Limited treated options to Salmonellosis exist due to the emergence of extended spectrum beta lactamases (ESBLs). This study determined the prevalence of common ESBL genes (blaSHV, blaCTX-M, blaOXA) and profiled their antimicrobial resistance drug pattern. A descriptive cross-sectional study that involved 16 Salmonella enterica isolates from diarrhoeic children at the University of Calabar Teaching Hospital and 39 isolates from healthy birds in poultry farms within the Calabar metropolis. Standard disc diffusion method determined antimicrobial susceptibility while Polymerase Chain Reaction (PCR) using specific primers identified the beta-lactamase genes. All 55 isolates phenotypically demonstrated ESBL production. All 55 salmonella isolates were resistant to Cefotaxime and Nalixidic acid. All the clinical isolates were susceptible Ciprofloxacin and Norflaxacin. The poultry isolates however exhibited varied degrees of resistance to Ciprofloxacin and Norflaxacin. All 55 Salmonella isolates carried the blaSHV genes, blaCTX-M gene (94.5%) blaOXA gene (18.2%). Molecular characterization identified the blaSHV gene as the most prominent. This study provides relevant data that would help in the implementation of policies regarding the use of antimicrobial agents in clinical and veterinary settings as well provide antimicrobial treatment guidelines that would strengthen infection control processes.
Salmonella enterica serovars remains one of the major causes of food borne gastroenteritis. Non-typhoid salmonellosis usually causes diarrhoea that may not necessarily require antimicrobial therapy (Andino and Hanning, 2015; Cohen Stuart et al., 2012; Nadimpalli et al., 2019). Nonetheless, in cases where the non-typhoid infection becomes invasive, severe complications may result hence requiring antimicrobial treatment options (Cohen Stuart et al., 2012). The emergence and spread of antibiotics resistance especially with the detection of extended spectrum beta lactamases (ESBLs) in Salmonella enterica serovars has however limited
treatment options because of their acquired resistance to most beta-lactam antibiotics and fluoroquinolones which would have been sufficient first line treatment for both mild and severe salmonellosis (Al Naiemi et al., 2008; Ifeanyi et al., 2015).
Studies have shown ESBL production by
Salmonella enterica to be plasmid mediated and readily transmitted amongst members of the Enterobacteriaceae family as a result (Rotimi et al., 2008). This potential exacerbates the spread of antimicrobial resistance amongst the other members of the Enterobacteriaceae family against beta-lactam antibiotics and other commonly used antibiotics (Oghenevo et al., 2016). Studies have also shown that ESBLs occur because of mutations in the
blaTEM-1,
blaTEM-2 or
blaSHV-1 genes, which are commonly found in the
Enterobacteriaceae family. Others enzymes, notably from the cefotaxime resistance family (CTX-M), represent plasmid acquisition of broad-spectrum beta-lactamases originally determined by
chromosomal genes (Bajpai et al., 2017). These mutations gave rise to the alterations in their amino acid configuration and conferred on these enzymes the ability to hydrolyze a broader spectrum of antibiotics (Bajpai et al., 2017).
More than 350 different naturally occurring ESBL variants have been discovered and classified into nine distinct structural and evolutionary families based on their amino acid sequence comparisons (Sharma et al., 2010). They include, blaTEM, blaSHV, blaCTX-M, blaPER, blaVEB, blaGES, blaBES, blaTLA, and blaOXA (Sharma et al., 2010).
Not much data have been documented regarding the prevalence of the ESBL-producing genes responsible for the antimicrobial drug resistance, especially in Nigeria. This study determined the prevalence of the common ESBL genes (blaSHV, blaCTX-M, blaOXA) in Calabar state, Nigeria and profiled their antimicrobial resistance drug pattern.
Study area
This study was carried out within the Calabar Metropolis in Cross River state, Nigeria. Calabar is the capital city of Cross River State in the South-South geopolitical zone of Nigeria. Cross River State shares boundaries with Benue State to the North, Ebonyi and Abia States to the west and to the east by the Republic of Cameroun. The city is administratively divided into Calabar Municipal and Calabar South Local Government Areas (LGAs). Calabar covers a surface area of about 406 km2 (157 sq mile) and a population of 371,022 at the 2006 census.
