ABSTRACT
A study on ecological preference of breeding sites of Anopheles gambiae compared to other mosquito species were carried out in small natural microcosms. A total of sixteen small ponds, four each from North, South, East and West of Maiduguri Metropolitan were surveyed for the breeding and non-breeding sites for mosquitoes. In each pond mosquito larvae samples were collected by dip method after identifying species by their angular positions and later confirmed by Polymerase Chain Reaction (PCR). Water samples from which collection of A. gambiae and non-Anopheles gambiae larvae were also collected and analyzed for physico-chemical properties following the standard laboratory procedures. The results revealed that, preference of breeding site of A. gambiae is dependent on the physico-chemical nature of water habitat. Of a total of 30 parameters analyzed, pairwise comparison using SPSS software revealed that 11of the physico-chemical parameters significantly varied (P<0.05) between A. gambiae and non-A. gambiae breeding sites. These parameters include Turbidity, pH, Conductivity, Temperature, Suspended solid, Sodium, Chloride, Sulphate, Calcium, Magnesium, Total hardness and Magnesium hardness. The proportion of A. gambiae in larval habitats were significantly (P<0.05) and positively correlated to these parameters. However, of the eleven parameters, PH (r = 0.429, P = 0.001), temperature (r = 0.269, P = 0.050), calcium(r = 0.256, P = 0.048) and turbidity (r = 0.515, P = 0.000), were found to be the most influential parameters that determine habitat selection for breeding.
Key words: Arid zone, ecological preference, physico-chemical, Anopheles gambiae, distribution.
Previously it was thought that the control of malaria would be easy, based on the assumption that the relationship between the parasites, the vector and the human host was clearly understood, that the effective therapeutic and chemotherapeutic agents were available and that insecticides held a great promise for vector control (Ranson et al., 2009; Ranson and Lissenden, 2016). However, despite the tremendous progress made in the acquisition of the knowledge of biology of the malarial parasites, the human host and the development of anti- malarial drugs, the disease has proven far harder to control (WHO, 2017). Malaria still remains an insidious and ever present scourge that constitutes obstacle to human development (Dobson, 1999; WHO, 2017). At every turn when it was believed that the disease could be eradicated through vector control, the Anopheles mosquito vectors eased their way out of extinction. A prominent factor responsible for this inability to control this disease has been attributed to lack of adequate knowledge of the vector ecology and species composition (CDC, 2010). The role of Anophele gambiae in malaria transmission have been reported to depends largely on the presence of favorable environment for egg laying and larval development (Manuela et al., 2014) that translate to adult populations in a geographically defined area (Rejmánková et al., 2005, Gimnig et al., 2005, Mwangangi et al., 2007). Traditionally, malaria control has been directed to the adult stages and studies of larval ecology have been neglected (Gimnig et al. 2005). This has been the main reason for the dearth of information on factors affecting larval productivity and distribution. A. gambiae complex laid eggs in a wide variety of selected aquatic environments (Himeidan et al., 2013) such as small pools that are partially or completely exposed to the sun, while others prefer to breed in shaded stagnant pools, or even in slow moving water (Lanzaro et al., 1998, Budiansky, 2002). However, the underlying ecological determinant of the distribution is not yet quite understood. For instance under laboratory conditions, A. gambiae carries out normal development when the pH varies as much as from 4.0 to 7.8, as long as there is sufficient phytoplankton and zooplankton for it to consume and maximum larval habitat temperature does not exceed 37°C (Mereta et al., 2013). However, these conditions rarely occur in nature, especially in the intense heat of equatorial Africa, hence the need to understand the ecological characteristics of their natural breeding sites (Paaijmans et al., 2009).
The finding that A. gambiae larvae in their natural habitats tend to aggregate in a particular aquatic microcosm suggests that there could be some variation between physico-chemical properties of the breeding and the non-breeding habitats (Mereta et al., 2013). Understanding this phenomenon would no doubt contribute tremendously towards effective control of malaria transmission from source preferably targeting different parts of the mosquito life cycle (Lardeux et al., 2008; Killeen, 2014). Source reduction (larval control) has been reported to be one of the best methods of choice for mosquitoes control (Killeen et al., 2002; Tusting et al., 2013; Bhatto et al., 2015), especially when mosquitoes species targeted are concentrated in small number of discrete habitats (Kitron and Spielman, 1989). Therefore for effective larval control to be achieved, larval habitat physical and chemical properties need to be considered as one of the priorities in the mosquitos’ abatement program hence the focus of this study. The aim of this study was to determine whether there is variation between ecological factors of habitats between A. gambiae larvae and non- A. gambiae larvae at breeding sites in arid zone of north eastern Nigeria by determining which of the ecological factors that influence the distribution and abundance of A. gambiae larvae at breeding sites.
