ABSTRACT
Anthropogenic disturbances (cultivation, harvesting of poles and firewood and protection) play a pivotal role in the recovery and growth of Baikiaea-Guibourtia-Pterocarpus woodland species. The aim of this study is to determine the reliability of tree growth rings in age determination and average radial growth of re-growing woodland stands. The relationship between stem diameter, number of growth rings, and age of selected stands was also investigated. The research assisted in understanding the growth patterns of the key commercial timber species (Baikiaea plurijuga, Guibourtia coleosperma and Pterocarpus angolensis) of the seasonally dry Baikiaea-Guibourtia-Pterocarpus woodlands around Gwaai and Tsholotsho in north-western Zimbabwe. Tree rings were physically counted on basal stem cross-sections collected from 20 trees of different age, for each species in each disturbance regime. STATISTICA statistical package version 7.0 (StaSoft inc, 1984 - 2006) was used for data analysis. The Simple Regression Model was used to test for relationships. The relationship between age and growth rings, showed a strong correlation (p<0.0001). Diameter and number of growth rings had a poor correlation. This showed that growth rings and not diameter can be used to determine the age of the three key timber species. Mean annual ring width was significantly different between species within the same disturbance category (p<0.005) within a specific stand age. Mean radial growth was highest in abandoned crop fields, compared to pole and firewood collection sites. It was therefore concluded that forest managers need to adopt disturbance regimes that prompt optimal mean radial growth of at least key species in the woodlands.
Key words: Disturbance, mean radial growth, tree rings, Baikiaea plurijuga, Guibourtia coleosperma; Pterocarpus angolensis.
Information on growth rates is key in determining the sustainability of harvesting systems and the formulation of sound forest management systems (Caetano et al., 2019; Khai et al., 2020) in natural forest and woodland systems. The relationship between stem diameter and stand age of key tree species and the variation in mean radial growth between different stand conditions should guide good silvicultural management. Several authors have reported that attempts at plantation cultivation of indigenous species have not been successful, for example Pterocarpus angolensis (DC) (Boaler, 1966; Van Daalen et al., 1991, Moses, 2013), Baikiaea. plurijuga (Harms) (Piearce, 1993) and Guibourtia coleosperma (Benth.) J. Leonard (Lemmens et al., 2008; Heita, 2019), making it difficult to estimate their age, mean annual increment (MAI), productivity and other growth parameters (Montoro et al., 2017). How then can forest managers determine the age and growth rates of indigenous tree species when it is difficult to raise them in plantations? However, some tropical and sub-tropical tree species can produce growth rings which correlate with age (Fahn et al., 1981; Gourlay and Barnes, 1994; Grundy, 1995; Stahle et al., 1999; Geldenhuys, 2005; Ngoma et al., 2017). Annual growth rings have been used in the past by forest managers in determining the age of Miombo woodlands in Zambia (Fanshawe, 1956; Syampungani et al., 2010) and in Zimbabwe (Grundy, 1995; Stahle et al., 1999). Grundy (1995) did a 4-year study on stems of unknown management history. Syampungani et al. (2010) showed that ring counts can be used in age determination, with a strong correlation between growth rings, known stand age, and stem diameter, for three Miombo woodland species. B. plurijuga was studied in Zambia by Miller (1952) about the determination of age and rotations and by Ngoma et al. (2017) in terms of dendrochronological potential. Both Miller (1952) and Ngoma et al. (2017) highlighted that B. plurijuga shows clear annual growth rings. Stahle et al. (1999) studied the correlation between the growth rings in P. angolensis and seasonal climatic data, using evidence from phenology, ring anatomy and cross-dating. However, they did not analyze the relationship between the number of growth rings, age of the study site and stem diameter.
There is need to understand the response of key species in terms of mean radial growth under different disturbance factors. Montoro et al. (2017) emphasized the need to consider the best treatment (disturbance) for each species according to the ecological requirements in order to maximize radial growth of the residual trees and enhance the species’ economic value. Therefore, forest managers need to adopt land use practices in line with woodland disturbance regimes and recovery potential to ensure prompt, adequate regeneration, and fast growth of key species. Tree rings and growth rate data hold key information for the development of sustainable forest management schemes, as they give indications on the time required to replace harvested trees. Such data are very useful when assessing species potential and sustainable timber exploitation (Caetano et al. 2019). The question to be answered by this study is “can we use growth rings, stem diameter or both, in age determination for the key species of Baikiaea-Guibourtia-Pterocarpus woodlands?”
The main objective of this study is to determine how reliable is the use of growth rings or stem diameter or both, in age determination, that is, what the relationship is between the number of growth rings, stem diameter and tree age of selected sites. The following research questions guided data collection and analysis: a) Can the individual growth rings be reliably differentiated on cut stems of key tree species in the woodlands? b) Is the number of growth rings the same in larger and smaller stems in a stand of known age? c) Can growth rings be more clearly differentiated in free-growing trees than in suppressed trees? d)Is there a relationship between the number of growth rings, stem diameter and stand age of a selected site, and can such a relationship be used for reliable stand age determination? and e) How does the mean radial growth of key tree species vary in different disturbance factors?
