ABSTRACT
Maize lethal necrosis (MLN) disease is a major constraint on maize production in Eastern Africa since its first report in 2012 in Kenya. The main causative agent is maize chlorotic mottle virus (MCMV) which co-infects maize with other viruses from the family Potyviridae. In Africa, Sugarcane mosaic virus (SCMV) is the most common potyvirus that co-infects maize synergistically with MCMV. MCMV can be transmitted by insect vectors, mechanically and via contaminated seeds. Monitoring of MCMV is important in farmers’ fields, seed fields, seed-lots and in grain. Robust diagnostic tools are essential in epidemiological studies, germplasm screening and exchange of disease-free materials across the regions and globally. Therefore, a sensitive, reliable and affordable diagnostic tool for MCMV is necessary in the laboratory and also in the field. A reverse transcription loop-mediated isothermal amplification (RT-LAMP) assay with two-end point analysis was developed to detect MCMV in active vegetative stages and in seed. Six sets of specific primers were designed and evaluated. Amplification was detected in 60 min using the SYBR green colour change and in 10 to 20 min for real-time amplification in the Genie II platform. The assay discriminated the common viruses infecting maize in Eastern Africa. The assay showed excellent specificity to MCMV. The simplicity, rapidity, and inexpensiveness of this technique make it a suitable choice for large-scale sample processing, especially by laboratories with limited resources and for field analysis performed by regulatory agencies in the region.
Key word: Maize lethal necrosis, maize chlorotic mottle virus, diagnostics, loop mediated isothermal amplification.
Abbreviation:
CIMMYT, International Maize and Wheat Improvement Center; DRC, Democratic Republic of Congo; EIAR, Ethiopian Institute of Agricultural Research; ELISA, Enzyme Linked Immunosorbent Assay; KEPHIS, Kenya Plant Health Inspectorate Service; NaCCRI, National Cereal Crops Research Institute, Uganda; NCBI, National Center for Biotechnology Information; NEB, New England Biolabs; NPPOs, National Plant Protection Organizations; OARDC, Ohio Agricultural Research and Development Center; SNNPR, Southern Nations, Nationalities, and People's Region.
Maize chlorotic mottle virus (MCMV) belongs to the family Tombusviridae and is the only species in the genus Machlomovirus (Nutter et al., 1989). The virions of this single-stranded RNA virus are isometric, and the single-component particles have a smooth spherical or hexagonal shape (Scheets, 2000). Previously, two genetically and geographically distinct strains of MCMV were reported, MCMV-P (Peru) and MCMV-K (Kansas) (Nyvall, 1999). However, recent studies on global phylogeny of MCMV reveal distinct MCMV strains for East Africa, North America, China and South America (Braidwood et al., 2018). The virus was first reported to infect Zea mays in Peru (Castillo and Hebert, 1974). MCMV is not widespread in the United States, having been reported only in Nebraska, Kansas (Uyemoto et al., 1980), and Hawaii (Niblett and Claflin, 1978). Globally, the virus occurs in Argentina, Mexico, and Peru and lately Eastern Africa, China, Spain and Ecuador (Quito-Avila et al., 2016). Maize Lethal Necrosis has spread fast in the eastern and central Africa regions after it was first reported in Kenya (Wangai et al., 2012). For instance, in Tanzania, the disease was first reported in 2012 in regions around Lake Victoria and Arusha (CIMMYT Periodic Newsletter, December 2012). The government of Tanzania and The Maize and Wheat Improvement Center (CIMMYT) carried out surveys and tests which revealed the presence of MCMV and SCMV in maize leaf samples (CIMMYT Periodic Newsletter, December 2012). MLN was also reported in Uganda in 2012 in the Kenyan border districts of Busia and Tororo (CIMMYT Periodic Newsletters, 2012) and has been detected in Eastern Uganda districts of Iganga and Mbale (Kagoda et al., 2016). Currently, the disease is present in the western, south western and recently in the northern parts of Uganda (MLN Surveillance reports, NACRRI, Uganda, 2017, 2019). In Rwanda, it was first reported in 2013 and was found endemic in all maize growing districts (Adams et al., 2014). The disease was officially reported in the Democratic Republic of Congo (DRC) predominantly in western provinces of North and South Kivu in 2014 (Lukanda et al., 2014). In Ethiopia, maize plants with MLN symptoms were first observed in 2014 prompting surveillance which led to the first report in 2014 (Mahuku et al., 2015a). Nowadays, the disease is endemic in Gambela, Oromia, Amhara, SNNPR and Gumuz administrative regions of Ethiopia (EIAR MLN surveillance report, 2019). There are reports of MLN in Southern Sudan (Mahuku et al., 2015b), unpublished results) and Burundi (Burundi Institute of Agronomic Sciences (ISABU) - MLN surveillance reports, 2017). MLN has had a serious impact on maize production and grain yield in Eastern Africa. In Kenya, maize yield losses of 23 to 100% were estimated in maize growing counties in 2012-2013. For instance, 26,000 ha of maize were affected by MLN in Kenya in 2012 estimated losses of up to US$ 52 million (Ministry of Agriculture, Kenya, 2012), and by 2013 (De Groote et al., 2016).
Continuous maize production in the field greatly increases the incidence of MCMV, because the virus can be spread by insect vectors, mechanically, and by seed at very low rates of 0.003% (Jensen et al., 1991).
Additionally, MCMV is possibly transmitted through infested soil, as the virus can survive in maize plant residues (Nyvall, 1999). Therefore, it is essential to test for the presence of this virus in seed lots and in the field aiming its management and to limit its spread.