Study area
The study was carried out in Maiduguri the capital of Borno State, North-Eastern Nigeria. Maiduguri is located within latitude 13°N and 14°N, longitude 12°N and 13°N. The area has a tropical climate with mean temperature of 34.8°C. The hottest part of the year are the months of March and April with the temperature ranging between 30-40°C. During the period between November to February, the weather is usually very cold and dry with a temperature as low as 18°C, being the coldest months. During the hot season, the ambient temperature is usually high between 40-45°C. The relative humidity is around 45% in August and October and gradually lowers to about 5% in April and May. The mean annual rainfall in Borno is about 650 mm/annum for 3-4 month which starts in June and ends in September, followed by a long period of dry season of about 8-9 months.
Identification of breeding sites and collection of water samples for physicochemical analysis
Survey was carried out to locate mosquito breeding and non-breeding sites forth nightly during rainy season from July to October (Gadzama et al., 2011). Water samples taken for physico-chemical analysis taken forth nightly for six months from where mosquito larvae were found and where they were not found. The water samples collected in each site was fixed immediately using alkaline iodide-azide to maintain it biochemical oxygen demand.
Physico-chemical analysis of water samples
The physical factors were measured right on the field using their respective measuring instruments. The chemical analyses of water were done following standard laboratory procedures as described by Fishman and Friedman (1989) and Sunil and Shyam (2017). The parameters measured include: Temperature, pH, conductivity, turbidity, suspended solid, total dissolved solids, sodium concentration, potassium, sulphate (SO4-), phosphates (PO4), calcium, bicarbonate, carbonate, calcium hardness, total hardness, total alkalinity, magnesium, Chloride, fluoride, silica (SiO2), total iron concentration, magnesium hardness, ammonium, dissolved oxygen, biochemical oxygen demand, chemical oxygen demand, nitrite and nitrate.
Data analysis
SPSS software (version 20) was used to carry out the statistical analysis. Pair wise comparison was used to compare mean value between physic-chemical factors of water samples where A. gambiae were found breeding as evidenced by the presence of A. gambiae larvae and where they were not found breeding as evidenced by absence of larvae of A. gambiae but presence of other mosquito species at the time of study. To select in hierarchical order of discriminatory ecological factors influencing habitat selection for breeding by A. gambiae, a stepwise discriminate function analysis was performed on dataset set that were significantly different for the model to select in order of strength the factors that influenced habitat selection for breeding. Multiple regression analysis was adopted to determine dependent factors within the factors.
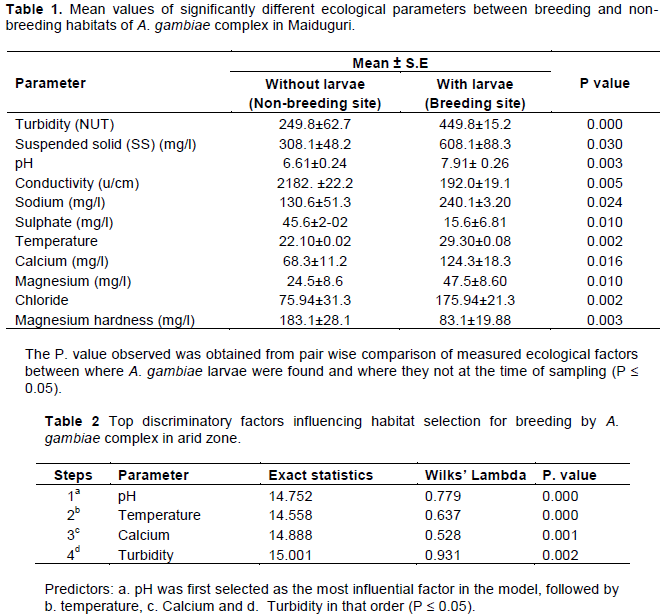
Mean comparison of values of physico-chemical parameters between habitats with Anopheles gambiae and those without
The comparison of the mean values of physic-chemical factors between A. gambiae breeding site and non-breeding sites revealed that 11 out of 30 analyzed parameters were significantly different (P ≤ 0.05). The values of the significantly different factors are presented in Table 1 while Table 2 shows the topmost influential factors from the eleven parameters that differed significantly. To determine which factors were dependent factors among the eleven significantly measured parameters, the results revealed that pH, temperature, calcium and turbidity (Table 2) were found to the most influential factors that discriminates habitat selection for breeding by A. gambiae.
