Full Length Research Paper

ABSTRACT
Bowdichia virgilioides Kunth (Fabaceae) is a tree species used in folk medicine against inflammatory disorders, and it is still unclear whether consumption of the B. virgilioides stem bark extract has any effect on immune cells. Thus, this study aimed to evaluate the effects of B. virgilioides stem bark extract (AEBv) on the functions of the thymus and B lymphocytes. The AEBv was obtained by decoction and administrated orally once a day in male mice for 7 days. After treatment, thymi were obtained for morphology analysis, relative weight, cellularity and deposition of laminin. Thymocytes obtained from control and treated animals were submitted to adhesion and migration assays followed by immunophenotyping. Also, mesenteric lymph nodes were removed, and the lymphoid cells were obtained to measure B cell proliferation and cytokine production. Mice treated with AEBv and their controls were also immunized with sheep red blood cells (SRBC), and 8 days post immunization the sera were collected to perform antibody titration using hemagglutination assays. AEBv administration decreased thymus weight and cellularity and increased the laminin deposition without any significant effect on the expression of the laminin receptor VLA-6 in the thymus microenvironment. Additionally, thymocytes of AEBv-treated mice showed reduced adhesion and migration on laminin substrates, and the B lymphocytes showed reduced proliferation, enhanced IL-10 secretion, decreased TNF-α production, and attenuated anti-SRBC antibody production. These findings showed that AEBv intake might cause modulatory effects on the functions of thymocytes and B-lymphocytes, contributing to the treatment of immune diseases.
Key words: Thymocytes, cell migration, cell adhesion, immunosuppression, B cell.
INTRODUCTION
It is widely recognized that alternative complementary methods have gained much attention in the treatment of various illnesses. Brazilian biodiversity, especially plant diversity, comprises a wide range of bioactive compounds with potential for the development of new drugs. Among them is Bowdichia virgilioides Kunth (Fabaceae), a tree found in the Northeast of Brazil and commonly known as “sucupira-preta”. In folk medicine, decoctions of the B. virgilioides stem bark are used to treat rheumatism, arthritis, osteoarthritis, skin diseases, diabetes, pain, and wound healing (Almeida et al., 1998; Alburquerque et al., 2007; Macedo et al., 2018). Experimental data have corroborated with the popular use of B. virgilioides. For instance, its anti-nociceptive, -inflammatory, -allergic, and wound healing properties have previously been reported (Silva et al., 2010, 2016; Thomazzi et al., 2010; Agra et al., 2013). Additionally, other effects have also been reported, including antimalarial (Deharo et al., 2001) and anxiolytic effects (Vieira et al., 2013). However, its other biological activities are yet to be characterized, especially on immune cells.
Chemical studies have shown that the stem bark of B. virgilioides has high concentrations of natural steroids and flavonoids (Melo et al., 2001) whose immunosuppressive activities have been described in murine cells (Chen et al., 2015; Le et al., 2017). The search for medicinal plants with immunomodulatory properties has intensified in recent years (Asadi-Samani et al., 2017; Ma et al., 2017; Azad et al., 2018; Bui et al., 2019; Chen et al., 2019). However, specific effects of the consumption of plant extracts on T cell differentiation in the thymus and B-lymphocyte functions are poorly known.
It is widely known that failures in the development and functions of lymphoid cells lead to impaired immunological activity, causing several diseases, including infections, autoimmunity, and cancer (Savino et al., 2016; Khan et al., 2013; Savino et al., 2004; Garcillán et al. 2018). To treat these disorders, some plant-derived substances have been used in clinical practice as adjuvant drugs (Fridlender et al., 2015; Kallifatidis et al., 2016). Furthermore, there have been successful cases of plant-derived chemotherapeutic agents used in cancer treatment, such as Vincristine, Vinblastine, and Taxol (Cragg and Newman, 2013), serving as an inspiration for the development of effective therapies against immune diseases. Based on these arguments, the present study aimed to investigate the immunomodulatory effects of consumption of B. virgilioides stem bark aqueous extract on the functions of thymocytes and B lymphocytes.