Description of study area
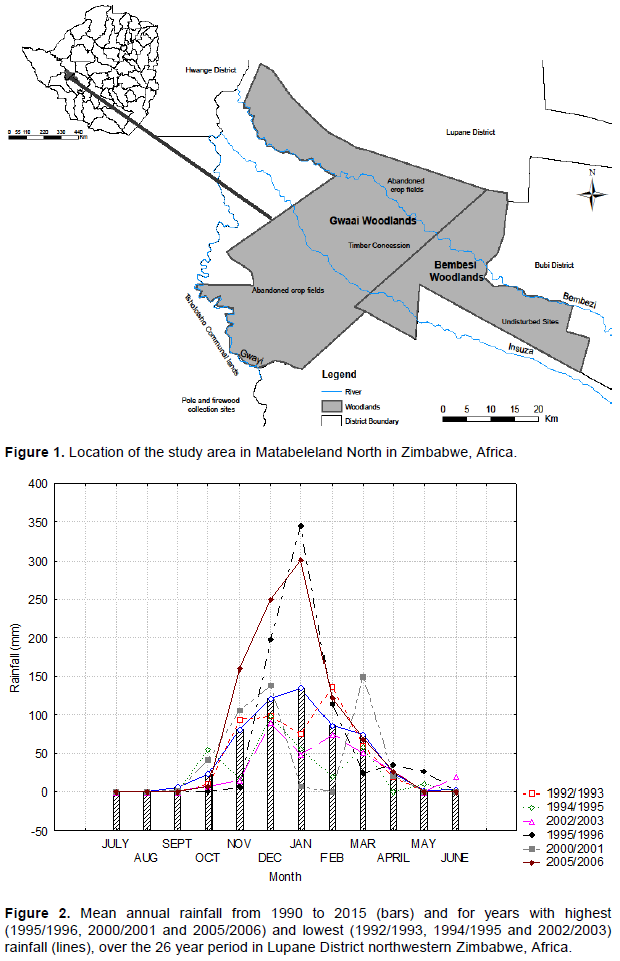
The study was conducted in the Gwaai and Tsholotsho indigenous Baikiaea-Guibourtia-Pterocarpus woodlands of north-western Zimbabwe (Figure 1). Gwaai forest (19°16¹20” S and 27°56¹36” E) and Tsholotsho (19°46¹00” S and 27° 45¹ 00” E) (JAFTA and Forestry Commission, 2001) are both located in the Matabeleland North Province at an altitude ranging between 1010 and 1055 m. Kalahari sands (uniform, both physically and chemically) cover the bulk of the study area. They belong to the regosol group in the amorphic soil order (Nyamapfene, 1991, as cited by Gambiza 2001). The underlying geology is of sedimentary rocks overlying Karoo basalt and sedimentary deposits (JAFTA and Forestry Commission 2001).
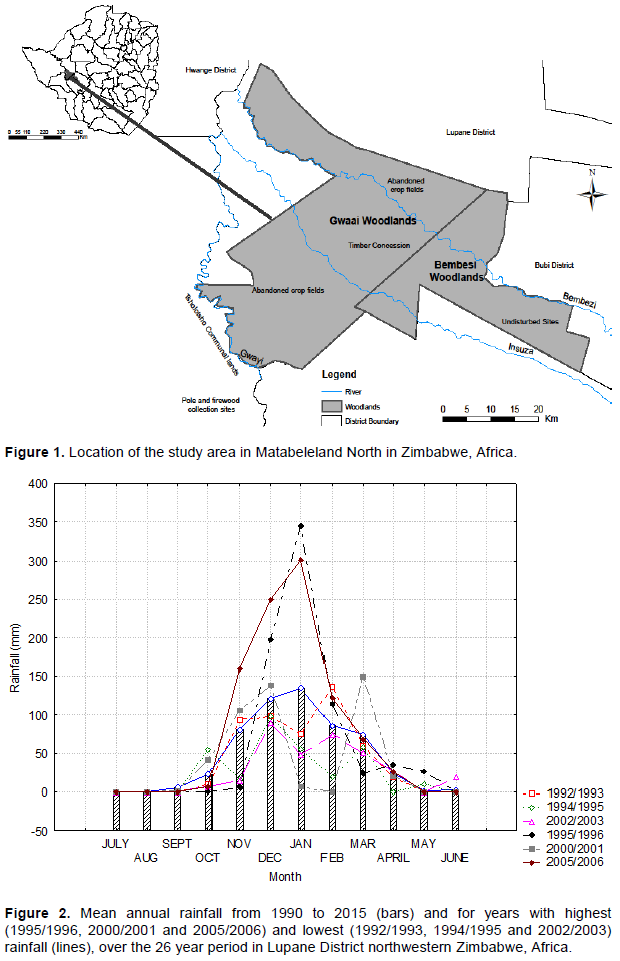
The area experienced much annual fluctuation around the average monthly rainfall over a 26 year period (bars in Figure 2). The highest rainfall was recorded in 2005/2006 followed by year 2000/2001 and 1995/1996 and lowest rainfall was recorded during the drought years of 1994/1995, 2002/2003 and 1992/1993. The short and erratic wet season is usually characterised by dry spells and sporadic droughts (Nemarundwe and Mbedzi, 1999). Mean annual temperature is 21.5°C, with mean monthly temperature ranging from 15°C (June to September) to 25°C (October to December) (Nyamapfene 1991; JAFTA and Forestry Commission 2001). Ground frosts are experienced especially in the valleys in most years between May and September (JAFTA and Forestry Commission 2001).