Study design
This was a descriptive cross-sectional study designed to investigate the prevalence of common ESBL (blaOXA, blaSHV, blaCTX-M) genes among Salmonella strains obtained from diarrheic patients in the University of Calabar teaching hospital and local poultry farms within the Calabar metropolis. Study period was between November 2017 and February 2018.
Ethical consideration
The Ethical Committees of the selected hospital approved the protocol for this study. Approval was also obtained from the Cross River State Ministry of Health, conveyed via CRS/MH/ CGSE-H/018/Vol/123 and the Health Research Ethical Committee of University of Calabar. Consent was sorted from all study participants and guardians of the poultry farms prior to the collection of samples.
Isolation and identification of Salmonella species
Sixteen (16) Salmonella enterica isolates were randomly recovered from the faeces of diarrhoeic children attending the University of Calabar teaching hospital and 39 Salmonella enterica isolates obtained from healthy birds randomly selected in some local poultry farms within the Calabar Mertopolis. This gave a total of 55 Salmonella isolates. Each of the isolates was sub-cultured on MacConkey agar prior to confirmation. Each typical Salmonella enterica colony was confirmed by means of the 16s rDNA amplification.
The typical Salmonella colonies were further sub-cultured overnight in 5 ml of Luria Bertani (LB) broth and centrifuged at 14000 rpm for 3 min. The cells were re-suspended in 500 µl of physiological saline and heated by means of a heating block at 95°C for 20 min. The heated bacterial suspensions were then brought to cool on ice and re-centrifuged at 14000 rpm for 3 min. The supernatant (containing the DNA) was transferred to a 1.5 ml micro-centrifuge tube and stored at -20°C for further analysis. DNA quantification of the extracted genomic DNA was carried out using the NanoDrop 1000 Spectrophotometer.
The DNA of each typical Salmonella isolate was extracted and used for the amplification of the 16S rRNA region of the isolates’ DNA using a specific primer set (27F: 5'-AGAGTTTGATCMTGGCTCAG-3’ and 1492R: 5'-CGGTTACCTTGTTACGACTT-3'), by means of the ABI 9700 Applied Biosystem thermal cycler at a final volume of 50 microliters for 35 cycles. The Polymerase Chain Reaction (PCR) mix included: X2 Dream Taq Master Mix supplied by Inqaba, South Africa (Taq polymerase, DNTPs, MgCl2), the primer sets at a concentration of 0.4 M and the extracted DNA as template. The PCR conditions were as follows: Initial denaturation at 95ºC for 5 min, denaturation at 95ºC for 30 s, annealing temperature at 52ºC for 30 s, extension at 72ºC for 30 s for 35 cycles and final extension at 72ºC for 5 min. The product was resolved on a 1% agarose gel at 120 V for 20 min and viewed by means of a UV trans-illuminator.
DNA sequence analysis was performed using direct sequencing of both strands by means of the BigDye Terminator kit on a 3510 ABI sequencer (Inqaba Biotechnological, Pretoria - South Africa). The obtained DNA sequences were edited using TraceEdit. Highly similar sequences were downloaded from GenBank in the National Center for Biotechnology Information using BLASTn (http://www.ncbi.nlm.nih.gov/blast). The Salmonella enterica isolates were confirmed by a 100% match with the 16S rDNA of the downloaded sequences of Salmonella enterica.