Disease diagnosis and virus detection in maize is important to monitor maize diseases through surveillance programs and to facilitate implementation of appropriate management measures (Riley et al., 2002). MCMV has been detected in leaves, pollen, female and male inflorescences, ear husks, cotyledons, and seeds (pericarps, endosperm, cotyledons, and embryo) (Scheets, 2004). The most reliable methods currently used for detecting MCMV in host tissues include ELISA (enzyme-linked immunosorbent assay), Northern blots, and polymerase chain reaction (PCR). maize chlorotic mottle virus infection may be difficult to diagnose based on symptoms alone, because some of its symptoms (stunting, chlorosis) resemble those caused by nutrient deficiencies, moisture stress or other maize infecting viruses like maize mosaic virus, maize streak virus and maize stripe virus (Marchand et al., 1995).
PCR has been cited for lower levels of sensitivity and specificity compared to quantitative real-time PCR (qPCR) (Mackay et al., 2002). However, qPCR involves acquisition of expensive equipment that are not affordable by most laboratories in developing countries. It is against this background that a simple, versatile and cheap molecular based method of diagnostics for MCMV was developed in this study. This is the Reverse Transcriptase Loop Mediated Isothermal Amplification (RT-LAMP). LAMP is a powerful innovative gene amplification technique used as a simple and rapid diagnostic tool for detection and identification of microbial diseases. This method first described by Notomi et al. (2000) employs a DNA polymerase and a set of four specially designed primers that recognize a total of six distinct sequences on the target DNA. An inner primer containing sequences of the sense and antisense strands of the target DNA initiates LAMP. Amplification can be achieved in 1 h under isothermal conditions with a set of six primers in a single tube. Amplification can be detected by a turbidimeter, colourimetric detection (Wong et al., 2015), agarose gel electrophoresis or by a real time fluorimeter platform. Since its discovery, LAMP has been widely used for diagnosis of various plant and animal disease worldwide (Parida et al., 2004, 2006).
The current documented LAMP assays for detection of MCMV (Chen et al., 2016; Zhanmin et al., 2016) exploit the colour change end point analyses which have limitations of uncertainty in correct colour change contrast observations and the high risk of false positives due to contamination of the testing equipment and environment. The other documented LAMP assay for MCMV detection (Xu et al., 2017) employs the colorimetric and turbidity visual assessment also with limitations as earlier indicated.
MCMV is a highly stable virus (Wang et al., 2015) hence the opening of reaction tubes for incorporating the DNA intercalating dyes leads to massive contamination and false positives (Hsieh et al., 2014) as used in the aforementioned MCMV LAMP assays.
As such, an extra real time end point analysis and a colourimetric end point analysis assay which does not involve handling of dyes and reaction tubes opening is vital. This study aimed at developing a LAMP assay with a two-end point analysis for the detection of MCMV. The two-end point analysis includes the observation of colour change of the SYBR green dye from orange to green without opening the tubes and real-time graphical plots of the amplification based on the detection of fluorescence in the samples using the Genie II platform. The Genie II allows real-time isothermal amplification to be performed on a low power portable platform. The closed tube system used in Genie II avoids any post-amplification handling, eliminating laboratory contamination from the amplified product. The isothermal amplification master mix that allows fluorescence detection of the product on the Genie II platform can also be used on generic qPCR instrumentation.
Ribonucleic acid (RNA) extraction and detection of MCMV by conventional RT-PCR
Total RNA was extracted from fresh leaf samples infected with MCMV sourced from the quarantine screen house of Ohio Agricultural Research and Development Center (OARDC) Wooster, Ohio State University. The ZR RNA MiniPrep™ kit (Zymo Research, USA) was used for RNA extraction from the samples. The quality and concentration of RNA was tested using a NanoDrop ND-1000 Spectrophotometer (Thermo Fischer Scientific, Wilmington USA). An RNA Agarose gel electrophoresis was used for quality check of the extracted RNA. A 2-step RT-PCR was performed with the samples to verify the presence of MCMV using primers (MCMV F 5’ – CCG GTC TAC CCG AGG TAG AAA – 3’ and MCMV R 5’ – TGG CTC GAA TAG CTC TGG ATT T – 3’) - Research unit at OARDC, Ohio State University. The first step involved cDNA synthesis using Maxima First strand cDNA synthesis kit for RT-qPCR, with ds DNase (Thermo Fisher Scientific, USA). The 20 µl reaction mixture contained 4 µl 5X reaction mix, 2 µl Maxima Enzyme mix, 1 µl of the RNA template and 13 µl of nuclease free water. The mixture was incubated at 25°C for 10 min, 50°C for 15 min and the reaction was terminated at 85°C for 5 min. PCR was then performed on the cDNA synthesized with a 50 µl reaction mixture containing 25 µl of Dream Taq, 0.1 µl of both forward and reverse primers, 2 µl of the cDNA and 22.8 µl of nuclease free water. The following cycle parameters were used: denaturing 95°C for 30 s, annealing 49.4°C for 30 s and Extension 72°C for 1 min. Final extension was 72°C for 15 min.
RT-LAMP Primer design
Complete sequences of MCMV genomes were collected from NCBI database (Table 1). The sequences were subjected to multiple sequence alignment using the Clustal Omega software (http://www.ebi.ac.uk/Tools/msa/clustalo/) to obtain the consensus sequences (Goujon et al., 2010). The target primer was designed using the Primer Explore V4 software (Notomi et al., 2000). The LAMP primer designing support software program, Netlaboratory, Japan was accessed at http://venus.netlaboratory.com. Regarding software instructions, four key factors were considered for LAMP primer design: (i) Melting Temperature (Tm); (ii) Stability at the end of each primer; (ii) GC content, and (iv) Secondary structure formation.