The area is characterised by six main vegetation types (JAFTA and Forestry Commission 2001): (i) Baikiaea-Guibourtia- Pterocarpus (Fabaceae) woodland occurs on the Kalahari sands;’ (ii) Brachystegia (Fabaceae) woodland occurs along the upper Bembesi river on shallower soils with more silt; (iii) Colophospermum mopane (Kirk ex Benth.) J Leonard woodland, dominated by either stunted or multi-stemmed C. mopane trees, occurs along river valleys on poorly drained and highly erodible alluvial soils; (iv) Vleis (a grassy or marshy wetland, mostly covered by water during the rainy season) are dominated by a single layer of grasses, with isolated trees occurring along vlei fringes; (v) P. angolensis belts growing in association with Burkea africana (Hook) occur as localised stands inside the BaikiaeaGuibourtia-Pterocarpus woodland; and (vi) G. coleosperma woodland dominated by G. coleosperma and occurs on the Kalahari sands.
Land use systems
In Tsholotsho communal areas, people harvest trees on an open-access regime for subsistence and commercial purposes (Matose 2002). Trees of different sizes are cut for various purposes (fuel wood, construction, fences, curios, etc.). Large canopy gaps are created when groups of trees are removed in the pole and firewood collection sites. Farmers grow maize, sorghum, and other crops on the agricultural fields. The agricultural fields are abandoned after many years (40 years or less) of cultivation. Fields are abandoned when they are old and are producing fewer yields (Mutsiwegota and Mudekwe, 1998). It has been noticed that trees of different species will start growing on the abandoned fields from seed or through sprouting from rootstocks.
Data collection
The data were collected in areas of known age after crop cultivation and pole and firewood collection had been terminated and in undisturbed woodland (unknown age). The following ages (8, 17, and 25 years) were selected for study in the abandoned crop fields and pole and firewood collection sites. These were based on the last date since disturbance cessation (that is, 2008, 1999 and 1992 respectively). The information on stand age was obtained from the local communities around the study areas and from Forestry Commission records. Plots were sampled in the undisturbed sites to have stems of similar height as in the development stages of the disturbed sites (Stage 1 height was <2 m; Stage 2 height was 2.1-5 m; Stage 3 height was 5.1-8 m; and Stage 4 height was >8 m). In each site of specific age or height, 20 trees per selected species (B. plurijuga, G. coleosperma and P. angolensis) were selected for sampling. The sampling approach of fixed length - variable width of Walker (1976) was adopted. The plots had a fixed length of 50 m; sampling would stop (at any width) when the required number of trees for the three species was reached, that is, 60 stems (20 stems per species) per site of specific age, giving a total of 180 stems for each disturbance factor, and an overall total of 540 stem sections. A Global Positioning System (GPS) was used to record the position of the plots in the field. A study by Chichinye et al. (2019) showed little variation in environmental factors (such as edaphic factors (soil texture, soil depth, soil nutrients), slope, aspect among others, across the studied disturbance factors hence data on edaphic factors was not collected. The following general information was recorded at each site.
(i) Study area, Study site name, Recorder name and Date
(ii) GPS coordinates: Latitude °S, Longitude °E (X and Y coordinates).
(iii) Condition of surrounding natural vegetation (the appropriate answer was selected):
(a) Height (m): 1 = <2; 2 = 2 - 5; 3 = 5 - 10; 4 = 10 - 20; 5 ≥ 20
(b) Stand cover: 1 = 0 - 30%; 2 = 30 - 50%; 3 >50%.
(c) Date when cutting or cropping was abandoned (to determine possible stand age)
(iv) Relevant information for the site that would be useful for the interpretation of the growth ring development.
The assumption was that the Baikiaea-Guibourtia-Pterocarpus woodland species are light-demanding. That means that a stand that developed after cultivation or harvesting of poles and firewood, would be composed of even-aged trees of different species. Smaller and larger stems would have the same number of rings and hence the same age. Trees with smaller stems would be suppressed trees. Each tree develops growth rings, with each ring consisting of a broader part of lighter wood (faster growth, possibly during the rainy period) and a narrower part of darker wood (slow to no growth, possibly during the dry period).
Single-stemmed trees with a normal, well-formed stem and with no stem defects in the lower 1 m of the stem were selected for sampling. For each selected tree, data on the following were recorded before the tree was cut and after it had been cut:
(i) Tree number (to accompany the collected stem section)
(ii) Species
(iii) Diameter at breast height (DBH, in cm) at 1.3 m above ground level
(iv) Tree height (m)
(v) Diameter (cm) for stem section (DS)
(vi) Number of visible annual growth rings for section at ground level (RS).
(vii) Bark thickness (mm) on two opposite sides of the stem section.
A horizontal smooth cut was made at the bottom of each selected stem to produce 1 cm thick discs. The stems were cut close to ground level. Site name, species code, tree number, DBH and section diameter were recorded on the backside of each disc. All discs dried under tree shade and the unmarked section was smoothened, using a belt-sanding machine. The machine used coarse sandpaper (Figure 3).
Three lines (radii) were drawn on the smoothened surface of each stem section, from the core to the inner edge of the bark of the cut section (Figure 4). Clearly visible rings were traced along each line and counted from the outside (current date) of the section. The position of every 5th ring along a line was marked around the entire section (Figure 4) to identify false and partial rings. Ring-width in mm (growth over one year) was determined by placing a ruler along each of the three radii (1, 2 and 3) with the zero point on the ruler at the inner edge of the bark (Figure 4). The average width of each ring was calculated from the ring widths from the three radii.