MATERIALS AND METHODS
Plant material
The stem bark of B. virgilioides Kunth was collected in March 2006 from the Arboretum of the Federal University of Alagoas, Brazil. The plant had been taxonomically categorized by a botanist of the Alagoas Environment Institute (Instituto de Meio Ambiente de Alagoas, IMA), and a voucher specimen had been deposited in the IMA Herbarium situated in Maceió, Alagoas, Brazil, under the record number MAC29914.
Aqueous extract preparation
Preparation of the aqueous extract was carried out according to the traditional method. The stem bark was dried at ambient temperature and triturated. The aqueous extract of B. virgilioides (AEBv) was prepared by infusing 50 μg of the powdered plant material with 300 ml of boiling water for 20 min. After filtration, the extract was lyophilized by a rotary evaporator until the total solvent evaporated. The yield of the infusion was 17.2%. The extract was reconstituted in 0.9% NaCl solution (vehicle) before the treatments at the required doses.
Animals
C57BL/6 and Swiss mice (4-5 weeks old) were obtained from the breeding colonies of the Federal University of Alagoas. The animals were housed at 22 ± 2°C with a 12 h light/dark cycle and provided with a standard diet of mouse chow and water ad libitum. All the experiments were carried out in accordance with the institutional guidelines and ethics (license number: 010095/2009-67).
AEBv administration
Previous studies have shown that AEBv at least 200 mg/kg was efficacious and potent to induce antinociception and attenuate an allergic inflammatory response (Silva et al., 2010; Silva et al., 2016). Based on these findings, AEBv at the doses of 20 and 200 mg/kg were used to evaluate their effects on the functions of thymocytes and B lymphocytes. Animals were daily gavaged with 20 or 200 mg/kg of AEBv in 0.9% NaCl solution in a final volume of 200 μl for 7 consecutive days. The control group received the same volume of the saline solution (vehicle). After 24 h of the last administration, all the animals were weighed and then euthanized to remove the thymus, mesenteric lymph nodes, liver, and kidney for further analyses. The body weights of the mice were measured daily.
Thymus weight, cellularity, and immunophenotyping
The thymi were removed and their relative weight (organ weight/100 g of the body weight) was determined for each animal. Single thymocyte suspensions were prepared by macerating the thymus with a sterile syringe plunger on Petri dishes in RPMI-1640 medium. The number of the viable thymocytes was counted using a Neubauer’s chamber.
Thymocyte subpopulation phenotyping and VLA-6 expression analyses were performed by flow cytometry following a previously described protocol (Smaniotto et al., 2005). Briefly, thymocytes were incubated with a mixture of anti-CD4/allophycocyanin (APC), anti-CD8/peridinin chlorophyll protein (PercP), and anti-CD49f/phycoerythrin (PE) (BD Pharmingen, San Diego, CA) monoclonal antibodies as well as isotype-matched negative controls for each fluorochrome diluted in 2% fetal calf serum in PBS for 20 min at 4°C. After washing, the cells were fixed and analyzed by flow cytometry using a FACSCanto II device (BD Biosciences). The cell debris and non-viable cells were gated out using the forward versus side scatter parameters. Analyses were made after recording 10.000-50.000 events for each sample.
Intracellular cytokine staining assay
Cytokines expressed by lymphocytes isolated from the mesenteric lymph nodes were examined by intracellular staining according to the manufacturer’s instruction (BD Biosciences, San Diego, CA). Lymphocytes were stained with anti-mouse CD3 (anti-CD3-APC; eBioscience) and anti-B220/CD45R (mouse anti-B220/CD45-PE; eBioscience) antibodies before treatment with the Cytofix/Cytoperm reagent (BD Biosciences). Next, cytokines were intracellularly stained using anti-TNF-α and anti-IL-10 antibodies conjugated to fluorescein isothiocyanate (FITC). The cells were then analyzed by flow cytometry using a FACSCanto II device (BD Biosciences).
Histopathological examination
Thymus, liver, and kidney samples were collected and then immersed in Bouin's solution. After overnight fixation, samples were dehydrated in alcohol, cleared in xylol and embedded in paraffin. They were then sectioned (5 μm thick) and stained with hematoxylin/eosin (HE) and finally observed under a light microscope.