Considering these parameters more than 30 possible LAMP primers were generated and six sets were selected for meeting all the requirements as described in the Primer Explorer V4 manual.
These primers were synthesized by Invitrogen Corporation, 1600 Faraday Avenue, P.O. Box 6482, Carlsbad CA, 92008, United States. All the six sets of each candidate primer were HPLC-purified.
RT-PCR using the LAMP inner primers
RT-PCR was performed using the derived inner LAMP primers on MCMV RNA used in the conventional RT-PCR earlier mentioned. The conventional RT-PCR procedure was used as earlier described. Detection of MCMV occurred by a one-step reverse transcriptase PCR (RT-PCR) assay performed in a GeneAmp 9700 PCR System thermocycler (Applied Biosystems, Foster City, CA, USA).
The inner primers used derived from the primer design and synthesis were:
FIB (F):
CACGGTAGGACACGGAGTACGAATTGTGGCTATCCCCAAAGC and
BIP (R):
CACGGTAGGACACGGAGTACGAATTGTGGCTATCCCCAAAGC
LAMP primer sets evaluation through initial reaction. The modified NEB LAMP protocol (Table 2) was used for LAMP primers evaluation.
The 25 µl reaction tubes with LAMP reaction contents were incubated at 65°C for 1 h and the reaction terminated by inserting the tubes in water bath at 80°C. The DNA intercalating dye, SYBR Green (1 µl) was placed in the inner part of the cap before closing the tubes to enable colour change visualization thereby avoiding aerosol contamination by the amplified product.
Analysis of the RT-LAMP products was mainly by observing colour change of the reaction mixture from orange to green for positive samples (Soliman and El-Matbouli, 2006). This is due to intercalation with the amplified RT-LAMP products produced. The LAMP primer efficiency evaluation was replicated 4 times to determine their reproducibility.
One step RT-LAMP in Genie II LAMP machine
The Genie II LAMP Assay instrument (Unit 5 Blatchford Road Horsham West Sussex RH13 5QR UK) was used. It works under an isothermal amplification temperature and can be programmed for different temperatures. For the specific primer set, the optimum temperature was determined as 65°C with detection duration of pproximately 20 min. The subsequent melting process (in Genie II called annealing process) starts with 98°C and ends with a temperature of 80°C with a ramp rate of 0.05°C/s. The reaction mixture was the same as illustrated in Table 1 with 1 μl fluorochrome dye (FD) added but maintaining the 25 μl reaction volume by reducing RNA free water by 1 to 11.69 μl. Before adding Bst 2.0 DNA Polymerase, Transcriptor Reverse Transcriptase and RNA template, mixes were incubated at 95°C for 5 min to melt any primer multimers and cooled immediately on ice for 5 min.
Specificity analysis
The developed LAMP assay was evaluated for its specificity to MCMV amplification by carrying it out with other maize infecting viruses namely Maize dwarf mosaic virus (MDMV), Wheat streak mosaic virus (WSMV), Sugarcane mosaic virus (SCMV) and Panicum mosaic virus (PMV). Samples with a combination of MCMV and SCMV were also included in the specificity evaluation. This test was performed with both colourimetric end point and real-time analysis on the Genie II LAMP machine.
Sensitivity of detection analysis: RT-LAMP, conventional PCR and qRT-PCR assays
The sensitivity of detection among RT-LAMP, conventional RT-PCR and qRT-PCR was compared using 9-fold serial dilutions of the virus-positive total RNA extracts. The LAMP sensitivity test was evaluated in the Genie II instrument while the conventional RT-PCR was carried out in the T100™ Thermal Cycler (Life Science Research, Bio-Rad) and the qRT-PCR in the 7500 Fast Real-time PCR System (Thermo Fisher Scientific, USA), respectively. The reaction volume for qRT-PCR was 25 μl which comprised 12.5 μl of the general master mix, 1 μl of the F and R primers each, 0.5 μl of the probe, 0.05 μl of the reverse transcriptase MMLV and 1 μl of the RNA template. The master mix contained Mg, dNPTs, BA and 8.95 μl of ddH2O. The cycling temperatures for MCMV amplification were 2 min at 50°C, 10 min at 90°C, 15 s at 95°C for 40 Cycles, 1 min at 60°C.
The same was performed for conventional RT-PCR and RT-LAMP with a 9-fold dilution. In all these assays, the original concentration used 10° was 25 ng/µl. These dilutions gave the following concentrations (ng): 25, 2.5, 0.25, 0.025, 0.0025, 0.00025, 0.000025, 0.0000025, and 0.00000025. The detection limits were determined by the lowest RNA concentration that provided a positive result.
Primer efficiency evaluations results
Table 3 shows the six primer sets that were designed for MCMV using the software Primer Explorer V4.
LAMP inner primers testing
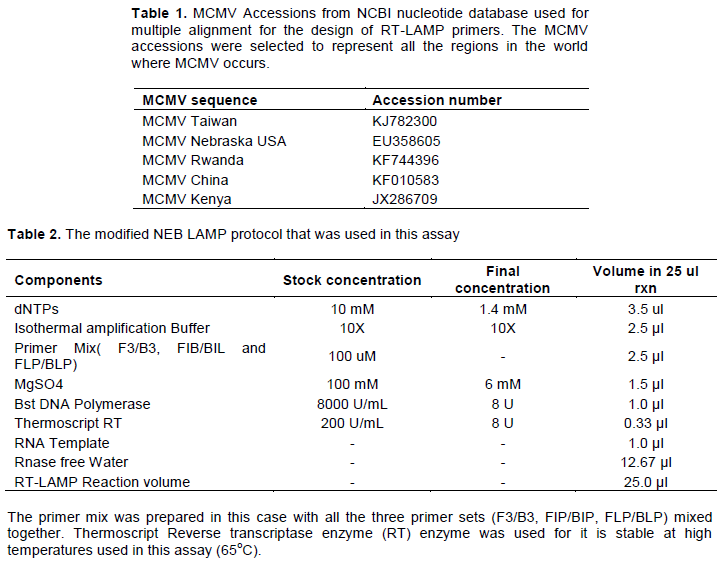
Based on RT-PCR performed with samples from Kenya, the LAMP inner primers FIB(F) and BIP(R) were successful for amplification of all positive MCMV samples as shown in Figure 1. All the RT-PCR evaluation with the outer primers for the LAMP assay amplified MCMV material in the samples tested.