Data analysis
STATISTICA statistical package version 7.0 (StaSoft inc, 1984 - 2006) was used for data analysis. The Simple Regression Model was used to test for relationships between number of rings and age, and stem diameter and number of rings, for the key species. The Bonferroni test in STATISTICA was used to determine the relationship that exists in mean radial growth within species under different disturbances, and between different species under similar land use disturbances.
Correlation between number of growth rings and stand age since disturbance cessation
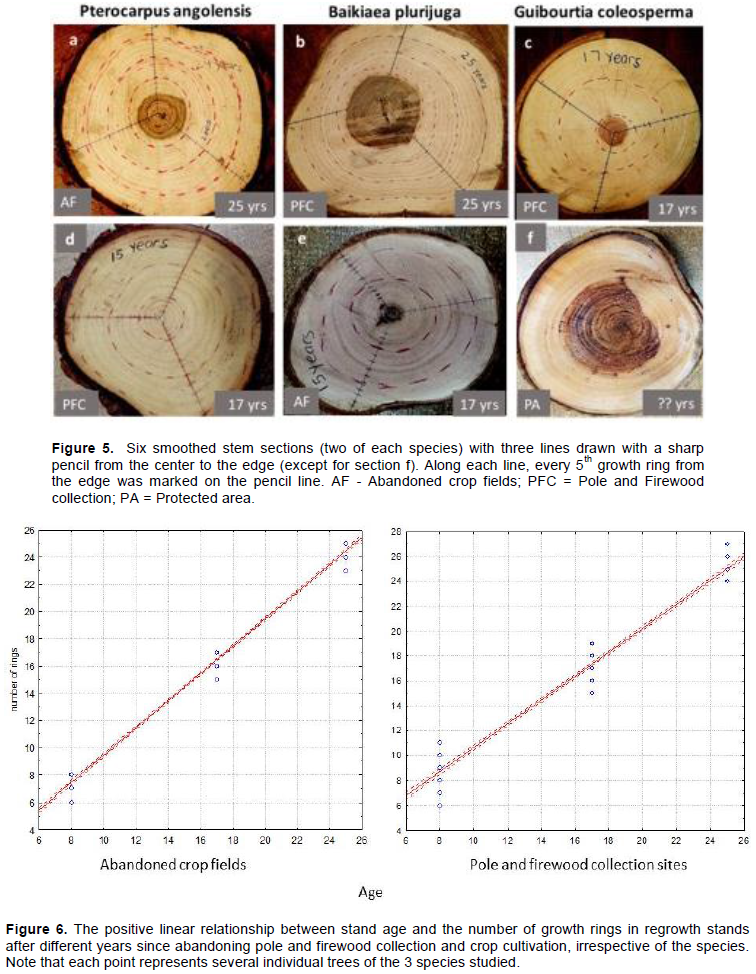
The growth ring boundaries were reasonably distinct for P. angolensis, as shown here for a 25-year old abandoned crop field (AF) site and a 17-year old pole and firewood collection (PFC) site (Figure 5a and d), and for B. plurijuga, as shown here for a 25-year old PFC site and a 17-year old AF site (Figure 5b and e). G. coleosperma did not show very distinct growth rings as shown here for a 17-year old PFC site and a protected area (PA) site (Figure 5c and f). Less clear rings were typical of the discs from mature woodlands (Figure 5f).
The number of growth rings showed a strong positive linear relationship with stand age in both regrowth stands of pole and firewood sites and abandoned crop fields (r² = 0.976; p<0.01; slope of curve = 0.97; n = 180) and (r² = 0.98; p<0.01; slope of curve = 0.99; n = 180 respectively) (Figure 6). Some discs from pole and firewood collection sites had 1 or 2 extra rings whilst those from abandoned crop fields had 1 or 2 fewer rings. However, the discs from mature woodland of the same diameter as those from the regrowth stands did not show any distinct growth rings (Figure 5f).
Correlation between the number of growth rings and DBH
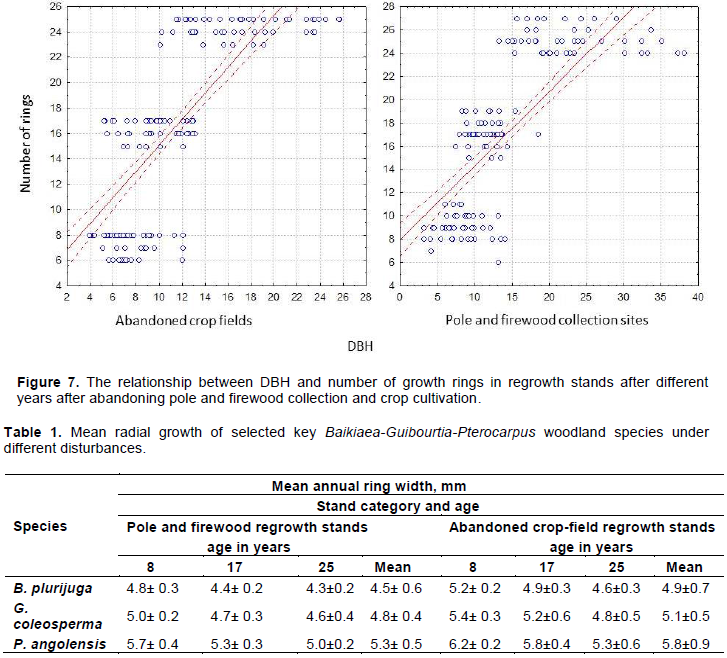
All the species studied showed weak correlation between the number of growth rings and the DBH of a tree, in regrowth stands of both abandoned crop fields (r² = 0.51; p< 0.01; slope of curve = 1.02; n = 180) and pole and firewood collection sites (r² = 0.46; p<0.01; slope of curve = 0.63; n = 180) (Figure 7).
