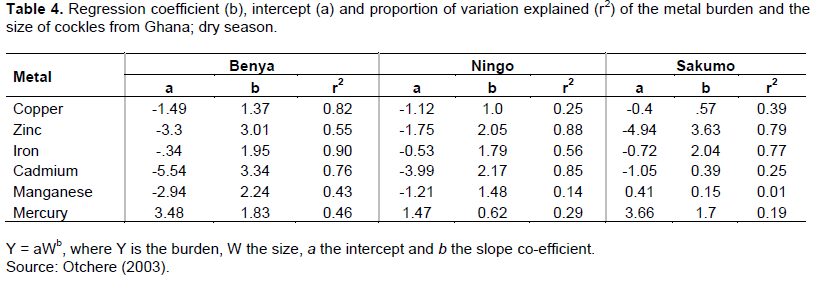
The changes in metabolic rates of bivalves with age/size and season as well as the variation in bioavailability of metals in the surrounding environment with time might be responsible for these variations in these mollusks (Fowler et al., 1993; Otchere, 2003). Metal availability probably fluctuated with salinity in these lagoons during the seasonal cycle (Phillips, 1990; Cossa and Rondeau, 1985). This probably caused the higher metal burden and concentration in the wet season (such as for Fe and Zn). Higher wet season levels of Fe and Zn might as well be due to 'import' as most roofing in Ghana are made of galvanized iron sheets. Lower wet season levels for Cd and Hg could be attributed to loss through spawning or washed-out of the lagoons during the rainy period. Many metals are also found in agricultural products. Those present in fertilizers include Cd, Cu, Cr, Ni, Mn, Mo and Zn. Eventually, many of these metals may accumulate in soils and become exposed to run-offs during the rainy season. Biological variables such as size, sex or changes in tissue composition and reproductive cycle as well as the season of sampling and the hydrodynamics of the lagoons have to be considered. Seasonal variations have been reported to be higher in winter/dry than in summer/wet season. These seasonal variations have been related to a great extent to seasonal changes in flesh weight during development of gonadic tissues (Fowler et al., 1993; Joiris et al., 2000; Otchere, 2019). Metabolic changes in mollusk with age and season, storage mechanisms as well as temporal variations in metal bioavailability in the surrounding environment might be reasons to explain these metal burden-size relationships in the lagoons (Tables 3 and 4).
Considering Cd concentration in cockles in the wet and dry seasons, the slope of the Cd/size relationship results from a combination of several factors which act separately or simultaneously. The figures reflect the competition between the opposite effect of age and growth on the accumulation of Cd. The metal accumulates with age whereas the weight gain of the individual has the effect of reducing the metal concentration. The relationship observed underlying the linear relation could effectively correspond in a given range of soft tissue weights to the presence of individuals of the same age class whose soft tissue weight differs considerably. In this way, while the Cd burden would increase with age (Figure 1), concentration would diminish with the weight/size (Figure 4) within each age class if growth is more rapid than the accumulation rate.
To the inverse effect of age and soft tissue weight is added the effect of change in the bioavailability of Cd in the environment. To demonstrate these mechanisms clearly, the phenomenon in the light of situations encountered in cockles will be restated: that in bioaccumulation in bivalves, the uptake of the metal is higher in the smaller individual than in the larger ones, generally resulting in “growth dilution” effect. The result is a negative slope for metal/size concentration relationships in the case of Hg (Figure 3) but not in Cd as in Figure 4 (for the non-essential metals), which was only due to seasonal effect. Higher metabolism in juvenile individuals can cause a reduction in metal concentrations in soft tissue, because the tissue grows more quickly than the metal can be absorbed. This mechanism can explain seasonal variations recorded in Cd concentrations (Cossa et al., 1980; Fowler et al., 1993).