All the 6 sets of inner primers amplified MCMV positive samples through the conventional RT-PCR using the designed inner LAMP primers (FIB and BIP) (Figure 1). This indicates that the primers can amplify the target MCMV genome sequence. However, the inner primers of the set 5 (5FIP and 5BIP) were used for conventional RT-PCR detection of MCMV as described earlier.
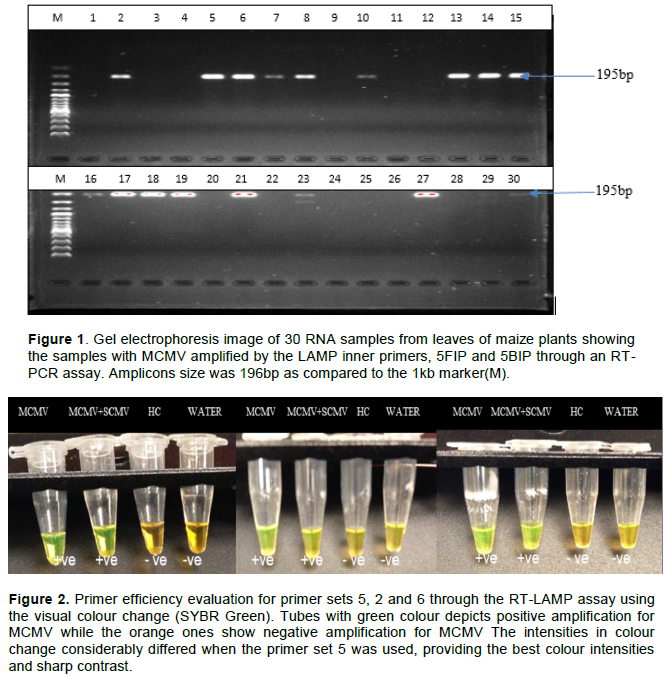
Amplification for MCMV and a combination of MCMV and SCMV by colour change (SYBR Green) assay and in Genie II was performed. Primer efficiency was analyzed by colour change intensity. All primer sets were able to promote amplification of MCMV, but at different efficiency levels. The colour changes were visually analyzed for that samples presenting clear variation of colour (Figure 2). The positive samples had colour changes from orange to green. The negative samples remained orange presenting no amplification of MCMV in those samples for the healthy control (HC) and water.
Specificity analysis
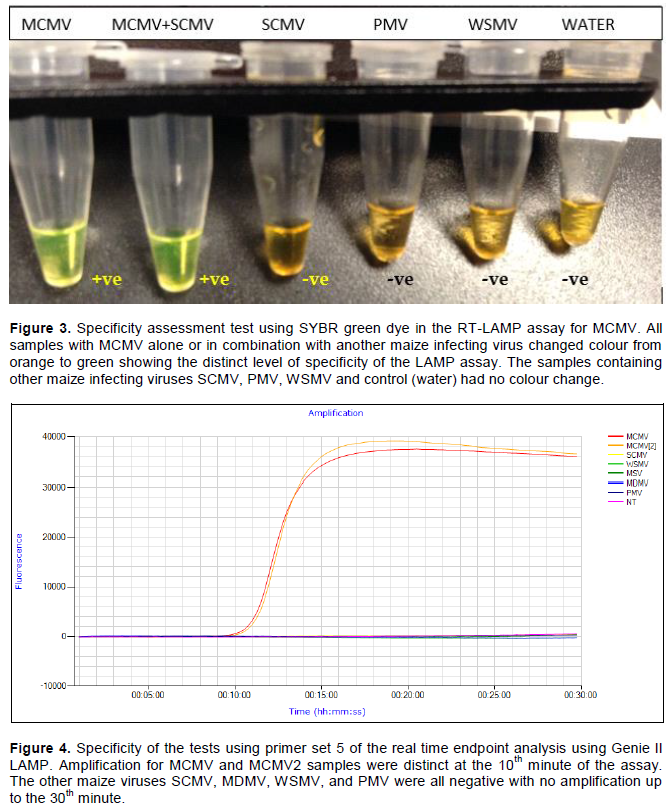
For the determination of specificity of the MCMV primer set, LAMP assay was carried out on samples with other viruses infecting maize namely Maize dwarf mosaic virus (MDMV), Wheat streak mosaic virus (WSMV), Sugarcane mosaic virus (SCMV) and Panicum mosaic virus (PMV). Samples with a combination of MCMV and SCMV were also included in the specificity evaluation. The samples with only MCMV or with a combination with SCMV presented colour shift to green demonstrating the amplification of MCMV (Figure 3). This clearly shows that the primers were specific to MCMV.