Mean radial growth in regrowth stands from abandoned crop fields and pole and firewood collection sites
Mean annual ring width was significantly different (p<0.005) between species within regrowth stands of the same disturbance category (pole and firewood collection, and abandoned crop-field) and stand age (Table 1). Mean radial growth was not assessed in the undisturbed sites because the stem discs did not show distinct rings making it difficult to measure ring width. P. angolensis exhibited the highest mean radial growth amongst the key species, with the mean ring width of 5.3 mm (10.6 mm radial or diameter growth per year) in pole and firewood regrowth stands and 5.8 mm (11.6 mm radial growth per year) in abandoned crop-field regrowth stands. Generally, the ring width is high in the youngest stands in all three species (Table 1). Thereafter, ring width tends to decrease as the stands get older. However, there is no significant difference in mean ring width within the same species under different disturbance factors.
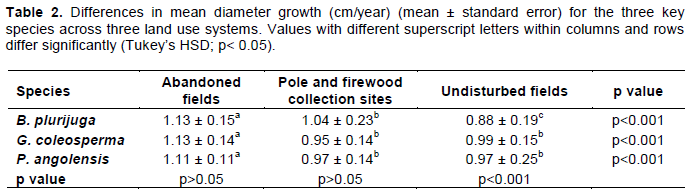
Diameter growth was also assessed for the three species across the different disturbance factors. Results showed a range in diameter growth from 0.88-1.13 cm/year for B. plurijuga; 0.95-1.13 cm/year for G. coleosperma and 0.97-1.11 cm/year for P. angolensis (Table 2). Analysis of Variance results showed significant differences in diameter growth amongst the disturbance factors (F (2,539) = 49.2617; p<0.0001) with abandoned fields having the highest diameter growth. All species showed no significant difference in diameter growth (p>0.05) between the abandoned crop fields and pole and firewood collection sites whilst the opposite is true for undisturbed sites (p<0.001). The Post-hoc Tukey test results showed that the three species had significant differences in diameter growth across the three disturbance factors (B plurijuga (F (2,177) = 26.45, p <0.0001); G. coleosperma (F (2,177) = 25.85, p <0.0001); P. angolensis (F (2,177) = 11.47, p <0.0001). The species had high diameter growth in the abandoned crop fields. B. plurijuga recorded the least diameter growth in the undisturbed sites (Table 2). G. coleosperma and P. angolensis recorded the least diameter growth in pole and firewood collection sites and undisturbed sites.
Stand age-rings and DBH-rings relationships for the key species
Understanding stand dynamics of indigenous woodlands is essential for determining the sustainability of a polycyclic selective harvesting system. Limited related studies have addressed the influence of different disturbance factors on growth of residual trees over time. The few investigations of growth rate, using growth rings, have been based on coring (Stahle et al., 1999) or whole discs (Gourlay, 1995; Ngoma et al., 2017) or a combination of the two. Grundy (1995) based her study on damaging the cambium and cutting the cross section of the stems to allow for counting of growth rings. Gourlay (1995) and Grundy (1995) based their studies on observations at 1.3 to 1.4 m height from the ground. The method has a disadvantage in that some growth rings may be missed because a seedling may take time to reach the 1.3 or 1.4 m height. Coring species with dense wood may be difficult and at times the operator may miss or fail to reach the pith.
Very few studies have been investigated the influence of different disturbance factors on mean radial growth of key species in Baikiaea-Guibourtia-Pterocarpus woodlands of Zimbabwe. Dendrochronology of key species such as P. angolensis in Sikumi and Mzola gazetted forests in Matabeleland (Stahle et al., 1999) and B. plurijuga in Zambia (Miller 1952; Ngoma et al., 2017) have been studied. Age determination using growth rings of G. coleosperma, a key species in the Baikiaea-Guibourtia-Pterocarpus woodlands of north western Zimbabwe (Chigwerewe, 1996), has not yet been considered.
Results from this study showed that B. plurijuga and P. angolensis showed distinct growth rings which can be used as a good estimate for stand age in both pole and firewood and abandoned crop-field regrowth stands. However, G. coleosperma discs did not show distinct rings hence they cannot be used as a good estimate of stand age in both pole and firewood and abandoned crop-field regrowth stands.
Results suggested that there are 1 to 2 missing growth rings in abandoned crop fields and an additional ring in pole and firewood collection sites. This shows that a tree in a 25-year old abandoned crop field will have 24 or 23 rings and a tree in pole and firewood regrowth will have 26 or 27 rings. An additional ring in pole and firewood regrowth stands may be attributed to the fact that young plants are left behind during harvesting for poles and firewood. Missing rings in abandoned crop fields may be attributed to the constant removal of seedlings or sprouts during cultivation. Delayed germination of plants, when the fields are abandoned, could explain why some plants had fewer rings. Delayed stem development may also be due to shoot die back because of frequent fires in woodlands.