Determination of dependent factors using all the measure parameters
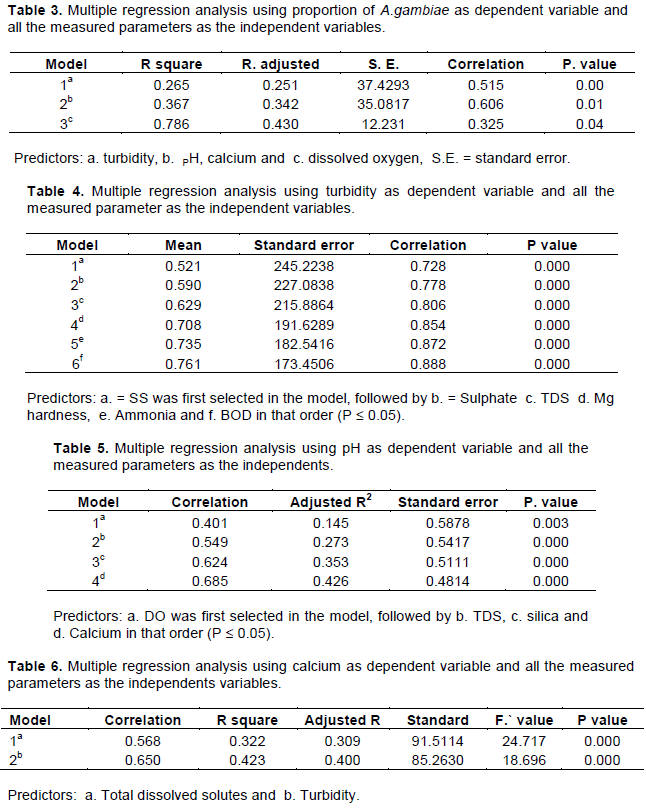
When all the measured parameters were analyzed to select discriminatory factors that influence the presence or absence of A. gambiae in the studied microcosms, the model revealed that turbidity, pH, calcium and dissolved oxygen (Table 3) are the significant factors that determine whether or not A. gambiae larvae would be found at a particular time.
Interrelationship between physico-chemical factors
To determine the association between the selected influential factors and all the measured parameters, each of the influential factors were analyzed as dependent factor on the other factors and the results are presented in Tables 4 to 6.
Water is an important component of ecosystem and its quality in the breeding sites of A. gambiae is an important determinant of whether or not the female Anopheles mosquito will lay their eggs and the resulting immature stages will successfully complete their development to adult stage (Piyaratne et al., 2005; Manuela et al., 2014). In this study the habitat characteristics of A. gambiae were found to be remarkably different from those of other mosquito species. Of the total 30 water parameters analyzed, the breeding habitat for A. gambiae were significantly different for 11 parameters (Table 1) compared to other species (P<0.05). However, of the 11 parameters temperature, pH, turbidity and calcium were found to be the best discriminatory parameters predicting the suitability of breeding habitat for A. gambiae (Table 2) in the model. pH was first selected followed by temperature, calcium and turbidity in that order. These four ecological factors together accounted for 62% of the total variation observed among the predictors (Table 3). All the three models that were applied selected temperature, pH calcium and turbidity in that order. However, when multiple regression analysis was performed using all the 30 analyzed parameters, only temperature, turbidity and dissolved oxygen were selected as best predictors associated with proportion of A. gambiae (Table 4). The two parameters however, accounted for 36.7% of the total variation.
This finding seem to some extent disagree with the report of Mwangangi et al. (2007) who reported that only temperature was found to be significantly associated with A. gambiae s.l. habitat along Kenyan Coast, East Africa. The observed disagreement could be as a result of variation in the ecology of the study regions. Kenya is located in the coastal region with its peculiar characteristics of deep valleys and high elevation, while Maiduguri is located in the arid Sahelian region of West Africa. Water temperature was not expected to vary significantly because all habitats were exposed to direct sunshine and the topographic relief of Maiduguri is flat and all areas will have similar temperature.