Real time end point analysis using Genie II LAMP
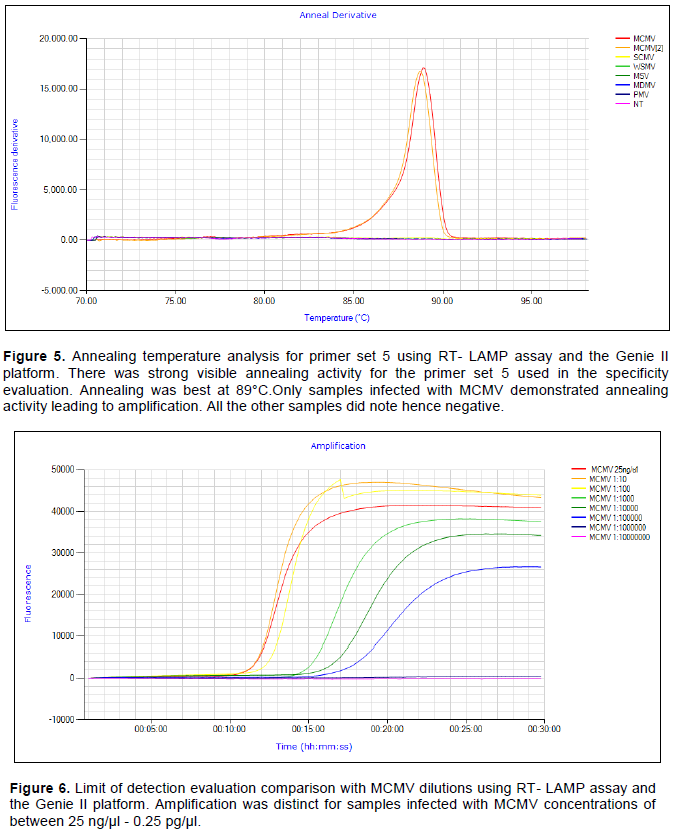
Specificity analysis was repeated using the Genie II LAMP assay with real-time evaluation. The amplification was only evident with MCMV samples. Other common maize infecting viruses namely SCMV, MDMV, MSV, WSMV and PMV were not detected by the assay hence demonstrated the high level of specificity of the assay as illustrated in Figure 4. Only the MCMV and MCMV2 samples were amplified showing the presence of the MCMV virus in these samples.
Only MCMV viruses’ samples showed the annealing activity which further describes the specificity of the LAMP primer sets shown in Figures 4 and 5. There was no primer annealing activity with samples without MCMV further demonstrating the specificity of the LAMP primer sets designed and synthesized.
MCMV RT-LAMP limit of detection (Sensitivity) results
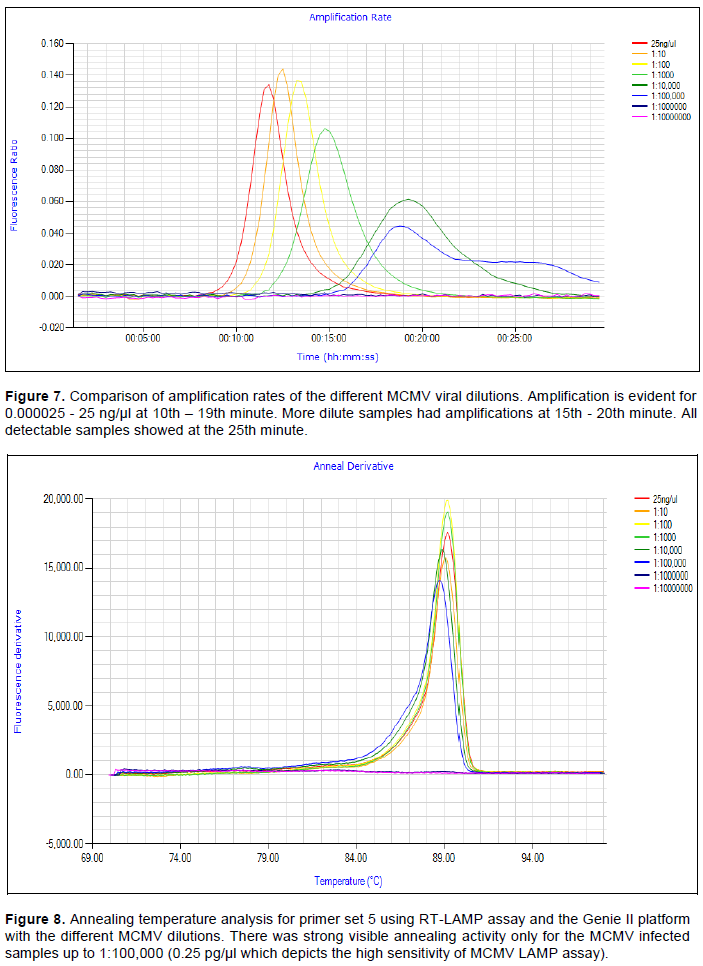
The MCMV LAMP assay hereby developed demonstrated a high level of sensitivity with six-fold diluted samples being detected (Figure 6) and the amplification rates were only evident with the same sample dilutions (Figure 7). This was equivalent to a concentration of 2.5 fg/µl. Samples within the range dilution of 25 ng/µl to 2.5 fg/µl were detected in this assay. The annealing activity occurred only with MCMV samples with concentrations between 25 and 2.5 pg/µl (Figure 8). The same MCMV concentrations evaluated under the MCMV qRT-PCR also detected MCMV up to 6-fold detection as in the LAMP assay (Figure 9). However, the conventional RT-PCR was able to detect MCMV up to 4-fold dilution of 0.0025 ng/ µl. This is shown in Figure 10 with amplification on the gel electrophoresis picture showing bands only in lane 4, 25 ng/µl to lane 8, 0.0025 ng/μl.