The occurrence of distinct growth rings may be attributed to the strong seasonality in both temperature and precipitation experienced in the north-western parts of Zimbabwe (JAFTA and Forestry Commission 2001). The woodlands are deciduous, with trees losing leaves during the dry season and become on leaf before the onset of the rainy season. The seasonality in flowering, leaf flush and leaf fall suggest that radial growth is restricted to the summer wet season (Borchert, 1991) with distinct annual ring formation (Stahle et al. 1999). Strong and consistent diameter growth of free growing stems in regrowth stands contribute to the clear and wider rings (Geldenhuys 2005) because the trees are growing in the absence from intense competition (Rozas, 2001). This could explain why stems from abandoned crop-field regrowth stands showed wider rings, and that rings of stems from undisturbed sites were less clear rings because of the shading effect from larger stems of the canopy. Stems from pole and firewood collection sites also showed wider rings compared to undisturbed sites because more space and resources are released for residual plants after selective harvesting occurs (Montoro et al., 2017). P. angolensis and B. plurijuga had clear rings, thus their cambial growth develops distinctively every year. However, growth rings for G. coleosperma are not very distinct, possibly resulting from poor cambial growth; the species only loses leaves for a very short period of time, or remain green throughout the year (Palgrave, 2002).
The study has revealed a weak significant relationship between DBH and the number of growth rings, in all three selected sites. This implies that the bigger and smaller stems showed the same number of rings. The smaller stems are the suppressed trees and the bigger stems are the fast-growing and vigorous stems in these even-aged stands of light-demanding species. This implies that DBH cannot be used as a reliable estimate of stand age. This contradicts the conclusions of Syampungani et al. (2010) that DBH can be used as a reliable predictor of stand age probably because they selected only bigger stems for the counting of rings; smaller stems were not considered.
Mean radial growth
The three species showed a significant difference in mean radial growth across the land use disturbances, with abandoned crop fields recording the highest ring width and diameter growth, as compared to pole and firewood collection sites. This suggests that there is more growth in the cleared areas, and that trees in open areas grow much faster than in mature stands because the younger plants grow in the absence of intense competition (Rozas, 2001). Several studies have reported high growth rates in disturbed areas than in undisturbed sites (Chapman and Chapman, 1997; Syampungani, 2008; Hawthorne et al., 2012; Gourlet et al., 2013; Khai et al., 2020). Total exposure to light and reduced competition for moisture and nutrients also contribute to increased mean radial growth (Rozas, 2001; Syampungani 2008; Khai et al., 2020). In the undisturbed stands, there is competition for space, light, moisture and nutrients between the young plants and the older trees hence mean radial growth is suppressed (Rozas, 2001). This was also observed in Mozambican Miombo woodlands (Geldenhuys, 2005). Montoro et al. (2017) recorded highest radial growth in younger stands after partial cutting while Khai et al. (2020) recorded highest diameter growth during the early post-harvesting period (5year old stands after selective harvesting) owing to the increases in open space and sunlight availability to residual trees. In their study growth rates decreased to a level of an unlogged stand at 11years post-logging in a Brazilian tropical forest and 16-years post-logging in a Ghanaian high forest. This shows that relative growth rates tend to decline with age of trees, as also mentioned by Johnson and Abrams (2009), Montoro et al. (2017) and Khai et al. (2020), since older stands are mainly composed of old and mature trees.This suggests that selective thinning of suppressed stems, for use as poles and firewood, could maintain good growth of the remaining stems.
Calvert (1986) reported a mean annual DBH increment of 1.78 mm for B. plurijuga, Mushove and Makoni (1993) reported 1.5 mm, and SAREC (1993) reported 1.25 to 2.04 mm for B. plurijuga, 1.02 to 2.37 mm for G. coleosperma and 1.30 to 2.72 mm for P. angolensis. In these studies, mean annual diameter increment was studied on permanent sample plots over several years. FAO pilot studies on forest data gathering and analysis, reported 1.75 mm DBH increment for B. plurijuga, 2.11 mm DBH increment for G. coleosperma and 2.00 mm DBH increment for P. angolensis. Grundy (2006) reported a mean growth of 2.7 cm.year-1 for all trees in an area protected from fire and human disturbance. Syampungani et al. (2010) reported a mean ring width ranging between 4.4 to 5.6 mm in charcoal regrowth stands and slash and burn regrowth stands. Results from this study are almost similar to findings by Syampungani et al. (2010), maybe because of similarity in climatic conditions between the two countries. The intensity of disturbances from the different land use systems in the two studies might also be similar.
The identification of annual growth rings in B. plurijuga, G. coleosperma and P. angolensis for both pole and firewood collection and abandoned crop-field regrowth stands has important implications for forest ecology and management of Baikiaea-Guibourtia-Pterocarpus woodlands. The study concludes several important points. Individual growth rings of P. angolensis and B. plurijuga, and not of G. coleosperma, can be reliably differentiated on cut stems and hence can be used in determining the age of young to intermediate aged Baikiaea-Guibourtia-Pterocarpus woodlands. The number of growth rings is the same in larger and smaller stems in a stand of known age. Trees growing in suppressed stands did not show clear rings compared to free-growing trees. Annual growth rings, and not diameter at breast height, can be used as a good estimate of stand age in both pole and firewood and abandoned crop-field regrowth stands. The mean radial and diameter growth data has shown that trees that grow in open areas (outside undisturbed woodland) grow much faster than trees growing under the canopy of the undisturbed woodlands. Such growth-ring width information would be particularly useful when used together with the climatic data for all the calendar years involved. It would provide an understanding of the disturbance and other biological factors that the land was exposed to in order to relate the growth rings and growth parameters with drought frequencies, disturbance factors and stand development (stand density suppression). However, it can be noted from this study that many stems were cut to determine stand age. This could be avoided if forest managers would monitor closely forest activities and growth of trees in stands of known age since disturbance cessation; hence tree age can be estimated without having to cut them. However, if stand age is not known, then smaller, suppressed two to three trees without any damage to the lower 1 m of the stem could be cut to determine stand age.