Although might not be as obvious as nutrition and larval density, ambient temperature can dramatically affect mosquito production. In the laboratory, most mosquito larvae are reared around 22-27°C which is within the similar range to the mean environmental temperature of 22.10°C found in this study from which the larvae were isolated. Too high or low temperatures can be lethal to almost all shallow aquatic organisms more especially mosquito larvae. For instance, in Anopheles albitarsis and Anopheles aquasalis, colder temperatures have been found to cause delayed embryo eclosion (Ayetkin et al., 2009). Additionally, there was also observed reduction in the hatch rate of A. albitarsis when reared at below 21°C. Similarly, it has also been reported that Anopheles albopictus larvae reared at 26°C pupate faster than those reared at below 22°C; however, fewer larvae completed ecdysis at the higher temperature (Alto and Juliano, 2001). This trend has also been reported in Culex tarsalis (Reisen et al., 1984) and Anopheles sergentii (Beier et al., 1987). In addition to reduced ecdysis rates, adults from temperature stressed environments have been found to have reduced longevity (Beier et al., 1987). Similarly, it was also reported that adults derived from heat stressed larval regimens were found to have reduced life spans by several days as was found in A. gambiae (Afrane et al., 2006), Anopheles superpictus (Ayetkin et al., 2009) and Culex tarsalis (Reisen et al., 1984). In Aedes dorsalis, high temperature regimen had a reduced number of ovarian follicles (Parker, 1982) while A. gambiae derived from the same environment had increased fecundity when compared to adults from a lower temperature region (Afrane et al., 2006). Finally, it was reported that as temperatures increased above the optimum rearing temperature in Anopheles merus, lead to reduction the larval head and adult wing size (Lesueur and Sharp, 1991; Ayetkin et al., 2009). The finding of PH as one of the influential factor for breeding in this study was not a surprise because many studies have shown that the effect of pH is of unquestionable importance and that its significance lies in the fact that, under natural conditions, it dictates the fevourability of association between chemical and biological factors in breeding places upon which the successful of unsuccessful development of larvae depend (MacGregor, 2009). The larvae of certain species tend to actually restrict themselves to water exhibiting a PH index within a definite short range, and that the PH indexes is consequently often reliable as to whether the chemical and biological group associations will fevour or preclude the successful development of such larvae (Malcom, 2009). The finding of Calcium as one of the important factor is not unusual because as reported by White, (1985), Calcium-rich water fevours the growth of macrophytic algae, Chara, whose presence positively correlated with abundance of A. gambiae larvae.
The selection of these parameters could be attributed to the correlation to other factor measured which also directly or indirectly influence habitat selection. For example, The bivariate correlation procedure computed using Pearson’s correlation coefficient revealed that the proportion of An. gambiae in larval habitats were significantly and positively correlated to Turbidity (r = 0.515, P = 0.000), pH (r = 0.429, P = 0.001), Conductivity (r = 0.380, P = 0.005), Suspended solids (r = 0.384, P = 0.004), Dissolved solid (r = 0.443, P = 0.001), Sodium (r = 0.310, P = 0.023), Calcium (r = 0.256, P = 0.048), Magnesium (r = 0.272, P = 0.046), Chloride (r = 0.363, P = 0.007), Sulphate (r = 0.299, P = 0.028) and Temperature (r = 0.269, P = 0.050).
To determine which of the non-selected parameters were either counting to or were associated with turbidity, pH and calcium, multiple regression analysis were performed using these as dependent variables. The results obtained are shown in Table 4 to 6. The results revealed that suspended solid, sulphate, total dissolved solid, magnesium hardness, ammonium and biological oxygen demand were the best variables, together accounting for approximately 79% of the variation existing in turbidity. Furthermore, all were significantly correlated to turbidity (p < 0.05 in all case). However, dissolved oxygen, total dissolved solids, silica and calcium were the best variables associated with pH, together accounting for 47% of the variation existing in PH whereas total hardness and suspended solids were the only variables associated with calcium and accounting for 42% of the observed variation. The pH of water is dependent on the concentration of anions and cations derived from salts, fertilizer compound and other synthetic compounds all together accounted for the pH (Bos, 1991). Therefore, it may directly or indirectly detect the life of any aquatic organisms. The anions and cations may indirectly affect mosquito breeding by fevouring certain aquatic organisms or vegetation on which mosquito larvae feed or affect potential biological control agents of A. gambiae.