Several remedial actions have been taken to reduce the devastating effects of MLN which has threatened food security and people’s livelihoods in the Eastern Africa region (Mahuku et al., 2015b). This includes breeding and deployment of resistance varieties and observing several cultural and agronomic practices. Seed transmission has been attributed to the rapid spread of the disease in the Eastern Africa region (Mahuku et al., 2015b; Boddupalli et al., 2020). Therefore, testing for the disease during active vegetative growth and in seed is vital for both seed certification agencies and seed producers to limit seed contamination and designing feasible management practices (Uyemoto, 1983). There is a thin
line between seed contamination and seed transmission but seed contamination by MCMV leads to seed transmission of this virus (CIMMYT MLN Epidemiology Project Report, 2019).
Currently, seed certification and plant health agencies in eastern Africa use RT-PCR for testing the virus in seed lots as a certification procedure and for confirmation in plant health laboratories. This involves expensive equipment for the tests in the laboratory and long turnaround time for the test results. The RT-PCR test is also expensive with some agencies charging up to USD 35 per sample tested. A detailed two end point analysis RT-LAMP was developed under this study to provide a reliable, effective efficient, sensitive, cost effective and field deployable molecular based assay for the detection of MCMV. The existing MCMV LAMP assay developed by Chen et al. (2016) and Zhanmin et al. (2016) only reports one end point analysis of the use of SYBRgreen dye and hydroxynapthol blue dye, respectively. In these assays, the dye was added in the tubes after incubation which can lead to cross contamination (Hsieh et al., 2014); hence false positives when not handled carefully. LAMP assays result in millions of copies of the target nucleic acid and therefore chances of cross contamination in the lab are quite high. The test cost was also analyzed and found to be USD 7.5 compared to the current USD 35 for RT-PCR.
All the LAMP primer sets developed during this study amplified the target regions of the MCMV genome but with varying efficiencies in MCMV detection. The six sets yielded amplicons of between 165 and 209 bp, hence quite suitable for the maximum activity of the DNA displacement Taq Polymerase (Ignatov et al., 2014). However, primer set 5 and 6 gave the best amplifications with the clearest contrast on colour change from orange to green and sharp curves on the Genie II platform. Primer set five was selected for all subsequent tests in this study and is the one already adopted by KEPHIS and KALRO for their MCMV testing on the LAMP Genie II platform. As indicated earlier, the assay was replicated four times and hence proved to be reproducible. The primer sets also discriminated against the common viruses infecting maize in the region, namely SCMV, MDMV, WSMV, PMV and MSV. The real-time amplification was carried out in the Genie II LAMP instrument as earlier mentioned. This instrument features low power requirements and includes a rechargeable lithium-polymer battery that can keep it running for a full working day. The lithium battery can be charged on the power outlets or using the car charger system hence suitable for field deployment. The Genie II assay analysis also showed the excellent specificity to MCMV (Figures 4 and 5) of this assay with absolutely no chances of cross contamination if all precautions are considered. This is because all the reaction tubes are not opened during the assay and are discarded safely after every reaction procedure. In the Genie II platform, an annealing curve generated in the fluorimeter confirms the presence of the amplification product. This eliminates the need for gel electrophoresis or turbidity detection and allows for a closed-tube system hence eliminating cross contamination that can lead to false positives.
The test also showed the same level of sensitivity with Real time RT-PCR detecting up to 106 dilutions of the initial RNA concentration of 25 ng/ul (Figures 6, 7, 8 and 9). The test was however more sensitive than the conventional RT-PCR which detected up to 4 (104) fold dilutions (Figure 10). In a study comparing RT-LAMP and RT-PCR for MCMV, the detection limit of RT-LAMP assay was 280 fg of MCMV RNA, which was 100 times more sensitive than that of RT-PCR assay (Shan et al., 2017). In the same study, high species-specificity of RT-LAMP method was confirmed by the assay of 5 pathogens such as MCMV, SCMV, MDMV, WSMV and Maize white line mosaic virus (MWLMV). Seed infected with MCMV can be identified through a grow out test using this method with the same sensitivity as qRT-PCR. Further sensitivity optimization may yield a protocol that can be utilized in detecting these viruses directly in seed maize without a grow out test which requires both time and resources to grow the seedlings in germination chambers.
The simplicity, rapidity, and inexpensiveness of this technique make it a suitable choice for large-scale sample processing, especially by laboratories with limited resources. Currently, phytosanitary regulatory institutions are using conventional and real time PCR for routine screening of seed material for MCMV which is quite expensive. However, trial runs for Genie II RT-LAMP for MCMV optimization done in KEPHIS prompted the Kenyan NPPO to adopt the use of this developed assay in their Plant pathology laboratories both in the KEPHIS headquarters and in The Plant Quarantine and Biosecurity station. The assay has also been adopted by the Kenya Agriculture, Livestock Research Organization (KALRO).
Strict regulation coupled with internal controls for self-regulation by seed companies utilizing clean seed maize in conjunction with growing resistant varieties and monitoring new virus strain emergence are necessary measures to prevent the spread of MCMV throughout the maize industry and in the region. MLN free seed Protocols (https://mln.cimmyt.org/) developed for clean seed production by CIMMYT and partners in eastern Africa (Boddupalli et al., 2020) will also go a long way in ensuring low or nil incidences of MLN viruses in both farmers’ fields and commercial seed fields. This will further contribute immensely in the efforts towards limiting the spread of this deadly disease to the southern African region where maize is a vibrant industry.
CIMMYT, International Maize and Wheat Improvement Center; DRC, Democratic Republic of Congo; EIAR, Ethiopian Institute of Agricultural Research; ELISA, Enzyme Linked Immunosorbent Assay; KEPHIS, Kenya Plant Health Inspectorate Service; NaCCRI, National Cereal Crops Research Institute, Uganda; NCBI, National Center for Biotechnology Information; NEB, New England Biolabs; NPPOs, National Plant Protection Organizations; OARDC, Ohio Agricultural Research and Development Center; SNNPR, Southern Nations, Nationalities, and People's Region.