Mean radial and diameter growth information generated from the study will help in size predictions of the key Baikiaea-Guibourtia-Pterocarpus species in that the study has provided a means of collecting mean radial and diameter growth data in both pole and firewood sites and abandoned crop fields. The current data may be supplemented with other growth rate data for trees of known age. We therefore conclude that forest managers need to adopt silvicultural practices of selective thinning and limited clear-felling (as disturbance regimes) that would stimulate optimal growth of at least key species in the woodlands.
The authors have not declared any conflict of interests.
The authors thank the Forestry Commission for allowing them carry out the study in their forest areas. Many thanks go to Nicole Chiwota and Mr Sibanda who helped with data collection. Special thanks go to Dr HGT Ndagurwa who assisted immensely with data analysis. This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
REFERENCES
|
Boaler SB (1966). The ecology of Pterocarpus angolensis D.C in Tanzania. Her Majesty, Stationery Office, London.
|
|
|
|
Borchert R (1991). Growth periodicity and dormancy: In Raghavendra AS (ed.), Physiology of trees. John Wileyn. New York, USA pp. 221-245.
|
|
|
|
|
Caetano AVL, Flores BM, Levis C, Clement CR, Roberts P, Schöngart J (2019). Growth rings of Brazil nut trees (Bertholletia excelsa) as a living record of historical human disturbance in Central Amazonia. PLoS ONE 14(4):e0214128.
Crossref
|
|
|
|
|
Calvert GM (1986). The ecology and management of the Kalahari sand forest vegetation of South-western Zimbabwe. In: Pierce GD (ed). The Zambezi teak Forests. Proceedings of the first national conference on the teak forests of Southern Africa. Livingstone, Zambia.
|
|
|
|
|
Chapman CA, Chapman LJ (1997). Forest Regeneration in Logged and Unlogged Forests of Kibale National Park, Uganda. Biotropica 29: 396-412.
Crossref
|
|
|
|
|
Chichinye A, Geldenhuys CJ, Chirwa PW (2019). Land-use impacts on the composition and diversity of the Baikiaea- Guibourtia-Pterocarpus woodlands of north-western Zimbabwe. Southern Forests: a Journal of Forest Science 81(2):151-165.
Crossref
|
|
|
|
|
Chigwerewe SK (1996). The ecology and management potentials of naturally regenerated Teak forests of the Kalahari-Namib sandstone formations in Zimbabwe. In: Mushove PT, Shumba EM, Matose F (eds.), Sustainable Management of Indigenous Forest in the Dry Tropics. Proceedings of an International Conference, ZFC & SAREC-SIDA, Kadoma, Zimbabwe pp. 148-154.
|
|
|
|
|
Fahn A, Burley J, Longman KA, Tomlinson PB (1981). Possible contributions of woody anatomy to the determination of the age of tropical trees. In: Age and growth rate of tropical trees: new direction for research. Yale University School of Forestry Bulletin 94:31-42.
|
|
|
|
|
Fanshawe DB (1956). Regeneration from clear felling. Forestry News Northern Rhodesia P 11.
|
|
|
|
|
Gambiza J (2001). The regeneration of Zambezi Teak after logging, influence of fire and herbivory. Unpublished PhD Thesis, University of Zimbabwe, Harare. Zimbabwe.
|
|
|
|
|
Geldenhuys CJ (2005). Basic Guidelines for Silviculture and Forest Management Practices in Mozambique. Report Number FW-04/05. Pretoria: Forestwood cc.
|
|
|
|
|
Gourlay ID, Barnes R (1994). Seasonal growth zones in the wood of Acacia karoo Hayne: Their definitions and implications. Commonwealth Forest Review 73:121-127.
|
|
|
|
|
Gourlet-Fleury S, Mortier F, Fayolle A, Baya F, Ouedraogo D, Benedet F, Picard N (2013). Tropical forest recovery from logging: a 24 year silvicultural experiment from Central Africa. Philosophical Transactions of the Royal Society B: Biological Sciences 368(1625):20120302.
Crossref
|
|
|
|
|
Grundy IM (1995). Regeneration and management of Brachystegia spiciformis and Julbernadia globiflora in Miombo woodland, Zimbabwe. Unpublished DPhil thesis, University of Oxford.
|
|
|
|
|
Gourlay ID (1995). Growth ring characteristics of some African Acacia species'. Journal of Tropical Ecology 11:121-140.
Crossref
|
|
|
|
|
Grundy IM (2006). Age determination of Miombo species Brachystegia spiciformis (Leguminosae- Caesalpinoideae) in Zimbabwe using growth rings. Southern African Forestry Journal 206(1):5-12.
Crossref
|
|
|
|
|
Hawthorne WD, Sheil D, Agyeman VK, Abu Juam M, Marshall CAM (2012). Logging scars in Ghanaian highforest: Towards improved models for sustainable production. Forest Ecology and Management 271:27-36.