Immunofluorescence staining
Indirect immunofluorescence assay was performed on thymus frozen sections as previously described (Smaniotto et al., 2005). Specimens were incubated with laminin-specific rabbit polyclonal antisera (Novotec, St. Martin-la-Garenne, France) for 1 h at room temperature, washed with PBS, and incubated with the secondary antibody (goat anti-rabbit Ig coupled to FITC) (Biosys. Compiegne, France) for 30 min at room temperature. The samples were examined under a fluorescence microscope (Nikon Eclipse 50i, Nikon Instruments). As a negative control, the primary antibody in some samples was replaced with an unrelated immunoglobulin. Quantitative fluorescence analyses were conducted with the Image J software (National Institute of Health, USA) by transforming the specific staining into pixels and dividing the total pixel numbers by the area analyzed, obtaining the pixel numbers/µm2.
Thymocyte adhesion assay
The cell adhesion assay was performed as previously described with some modifications (Smaniotto et al., 2005). Briefly, 60 × 15 mm culture dishes were coated with 10 μg/ml laminin (Nunc, Copenhagen, Denmark) or bovine serum albumin (BSA, Sigma-Aldrich) as a control at room temperature for 1 h (plus 30 min for air-drying). In these assays, 107 thymocytes derived from three pooled thymi of AEBv-treated C57BL/6 mice or their corresponding controls were let adhere on the precoated dishes for 1 h. The adherent thymocytes were harvested, counted, and analyzed for the expression of CD4 and CD8 surface molecules using flow cytometry.
Thymocyte migration assays
Thymocyte migratory activity was assessed in the transwell system (Mendes-da-Cruz et al., 2014). Briefly, 5 μm pore size Transwell inserts (Costar; Corning, USA) were coated with 10 μg/ml laminin at 37°C for 1 h, following incubation with 10 μg/ml BSA to block any unspecific sites. Then, thymocytes (2.5 × 106) in 100 μl of RPMI with 1% BSA were seeded into the upper chambers. The lower chambers were filled only with the medium. After 3 h of incubation at 37°C in a 5% CO2 humidified atmosphere, migration was evaluated by counting the cells in the lower chambers. The cells were then labelled with the appropriate antibodies and analyzed by using flow cytometry.
Hemagglutination assay
Humoral antibody response was performed as previously described with some modifications (Mahajan and Mehta, 2010). Twenty hours after the 7-consecutive-day oral treatment with 20 mg/kg of AEBv, the animals were immunized with an intraperitoneal injection of 100-μL sheep red blood cells (SRBC; 10%) or saline (control non-immunized). Blood samples were collected from the retro-orbital plexus 5 days later to obtain the serum, and the antibody levels were determined by the hemagglutination technique. The serum was serially diluted in 96-well plates until 1:1024 and incubated with 1% SRBC at room temperature, and hemagglutination was observed after 2 h. The antibody titration was determined according to the last dilution that showed hemagglutination. The data were displayed following the method described by Mahajan and Mehta (2010).
B lymphocyte proliferation assay
Total lymphocytes obtained from the mesenteric lymph nodes of AEBv-treated Swiss mice were cultured in 96-well plates (106 cells/well) and incubated with 2 μg/ml of lipopolysaccharides (LPS, Sigma-Aldrich) in 5% CO2 at 37°C to stimulate the proliferation of B cells as described by Gao et al. (2008). Forty-eight hours later, 5 mg/ml of 3-(4,5-Dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT; Sigma-Aldrich) was added into the cells, which were then maintained for an additional 4 h. After the incubation, the MTT solution was removed and the formazan crystals were solubilized with dimethylsulfoxide. The absorbance was then measured on a microplate reader at 540 nm.
Statistical analysis
Results were expressed as mean ± standard error of the mean (SEM) for 3 animals per group. Mean values were statistically analyzed by ANOVA followed by Tukey’s post-test, or Student's t-test. Values of p < 0.05 were considered statistically significant.
RESULTS
Effect of AEBv on thymic microarchitecture and cellularity
Four-week-old male C57BL/6 mice were orally treated with 20 or 200 mg/kg AEBv for 7 consecutive days. As shown in Figure 1, when compared with the saline-treated mice (control group), the AEBv-treated mice did not show significant changes in thymus microarchitecture, since the capsule, septa, and medullary region in all the studied groups remained the same (Figure 1A-C). Nevertheless, the AEBv-treated mice showed a slight reduction in the cortical region alongside the appearance of spacing between cells (Figure 1D-F).