Finding from this study suggest that A. gambiae breeding sites were strongly under the influence of ecological factors of the breeding sites at a particular time. Though eleven parameters were found to be significant, however, the top discriminatory factors influencing the distribution and abundance of A. gambiae larvae in small microcosms were found to be pH, temperature, calcium, turbidity and dissolved oxygen.
The authors have not declared any conflict of interests.
REFERENCES
|
Afrane YA, Zhou G, Lawson BW, Githeko AK, Yan G (2006). Effects of microclimatic changes caused by deforestation on the survivorship and reproductive fitness of Anopheles gambiae in western Kenya highlands. American Journal of Tropical Medicine and Hygiene 74:772-778.
Crossref
|
|
|
|
Alto BW, Juliano SA (2001). Temperature effects on the dynamics of Aedes albopictus (Diptera: Culicidae) populations in the laboratory. Journal of Medical Entomology 38:548-556.
Crossref
|
|
|
|
|
Ayetkin S, Ayetkin AM, Alten B (2009). Effect of different larval rearing temperatures on the productivity (Ro) and morphology of the malaria vector Anopheles superpictus Grassi (Diptera: Culicidae) using geometric morphometrics. Journal of Vector Ecology 34:32-42.
Crossref
|
|
|
|
|
Beier MS, Beier JC, Merdan AA, Sawaf BM, Kadder MA (1987). Laboratory rearing techniques and adult life table parameters for Anopheles sergentii from Egypt. Journal of American Mosquito Control Association 3:266-270.
|
|
|
|
|
Bhatt S, Weiss DJ, Cameron E. (2015). The effect of malaria control on Plasmodium falciparum in Africa between 2000 and 2015. Nature 526(7572):207-211.
Crossref
|
|
|
|
|
Bos R (1991). Water Quality, Disease and Human Health. Paper presented at the Water Quality Conference, Bratislava.
|
|
|
|
|
Budiansky S (2002). The mosquito genome: Anopheles gambiae. Creatures of our own making. Science 298:80-86.
Crossref
|
|
|
|
|
CDC (2010). Anopheles mosquitoes. Malaria (04 September 2014).
|
|
|
|
|
Dobson MJ (1999). The Malariology Centenary. Parasitologia 41:21-32.
|
|
|
|
|
Fishman MJ, Friedman LC (1989). Methods for determination of inorganic substances in water and fluvial sediments. U.S. Geological survey techniques of Water-Resources Investigations, book 5, chap A1, 545.
|
|
|
|
|
Gadzama UN, Toyosi RI, Madakan SP, Ali AF (2011). Effect of livestock, human hosts and aquatic habitat distribution on the abundance of Anopheles gambiae complex in Borno State, Arid Zone of Nigeria. International Journal of World Rural Observations 3(4):71-77.
|
|
|
|
|
Gimnig JE, Oombe M, Kamau L, Hawley WA (2005). Characteristics of larval Anopheline (Diptera: Culicidae) habitat in Western Kenya. Journal of Medical Entomology 38:82-88.
|
|
|
|
|
Himeidan YE, Temu EA, El Rayah EA, Munga S, Kweka EJ (2013). Chemical cues for malaria vectors oviposition site selection: challenges and opportunities. Journal of Insects 685-82.
Crossref
|
|
|
|
|
Killeen GF (2014). Characterizing, controlling and eliminating residual malaria transmission. Malaria Journal 13(1):330.
Crossref
|
|
|
|
|
Killeen GF, Fillinger U, Kiche I (2002). Eradication of Anopheles gambiae from Brazil: lessons for malaria control in Africa? Lancet Infectious Diseases 2(10):618-62711.
Crossref
|
|
|
|
|
Kitron U, Spielman A (1989). Suppression of transmission of malaria through source reduction: Anti-anopheline measures applied in Israel, the United States, and Italy. Reviews of Infectious Disease 11:391-406.
Crossref
|
|
|
|
|
Lanzaro GC, Toure YT, Carnahans J, Zheng L, Dolo G, Troare S, Petrarca V, Vernick KD, Taylor C E(1998). Complexity in the genetic structure of Anopheles gambiae population in West Africa as revealed by microsatellite DNA analysis. Proceeding of the National Academy of Sciences USA 95:14260-14265.
Crossref
|
|
|
|
|
Lardeux FJ, Tejerina RH, Quispe V, Chavez TK (2008). A physiological time analysis of the duration of the gonotrophic cycle of Anopheles pseudopunctipennis and its implications for malaria transmission in Bolivia. Malaria Journal 7:141.