Crossref
|
|
|
|
|
Heita HTN, Ham H, Mensah S (2019). Effects of seed viability and pre-treatments on seed germination of two indigenous species-Strychnos cocculoides Baker and Guibourtia coleosperma (Benth.) Leonard in Namibia. Agroforestry Systems 93(6):2173-2180.
Crossref
|
|
|
|
|
JAFTA and Forestry Commission (2001). Final Report, March 2001. Japan Forest Technical Association (JAFTA) Kogyo Co Ltd, Forestry Commission, Harare, Zimbabwe.
|
|
|
|
|
Johnson SE, Abrams MD (2009). Age class, longevity and growth rate relationships: proctracted growth increases in old trees in the eastern United States. Tree Physiology 29:1317-1328.
Crossref
|
|
|
|
|
Khai TC, Mizoue N, Ota T (2020). Post-Harvest Stand Dynamics over Five Years in Selectively Logged Production Forests in Bago, Myanmar. Forests 11(2):1-13.
Crossref
|
|
|
|
|
Lemmens RHMJ, Louppe D, Oteng-Amoako AA (2008). Plant resources of tropical Africa. Wagenngen: PROTA Foundation, Backhuys CTA.
|
|
|
|
|
Matose F (2002). Local people and reserved forests in Zimbabwe: What prospects for co-management. PhD Thesis, University of Sussex. United Kingdom.
|
|
|
|
|
Mushove PT, Makoni JT (1993). Coppicing ability of Colophospermum mopane. In: Pierce GD, Gumbo DJ (eds.). Proceedings of the International Symposium on the Ecology and Management of Indigenous Forests in Southern Africa, Victoria Falls, Zimbabwe. Zimbabwe Forestry Commission and SAREC, Harare, Zimbabwe. pp. 226-230.
|
|
|
|
|
Montoro GM, Rossi S, Lussier JM, Walsh D, Morin H (2017) Understanding tree growth responses after partial cuttings: A new approach. PLoS ONE 12(2):e0172653.
Crossref
|
|
|
|
|
Moses M (2013). Assessment of trade-offs between timber and carbon values of Pterocarpus angolensis (Kiaat) in the Kavango Region of Namibia - a comparison of current and potential values. MSc thesis, Stellenbosch University, South Africa.
|
|
|
|
|
Mutsiwegota CR, Mudekwe J (1998). Towards shared forest management in Zimbabwe. Paper presented during the international conference on Decentralisation and Devolution of forest management in Asia and the Pacific. 28 November - 4 December 1998. Davao, Philippines.
|
|
|
|
|
Miller RG (1952). A girth increment study of Baikiaea plurijuga in northern Rhodesia. Empire Forestry Review JSTOR 31(1):45-52.
|
|
|
|
|
Ngoma J, Speer JH, Vinya R, Kruijt B, Moors E, Leemans R (2017). The dendrochrological potential of Baikiaea plurijuga in Zambia. Dendrochronologia 41:65-77.
Crossref
|
|
|
|
|
Nemarundwe N, Mbedzi D (1999). Social Development Studies for the Gwayi- Mbembesi settlement option study. Forestry Commission, Harare, Zimbabwe.
|
|
|
|
|
Nyamapfene K (1991). Soils of Zimbabwe. Nehanda Press, Harare, Zimbabwe P 56.
|
|
|
|
|
Palgrave M (2002). Keith Coates Palgrave trees of Southern Africa Edn 3. Cape Town.
|
|
|
|
|
Piearce GD (1993). Natural regeneration on indigenous trees: The key to their successful management. In: Piearce GD, Gumbo DJ (eds.), The Ecology and Management of Indigenous Forests in Southern Africa, pp 347-365, Proceedings of an international symposium, Victoria Falls, Zimbabwe, 27-29 July 1992, Zimbabwe. Forestry Commission and SAREC, Harare.
|
|
|
|
|
Rozas V (2001). Detecting the impact of climate and disturbances on tree-rings of Fagus sylvatica L. and Quercus robur L. in a lowland forest in Cantabria, Northern Spain. Annals of Forest Science,Springer Verlag/EDP Sciences 58(3):237-251.
Crossref
|
|
|
|
|
SAREC (1993). The Ecology and management of indigenous forests in southern Africa: Proceedings of an international symposium, Victorial Falls, Zimbabwe, 27-29 July 1992 Paperback - January 1, 1993.
|
|
|
|
|
Stahle DW, Mushove PT, Cleaveland MK, Roig F, Haynes GA (1999). Management implications of annual growth rings in Pterocarpus angolensis from Zimbabwe. Forest Ecology and Management 124:217-229.
Crossref
|
|
|
|
|
Syampungani S (2008). Vegetation change analysis and ecological recovery of the Copperbelt Miombo woodland of Zambia, PhD thesis. University of Stellenbosch, Stellenbosch, South Africa.
|
|
|
|
|
Syampungani S, Geldenhuys CJ, Chirwa PW (2010). Age and growth rate determination using growth rings of selected miombo woodland species in charcoal and, slash and burn regrowth stands in Zambia. Journal of Ecology and the Natural Environment 2(8):167-174.
|
|
|
|
|
Van Daalen JC, Vogel JC, Malan FS, Fuls A (1991). Dating of Pterocarpus angolensis Trees. South African Forestry Journal 162:1-7.
Crossref
|
|