Applying the same principle as in Cd to Zn and Cu would explain the variations observed in Table 1. For example, while load (body burden) was higher in dry season, Cu expresses the opposite effect on concentration to explain the temporal difference observed. Comparative differences between wet and dry seasons for Cu and Zn were about 1.3-fold and 3-fold increase respectively. The difference was not significant in Cu because as an essential element, it is strictly regulated. Contrarily, Zn showed a significant seasonal variation due to the elevated level in the Ghana environment as a result of galvanized roofing sheet used, thus Zn becomes more available during the rainy period than the dry season. Therefore, Zn regulation in cockles could be described to have broken down due to elevated levels in the environment. In such situation, cockles cannot be used as a bio-indicator for Zn. It is clear that concentrations of all three metals were mainly determined by cockle condition and seasonal fluctuations in metal concentrations, which in turn depend to a large extent on the magnitude and direction of temporal alteration in soft tissue dry weight of the cockles (Otchere et al., 2003).
Boyden and Phillips (1981) studying Crassostrea gigas concluded that temporal variations in metal concentrations were mainly caused by changes in soft tissue weights of the bivalves according to the sexual cycle. Thus, alterations in the concentrations of metals reciprocated those of the whole soft tissue weights. Investigations of element seasonality in bivalves are important in terms of their implications for the use of these organisms as biological indicators of metal abundance. The present study suggests that element concentrations may commonly vary in A. senilis by a factor of two or more during the annual cycle. Clearly, such variation is a potential disruptive factor in the use of A. senilis to monitor metal abundance at different locations and seasons. The simultaneous sampling of cockles at all sites in such an indicator survey might be considered sufficient to overcome this source of variations; but even in this case, there can be no guarantee that cockles from each locations are of similar condition. The best method for diminishing the effects of seasonality and species inherent physiology is therefore to time sampling programs to occur during a period of relatively little change in element concentrations. In addition, some other effects of season could be eliminated by the removal of gonads prior to analysis.
CONCLUSION
The four essential elements which were studied, were present in similar respective concentrations to those found for other bivalves elsewhere and exhibited similar seasonal pattern in terms of their concentrations, although of different magnitudes. There was no influence of season and location on Mn and Cu concentrations, but location played an important role in Fe and Zn concentrations while season exhibited a moderate influence. The wet season maxima in Zn, Fe and Mn observed should reflect a higher metal availability during this season (through ‘import’). Temporal variations in metal concentrations were mainly caused by changes in soft tissue weights of the bivalves according to the sexual cycle. Element concentrations in mollusks at the same location differ between different species and individuals due to species-specific ability/capacity to regulate or accumulate trace metals. Hence, in monitoring programs, the type of species matter due to the fact that specific species have inherent physiological capacity and ability to regulate or accumulate specific trace elements in the environment. Seasonal and regional variations observed in this study were due to these factors mentioned above. For example, the present study suggests that element concentrations may commonly vary in A. senilis by a factor of two or more during the annual cycle. Clearly, such variation is a potential disruptive factor in the use of A. senilis to monitor metal abundance at different locations and seasons. Results on metal contamination in bivalves should therefore not be used directly as reflecting environmental contamination; they should first be normalized for these physical and/or environmental factors (season of sampling, hydrodynamics of the water and the position of the mollusk at the shore) before any spatial and/or temporal comparison is made.
CONFLICT OF INTERESTS
The author has not declared any conflict of interests.