Crossref
|
|
|
|
|
Lesueur D, Sharp BL (1991). Temperature-dependent variation in Anopheles merus larval head capsule width and adult wing length: implications for Anopheline taxonomy. Medical and Veterinary Entomology 5:55-62.
Crossref
|
|
|
|
|
Manuela H-V, Jenny L, Steven WL, Ulrike F (2014). Habitat discrimination by gravid Anopheles gambiae sensu lato – a push-pull system, Malaria Journal 13:133.
Crossref
|
|
|
|
|
Mwangangi JM, Mbogo CM, Muturi EJ, Nzovu JG, Kabiru EW, Githure JI, Novak RJ, Beier JC (2007). Influence of biological and physicochemical characteristics of larval habitats on the body size of Anopheles gambiae mosquitoes (Diptera: Culicidae) along the Kenyan coast. Journal of Vector Borne Diseases 44:122-127.
|
|
|
|
|
Mereta ST, Yewhalaw D, Boets P, Ahmed A, Duchateau L, Speybroeck N (2013). Physico- chemical and biological characterization of anopheline mosquito larval habitats (Diptera: Culicidae): implications for malaria control. Parasite and Vectors 6:320.
Crossref
|
|
|
|
|
Paaijmans KP, Read AF, Thomas MB (2009). Understanding the link between malaria risk and climate. Publication of National Academy of Science, USA 106:13844-13849.
Crossref
|
|
|
|
|
Parker BM (1982). Temperature and salinity as factors influencing the size and reproductive potentials of Aedes dorsalis (Diptera: Culicidae). Annual Entomological Society America 75:99-102.
Crossref
|
|
|
|
|
Mattah ADP, Godfred F, Leonard KA, Memuna MM, Dziedzorm KS, Worlasi DK, Langbong B, Michael DW ((2017). Diversity in breeding sites and distribution of Anopheles mosquitoes in selected urban areas of southern Ghana. Parasites and Vector 10:25.
Crossref
|
|
|
|
|
Piyaratne MK, Amerasinghe MP, Amerasinghe PH, Konradsen F (2005). Physico chemical characteristics of Anopheles culifacies and Anopheles varuna breeding water in a dry zone stream in Sri Lanka. Journal of Vector Borne Diseases 42:61- 67.
|
|
|
|
|
Ranson H, Abdallah H, Badolo A, Guelbeogo WM, Kerah-Hinzoumbe C, Yangalbe-Kalnone E, Sagnon N, Simard F, Coetzee M (2009). Insecticide resistance in Anopheles gambiae: data from the first year of a multi-country study highlight the extent of the problem. Malaria Journal 8(1):299.
Crossref
|
|
|
|
|
Ranson H, Lissenden N (2016). Insecticide Resistance in African Anopheles Mosquitoes: A Worsening Situation that Needs Urgent Action to Maintain Malaria Control. Trends Parasitology, Elsevier Ltd.
Crossref
|
|
|
|
|
Rejmánková E, Higashi R, Grieco J, Achee N, Roberts D (2005). Volatile substances from larval habitats mediate species-specific oviposition in Anopheles mosquitoes. Journal of Medical Entomology 42:95-103.
Crossref
|
|
|
|
|
Reisen WK, Milby MM, Bock ME (1984). The effects of immature stress on selected events in the life history of Culex tarsalis. Mosquito News 44:385-395.
|
|
|
|
|
MacGregor ME (2009). The significance of PH in development of mosquito larvae. Parasitology 2(1-2):132-157.
Crossref
|
|
|
|
|
Sunil RV, Shyam NL (2017). Determination of Physico-Chemical Parameters and Water Quality Index (WQI) for drinking water available in Kathmandu Valley, Nepal: A review International Journal of Fisheries and Aquatic Studies 5(4):188-190.
|
|
|
|
|
Tusting LS, Thwing J, Sinclair D (2013). Mosquito larval source management for controlling malaria. Cochrane Database Systematic Reviews (8):CD008923.
Crossref
|
|
|
|
|
White GB (1985). Anopheles bwambae species n., a malaria vector in the Semliki valley, Uganda, and its relationship with other sibling species of the Anopheles gambae complex (Diptera: Culicidae). Systematic Entomology 10:501-522.
Crossref
|
|
|
|
|
World Health Organization (WHO) (2017). World Malaria Report, 2017, Geneva.
|
|