The authors have not declared any conflict of interests.
The author thank the US government through the USDA-FAS office in Nairobi for granting Francis Mwatuni a research fellowship under the Norman Borlaug fellowship to carry out this work in the department of Plant Pathology, OARDC-Wooster, Ohio State University. They also recognize the extremely valuable guidance by Dr. Margaret Peg Redinbaugh and Dr. Sally A. Miller during the entire research study at OARDC. Their uttermost gratitude also go to Christine Willey, Xing Ma, Fulya Baysal for their technical backstopping on the bench while in OARDC, Ohio State. They are indebted to Ivan Obare and Bramwel Wanjala for facilitating optimization of the Genie II LAMP assay in KEPHIS Plant Health Laboratory, Kenya. With the support from BMGF, we were able to optimize this developed RT-LAMP protocol on the Genie II platform for use in the laboratory and in the field.
REFERENCES
|
Adams IP, Harju VA, Hodges T, Hany U, Skelton A, Rai S, Deka MK, Smith J, Fox A, Uzayisenga B, Ngaboyisonga C, Uwumukiza B, Rutikanga A, Rutherford M, Ricthis B, Phiri N, Boonham N (2014). First report of maize lethal necrosis disease in Rwanda. New Diseases Reports 29:22.
Crossref
|
|
|
|
Boddupalli P, Suresh LM, Mwatuni F, Beyene Y, Makumbi D, Gowda M, Olsen M, Hodson D, Worku M, Mezzalama M, Molnar T (2020). Maize lethal necrosis (MLN): Efforts toward containing the spread and impact of a devastating transboundary disease in sub-Saharan Africa. Virus Research 282:197943.
Crossref
|
|
|
|
|
Braidwood L, Quito-Avila DF, Cabanas D, Bressan A, Wangai A, Baulcombe DC (2018). Maize chlorotic mottle virus exhibits low divergence between differentiated regional sub-populations. Scientific Reports 8(1):1-9.
Crossref
|
|
|
|
|
Burundi Institute of Agronomic Sciences (ISABU) (2017). MLN surveillance report 2017. Burundi Institute of Agronomic. Sciences (ISABU).
|
|
|
|
|
Castillo J, Hebert TT (1974). A new virus disease of maize in Peru. Fitopatologia 9(2):79-84.
|
|
|
|
|
Chen L, Zhiyuan J, Dongmei L, Xingliang L, Zihao X, Congliang D, Tao Z, Zaifeng F (2016). One-step reverse transcription loop-mediated isothermal amplification for the detection of Maize chlorotic mottle virus in maize. Journal of Virological Methods 240:49-53.
Crossref
|
|
|
|
|
CIMMYT MLN Epidemiology Report (2019). Maize Lethal Necrosis Diagnostics and Prevention of Seed Transmission. Available at: View
|
|
|
|
|
De Groote H, Oloo F, Tongruksawattana S, Biswanath D (2016). Community-survey based assessment of the geographic distribution and impact of maize lethal necrosis (MLN) disease in Kenya. Crop Protection 82:30-35.
Crossref
|
|
|
|
|
EIAR MLN surveillance Report (2019). Ethiopian Institute of Agricultural research (EIAR). Available at:
View
|
|
|
|
|
Goujon M, McWilliam H, Li W, Valentin F, Squizzato S, Paern J, Lopez R (2010). A new bioinformatics analysis tools framework at EMBL-EBI. Nucleic Acids Research 38(suppl_2):W695-W699.
Crossref
|
|
|
|
|
Hsieh K, Mage PL, Csordas AT, Eisenstein M, Soh HT (2014). Simultaneous elimination of carryover contamination and detection of DNA with uracil-DNA-glycosylase-supplemented loop-mediated isothermal amplification (UDG-LAMP). Chemical Communications 50(28):3747-3749.
Crossref
|
|
|
|
|
Ignatov KB, Ekaterina VB, Arkady FF, Konstantin AB, Tatiana VK, Vladimir MK (2014). A strong strand displacement activity of thermostable DNA polymerase markedly improves the results of DNA amplification. Biological Techniques 57:81-87.
Crossref
|
|
|
|
|
Jensen SG, Wysong DS, Ball EM, Higley PM (1991). Seed transmission of maize chlorotic mottle virus. Plant Diseases 75:497-498.
Crossref
|
|
|
|
|
Kagoda F, Gidoi R, Isabirye BE (2016). Status of maize lethal necrosis in eastern Uganda. African Journal Agricultural Research 11:652-660.
Crossref
|
|
|
|
|
Lukanda M, Owati A, Ogunsanya P, Valimunzigha K, Katsongo K (2014). First report of maize chlorotic mottle virus infecting maize in the Democratic Republic of the Congo. Plant Diseases 98:1448-1449.
Crossref
|
|
|
|
|
Mackay IM, Arden KE, Nitsche A (2002). Real-time PCR in virology. Nucleic Acids Research 30:1292-1305.
Crossref
|
|
|
|
|
Mahuku G, Lockhart BE, Wanjala B, Jones MW, Kimunye JN, Stewart LR, Cassone BJ, Sevgan S, Nyasani JO, Kusia E, Kumar PL (2015b). Maize lethal necrosis (MLN), an emerging threat to maize-based food security in sub-Saharan Africa. Phytopathology 105(7):956-965.
Crossref
|
|
|
|
|
Mahuku G, Wangai A, Sadessa K, Teklewold A, Wegary D, Ayalneh D, Adams I, Smith J, Bottomley E, Bryce S, Braidwood L (2015a). First report of maize chlorotic mottle virus and maize lethal necrosis on maize in Ethiopia. Plant Diseases 99:1870-1874.