REFERENCES
|
Baeyens W (1998). Evolution of trace metal concentrations in the Scheldt estuary (1978-1995). A comparison with estuarine and ocean levels. Hydrobiologia 366(1):157-167. |
|
|
Boyden CR, Phillips DJH (1981). Seasonal variation and inherent variability of trace elements in oysters and their implications for indicator studies. Marine Ecology Program Series 5:29-40. |
|
|
Cossa D, Rondeau JG (1985). Seasonal, geographical and size induced variability in mercury content of Mytilus edulis in an estuarine environment: a re-assessment of mercury pollution level in the Estuary and Gulf of St. Lawrence. Marine Biology 88(1):43-49. |
|
|
Cossa D, Bourget E, Pouliot D, Chamut JP (1980). Geographical and size variations in the relationship between trace metal content and body weight in Mytilus edulis. Marine Biology 58(1):7-14. |
|
|
Fowler SW (1990). Critical review of selected heavy metals and chlorinated hydrocarbon concentrations in the marine environment. Marine Environmental Research 29(1):1-64. |
|
|
Fowler SW, Readman JW, Oregioni B, Villeneuve J-P, McKay K (1993). Petroleum hydrocarbons and trace metals in Nearshow Gulf sediments and biota before and after the 1991 War: An assessment of temporal and spatial trends. Marine Pollution Bulletin 27:171-182. |
|
|
Joiris CR, Azokwu MI, Otchere FA, Ali IB (1998). Mercury in the bivalve Anadara (Senilia) senilis from Ghana and Nigeria. The Science of Total Environment 224(1-3):181-188. |
|
|
Joiris CR, Holsbeek L, Otchere FA (2000). Mercury in the bivalves Crassostrea tulipa and Perna perna from Ghana. Marine Pollution Bulletin 38(7):618-622. |
|
|
Laporte JM, Truchot JP, Ribeyre F, Boudou A (1997). Combined effect of water pH and salinity on the bioaccumulation of inorganic mercury and methylmercury in the shore crab Carcinus maenas. Marine Pollution Bulletin 33(11):880-893. |
|
|
Otchere FA (2003). Heavy metals concentration and burden in the bivalves (Anadara (S.) senilis, Perna perna and Crassostrea tulipa) from lagoons of Ghana: model to describe mechanism of accumulation /excretion. African Journal of Biotechnology 2(9):280-287. |
|
|
Otchere FA, Joiris CR, Holsbeek L (2003). Mercury in the bivalves Anadara (Senilis) senilis, Perna perna and Crassostrea tulipa from Ghana. The Science Total Environment 304 (1-3):369-375. |
|
|
Otchere FA (2005). Organochlorines (PCBs and Pesticides) in the bivalves: Anadara (Senilis) senilis, Perna perna and Crassostrea tulipa from the lagoons of Ghana. The Science of Total Environment 348(1-3):102-114. |
|
|
Otchere FA (2019). A 50-year review on heavy metal pollution in the environment: Bivalves as bio-monitors. African Journal of Environmental Science and Technology 13(6):220-227. |
|
|
Otchere FA (2020). Population Patterns, Biology and Ecology of Bivalves from Lagoons and Rocky Shores in Ghana. International Journal of Research - Granthaalayah 8(4):53-66. |
|
|
Phillips DJH (1990). In: Furness RW and Rainbow PS (eds) Heavy metals in the marine environment. CRC Press, Florida pp. 81-99. |
|
|
Reinfelder JR, Wang W-X, Luoma SN, Fisher NS (1997). Assimilation efficiencies and turnover rate of trace elements in marine bivalves: a comparison of oysters, clams and mussels. Marine Biology 129(3):443-452. |
|
|
Silva CAR, Rainbow PS, Smith BD (2003) Biomonitoring of trace metal contamination in mangrove-lined Brazilian coastal systems using the oyster Crassostrea rhizophorae: comparative study of regions affected by oil, salt pond and shrimp farming activities. Hydrobiologia 501(1):199-206. |
|
|
UNEP/FAO/IAEA (1982). Determination of total Cadmium, Zinc, Lead and Copper in selected marine organisms by Atomic Absorption Spectrometry. Reference methods for marine pollution studies No. 11. United Nations Environmental Programme. |
|
|
Vaisman AG, Marins RV, Lacerda LD (2005) Characterization of the mangrove oyster, Crassostrea rhizophorae, as a bio monitor for mercury in tropical estuarine systems northeast Brazil. Bulletin of Environmental Contamination and Toxicology 74(3):582-588. |
|
|
Yankson K, Obodai EA (1999). An update of the number, types, and distribution of coastal lagoons in Ghana. Journal of the Ghana Science Association (Special Edition) 2(2):26-31. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