Antimicrobial susceptibility testing and detection of ESBL production
This study considered the use of thirteen commercially available antibiotic discs types (LiofilChemDiagnostici, LD) which included; Norfloxacin (NOR-10 μg), Nalidixic acid (NA-30 μg), Clotrimoxazole (CLO-50 μg), Amikacin (AK-30 μg), Ciprofloxacin (CIP-5 μg), Imipenem (IMI-10 μg), Chloramphenicol (C-30 μg), Ceftazidime (CAZ-30 μg), Cefotaxime (CTX-30 μg), Cefepime (FEP-30 μg), Aztreonam (ATM-30 μg), ESBL screen disc kit (Cefotaxime, Cefotaxime+Clavilanic acid, Cefotaxime+Clxacillin and Cefotaxime+Clavilanic acid+ Cloxacillin). The susceptibility of the isolates to antimicrobial agents was determined using the Kirby-Bauer disk diffusion method (Cheesbrough, n.d.; Fallah et al., 2013). A bacterial lawn was prepared by transferring 4 to 5 colonies of the same morphological type in a tube containing 2.5 ml sterile normal saline by means of a sterile inoculating loop. The suspension was vortexed and its turbidity compared with barium chloride (0.5 McFarland Turbidity Standard; 1.0 x 108 CFU/µL). The optical density of the standard was regularly monitored with a spectrophotometer at λ=625 nm and 1 cm light path (ODλ=0.08 - 0.1) (Cheesbrough, n.d.). One hundred micro liters of the inoculum was spread on Iso-sensitivity test agar plates. The excess inoculum was siphoned with sterile Pasteur pipettes. Plates were allowed to dry at room temperature in a laminar flow. Disks containing predetermined amounts of antibiotics were then dispensed unto the bacterial lawn using a pair of sterile forceps and gentle pressure applied to ensure complete contact with the agar. The disks were placed 15 mm away from the edge of the plate and 25 mm away from each other. The plates were inverted within 15 minutes after the disks were dispensed, and incubated at 37°C for 16 to 18 h. After incubation, they were examined by reading the diameters of the inhibition zones and interpreted in accordance with the description of the United States Clinical and Laboratory Standard Institute (Cheesbrough, n.d.). The isolates showing resistance towards more than three different groups of antimicrobials such as quinolones, aminoglycosides, and cephalosporins, etc., were considered multidrug resistant strains.
The detection of ESBLs producers using the ESBL screen disc kit was performed on the isolates that showed resistance to the third generation cephalosporin antibiotics by disc diffusion method. Briefly, a disc of amoxicillin-clavulanic acid-30 µg was placed on the center of the Muller-Hinton agar plate inoculated with the resistant strain, followed by placing three different cephalosporin discs (cefotaxime-30 µg, ceftrixone-30 µg and ceftazidime-30 µg) around the amoxicillin-clavulanic acid-30 µg, 20 mm each and incubated for 24 h at 37°C.
Examination/interpretation of plates
Enhancement of the inhibition zone, indicating synergy between clavulanic acid and any one of test antibiotics, will be regarded as presumptive for the production of ESBLs. Zone diameters of Ceftriaxone ≤ 26 mm, Cefotaxime ≤ 24 mm and Ceftazidime ≤ 28 mm will be considered presumptive for ESBLs production by screening.
Quality control
Klebsiella pneumoniae 700603 and Escherichia coli 25922 were used as positive and negative control strains respectively for ESBLs production.
Detection of blaCTX-M, blaSHV and blaOXA genes by PCR
The CTX-M gene was amplified using the primer set CTX-M/F: 5'-CGCTTTGCGATGTGCAG-3' and CTX-M/R: 5'-ACCGCGATATCGTTGGT-3’ on an ABI 9700 Applied Biosystems thermal cycler at a final volume of 25 μl for 35 cycles. The PCR mix included: X2 Dream Taq Master Mix supplied by Inqaba, South Africa (Taq polymerase, DNTPs, MgCl), the primers at a concentration of 0.4 M and the extracted DNA as template. The PCR conditions were as follows: Initial denaturation, 95ºC for 5 min; denaturation, 95ºC for 30 s; annealing, 52ºC for 30 s; extension, 72ºC for 30 seconds for 35 cycles and final extension, 72ºC for 5 min. The PCR product was resolved on 1% agarose gel at 120V for 20 min and visualized on a UV trans-illuminator.
The SHV gene was amplified using the primer set SHV/F: 5'-CGCCTGTGTATTATCTCCCT-3' and SHV: 5'-CGAGTAGTCCACCAGATCCT-3' on an ABI 9700 Applied Biosystem thermal cycler at a final volume of 25 microliters for 35 cycles. The PCR mix included: X2 Dream Taq Master Mix supplied by Inqaba, South Africa (Taq polymerase, DNTPs, MgCl), the primers at a concentration of 0.4 M and the extracted DNA as template. The PCR conditions were as follows: Initial denaturation, 95ºC for 5 min; denaturation, 95ºC for 30 s; annealing, 56ºC for 30 s extension, 72ºC for 30 s for 35 cycles and final extension, 72ºC for 5 min. The product was resolved on 1% agarose gel at 120V for 15 min and visualized on a UV trans-illuminator.