Crossref
|
|
|
|
|
Marchand JL, Peterschmitt M, Reynaud B, Dintinger J (1995). Maize streak virus, maize stripe virus and maize mosaic virus in the tropics (Africa and islands in the Indian Ocean). Agriculture and Development Special Issue. pp. 55-69. Available at:
View
|
|
|
|
|
Ministry of Agriculture Kenya (2012). Ministry of Agriculture, Livestock and Development Biannual report.
|
|
|
|
|
National Cereal Crops Research (NaCCRI) (2017). MLN Surveillance reports 2017. National Cereal Crops Research Institute (NaCRRI, Uganda). Available at:
View
|
|
|
|
|
National Cereal Crops Research (NaCCRI) (2019). MLN Surveillance reports 2018. National Cereal Crops Research Institute (NaCRRI, Uganda). Available at:
View
|
|
|
|
|
Niblett CL, Claflin LE (1978). Corn lethal necrosis a new virus disease of corn in Kansas. Plant Disease 62:15-19.
|
|
|
|
|
Notomi T, Okayama H, Masubuchi H, Yonekawa T, Watanabe K, Amino, N, Hase T (2000). Loop-mediated isothermal amplification of DNA. Nucleic Acids Research 28(12):e63-e63.
Crossref
|
|
|
|
|
Nutter RC, Scheets K, Panganiban L, Lommel SA (1989). The complete nucleotide sequence of maize chlorotic mottle virus. Nucleic Acids Research 17:3163-3177.
Crossref
|
|
|
|
|
Nyvall R (1999). Field crop diseases (Third Edition). Iowa State University Press, Ames, Iowa. ISBN-13: 978-0813820798.
|
|
|
|
|
Parida MM, Guillermo P, Inoue S, Hasebe F, Morita K (2004). Real-time reverse transcription-loop-mediated isothermal amplification for rapid
Crossref
|
|
|
|
|
detection of West Nile virus. Journal of Clinical Microbiology 42:257-263.
|
|
|
|
|
Parida MM, Santosh SR, Dash PK, Tripathi NK, Saxena P, Ambuj S, Sahini AK, Laksman PV, Rao MK (2006). Development and evaluation of reverse transcription loop mediated isothermal amplification assay for rapid and real-time detection of Japanese encaphalitis virus. Journal of Clinical Microbiology 44(11):4172-4178.
Crossref
|
|
|
|
|
Quito-Avila DF, Alvarez RA, Mendoza AA (2016). Occurrence of maize lethal necrosis in Ecuador. A disease without boundaries? European Journal Plant Pathology 146:705-710.
Crossref
|
|
|
|
|
Riley M, Williamson MR, Maloy O (2002). Plant disease diagnosis. The Plant Health Instructor.
Crossref
|
|
|
|
|
Scheets K (2000). Maize chlorotic mottle machlomovirus expresses its coat protein from a 1•47-kb subgenomic RNA and makes a 0•34-kb subgenomic RNA. Virology 267:90-101.
Crossref
|
|
|
|
|
Scheets K (2004). Maize chlorotic mottle virus; Viruses and Virus Diseases of Poaceae (Gramineae). H. Lapierre and P. A. Signoret, eds. Institut National de la Rocherche Agronomique, Paris, France. pp. 642-644.
|
|
|
|
|
Shan C, Zhou Y, Li X, Yang S, Shao W (2017). Comparison and application of RT-LAMP and RT-PCR methods for MCMV detection. Guangdong Agricultural Sciences 7:14.
|
|
|
|
|
Soliman H, Ei-Matbouli M (2006). Reverse transcription loop-mediated isothermal amplification (RT-LAMP) for rapid detection of viral hemorrhagic septicaemia virus (VHS). Veterinary Microbiology 114:205-213.
Crossref
|
|
|
|
|
Uyemoto JK (1983). Biology and control of maize chlorotic mottle virus. Plant Disease 67:7-10.
Crossref
|
|
|
|
|
Uyemoto JK, Bockelman DL, Claflin LE (1980). Severe outbreak of corn lethal necrosis disease in Kansas. Plant Disease 64:99-100.
Crossref
|
|
|
|
|
Wang CY, Zhang QF, Gao YZ, Zhou XP, Ji G, Huang XJ (2015). Insight into the three-dimensional structure of maize chlorotic mottle virus revealed by Cryo-EM single particle analysis. Virology 485:171-178.
Crossref
|
|
|
|
|
Wangai AW, Redinbaugh MG, Kinyua ZM, Mahuku G, Scheets K, Jeffers D (2012). First report of Maize chlorotic mottle virus and maize lethal necrosis in Kenya. Plant Disease 96(10):1582-1583.
Crossref
|
|
|
|
|
Wong YP, Othman S, Lau YL, Radu S, Chee HY (2017). Loop-mediated isothermal amplification (LAMP): a versatile technique for detection of micro-organisms Journal of Applied Microbiology 124:626-643.
Crossref
|
|
|
|
|
Xu Y, Liu Y, Qiu Z, Zheng W (2017). Reverse Transcription-loop-mediated Isothermal Amplification for Detection of Maize chlorotic mottle virus. Agricultural Science and Technology 1:123-126.
|
|
|
|
|
Zhanmin L, Xueying X, Cuiyun Y, Junyi H (2015). Colourimetric detection of Maize chlorotic mottle virus by reverse transcription loop-mediated isothermal amplification (RT-LAMP) with hydroxynapthol blue dye. Royal Society of Chemistry Advances 6:73-78.
Crossref
|
|