The OXA gene was amplified using the primer pair OXA/F: 5'-AGCCGTTAAAATTAAGCCC-3' and OXA/R: 5'-CTTGATTGAAGGGTTGGGCG-3' on an ABI 9700 Applied Biosystems thermal cycler at a final volume of 25 microliters for 35 cycles. The PCR mix included: X2 Dream Taq Master Mix supplied by Inqaba, South Africa (Taq polymerase, DNTPs, MgCl), the primers at a concentration of 0.4 M and the extracted DNA as template. The PCR conditions were as follows: Initial denaturation, 95ºC for 5 min; denaturation, 95ºC for 30 s; annealing, 55ºC for 30 s; extension, 72ºC for 30 s for 35 cycles and final extension, 72ºC for 5 min. The product was resolved on a 1% agarose gel at 120V for 15 min and visualized on a UV trans-illuminator.
Statistical analysis
The data generated in the cause of this study from the antimicrobial susceptibility tests were analyzed using the Statistical Package for Social Sciences (SPSS Inc., Chicago, IL, USA, version 22.0.). P-values of less than 0.05 (< 0.05), was considered of statistical significance.
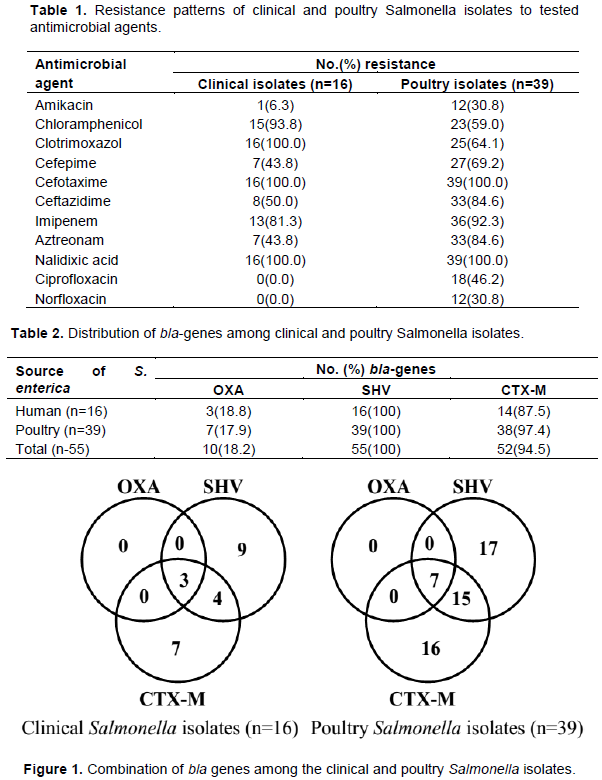
Table 1 revealed the antimicrobial resistance profile of the Salmonella enterica isolates from clinical and poultry sources. All the Salmonella enterica isolates (n=55) from both clinical and poultry sources produced ESBL. All the Salmonella isolates from the clinical source (n=16) were resistant to Clotrimoxazol. All 55 the Salmonella isolates were resistant to Cefotaxime and Nalidixic acid. All the clinical Salmonella enterica isolates showed no resistance to Ciprofloxacin and Norfloxacin.
Table 2 showed the distribution of blaSHV, blaOXA and blaCTX-M genes. All 55 Salmonella enterica isolates carried the blaSHV gene while blaOXA and blaCTX-M were observed in 18.2%(10) and 94.5%(52) of the isolates, respectively. The proportions of occurrence of the bla genes in both the clinical and the poultry Salmonella isolates were not of statistical significance (P>0.05). The blaSHV and blaCTX-M occurred well in predominance over the blaOXA gene in both the clinical and poultry Salmonella enterica isolates.
Figure 1 characterized the combination of bla genes possessed by the Salmonella enterica isolates from clinical and poultry sources. From the clinical sources, 7 and 9 S. enterica isolates carried only blaCTX-M and blaSHV while 4 S. enterica isolates carried both the blaCTX-M and blaSHV genes in combination. Also, all three bla genes under study were carried by 3 clinical Salmonella isolates.
From the poultry isolates, 16 and 17 isolates respectively carried only the blaCTX-M and blaSHV gene while 15 isolates carried both the blaCTX-M and the blaSHV gene combination. Also, 7 isolates carried all three bla genes under study in combination.
It became imperative to profile antibiotic resistance patterns because incorrect identification of antibiotic resistance usually leads to inappropriate antibiotic prescription, which may in turn select and give rise to new resistant genes. This study profiled antibiotic resistance and determined the prevalence of the common ESBL-producing genes (blaSHV, blaCTX-M, blaOXA) in Calabar state, Nigeria.
S. enterica isolates were extracted from 55 isolates of poultry and clinical sources. Based on the result of the ESBL phenotypic screening test kit carried out in this study, all the S. enterica isolates were ESBL producers. All 55 isolates demonstrated 100% resistance to nalixidic acid and 100% susceptible to ciprofloxacin and norflaxacin antibiotics. This agrees with several studies carried out in India where salmonella isolates demonstrated 100% resistance to nalixidic acid with susceptibility to ciprofloxacin (Crump et al., 2003; Kownhar et al., 2007; Ye et al., 2018). These antibiotics belong to the quinolone/fluoroquiniolone drug class. Studies have shown Quninolone resistance to be as a result of mutations in the DNA gyrase (gyrA and gyrB) and topoisomerase IV encoding (parC and parE) genes (Heisig et al., 1995; Ye et al., 2018). Other studies have also reported the presence of plasmid mediated quinolones resistant (PMQR) genes carried by the ESBL-producing plasmid, which facilitates the selection of higher-level resistance to quinolone drugs (Ahamed Riyaaz et al., 2018; Carfora et al., 2018; Jacoby et al., 2014; Kongsoi et al., 2015). A combination of both factors with the possible presence of PMQR genes to a greater extent, we believe could be responsible for the observed resistance to Nalixidic acid and the increased resistance to Ciprofloxacin and Norfloxacin in the poultry isolates.
Other studies have also suggested that the in vitro resistance to nalixidic acid could be used as a pointer to the actual level of in vitro resistance to ciprofloxacin (Campioni et al., 2017; Klemm et al., 2018). According to the Clinical and Laboratory Standard Institute (CLSI) guidelines, the resistance to any antimicrobial agent in the fluoroquinolone drug class has an impact on the resistance of other antimicrobial agents within this drug class (Sahu et al., 2018). This implies that the resistance observed by the Salmonella enterica to nalidixic acid in this study is a pointer to the development of resistance to other members of the fluoroquinolone class of antimicrobial agents such as ciprofloxacin and norfloxacin in humans in the Calabar region. This is important information given that fluoroquinolones are regarded as the antimicrobial class of first choice for the treatment of severe infections caused by S. enteica as well as other pathogenic Enterobacteriaceae in humans.
Results further indicated that fluoroquinolone resistance was higher in poultry isolates in comparison to the clinical isolates. This agrees with several studies carried out in Europe, Asia and North America where resistance to fluoroquinolones by S. enterica was far higher in animals than in humans (Britto et al., 2018; Gouvêa et al., 2015; Nelson et al., 2007). Given that poultry is one of the most preferred foods of animal origin, this study suggests that the resistant S. enterica isolates in humans could have come from animals via the food chain.
This study showed high rates (93.8 and 59% from clinical and poultry sources respectively) of resistance by the Salmonella isolates to chloramphenicol. This finding was in consonance with a similar study carried out in Brazil were the sensitivity of chloramphenicol was found to be as high as 95% among children with salmonellosis (Brown et al., 2018; Ferreira et al., 2011). The high resistance observed suggests the frequent use of chloramphenicol in both the clinical and veterinary setting.
The emergence of ESBL resistance to beta-lactam antibiotics has also introduced a new hurdle in the treatment of salmonellosis as extended-spectrum cephalosporins (beta-lactam antibiotics drug class), which were typically the antibiotics of choice for mild salmonellosis has become insufficient. Results showed an enhanced degree of resistance to the third and fourth generation Cephalosporins (Cefepime, Cefotaxime, Ceftazidime) thus thwarting the reliability of these antibiotics in the treatment of salmonellosis. In judicious use in both humans and poultry, we believe could be the most probable reason for the selective pressure and consequent resistance to these drugs. Therefore, we suggest strict selection and rotation of antimicrobial agents coupled with the continuous monitoring of susceptibility profiles of antimicrobial agents in order to manage the emergence and spread of cephalosporin resistance.
All the salmonella isolates in this study carried the ESBL-producing blaSHV gene in combination with the blaCTX-M and blaOXA genes making blaSHV the most prominent ESBL-producing gene from S. enterica isolates in this study.