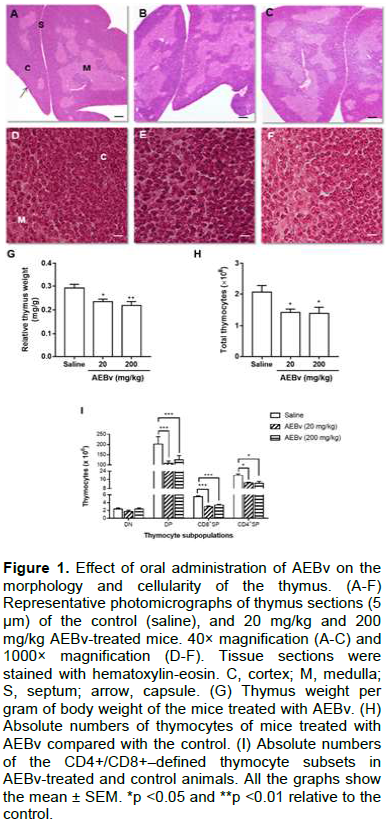
The relative weight of fresh thymus isolated from mice treated with 20 or 200 mg/kg AEBv was significantly less than that of the control thymus (Figure 1G). This difference was reflected with a significant reduction in the total thymocyte numbers following treatment with 20 or 200 mg/kg AEBv compared with the control values (Figure 1H). Interestingly, upon examination of CD4/CD8–defined subpopulations, the AEBv-treated mice were found to have significantly fewer double-positive (DP; CD4+CD8+) and single-positive (CD4SP; CD4+ and CD8SP; CD8+), but not double-negative (DN; CD4-CD8-) thymocytes (Figure 1I).
Effect of AEBv on deposition of laminin and expression of its functional receptor VLA-6 in the thymus
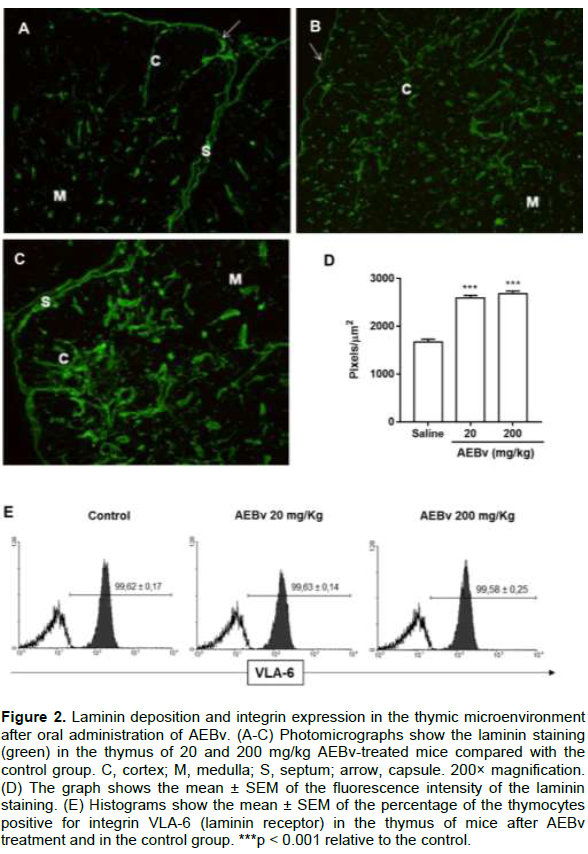
Laminins are glycoproteins that compose the extracellular matrix and have been studied as an essential component for T cell maturation in the thymus (Savino et al., 2015). Thus, it was evaluated whether AEBv treatment could modulate the expression of laminin and its receptor VLA-6 in the thymus. As shown in Figure 2, treatment with 20 and 200 mg/kg AEBv increased laminin deposition in cortical and medullary regions of the thymic lobules compared with the control group (Figure 2A-C). This phenomenon was quantitatively measured by fluorescence intensity in the thymus as a whole (Figure 2D). When evaluating the VLA-6 expression in the thymocytes from AEBv-treated mice, we noted that the treatment did not change the levels of this integrin (Figure 2E).
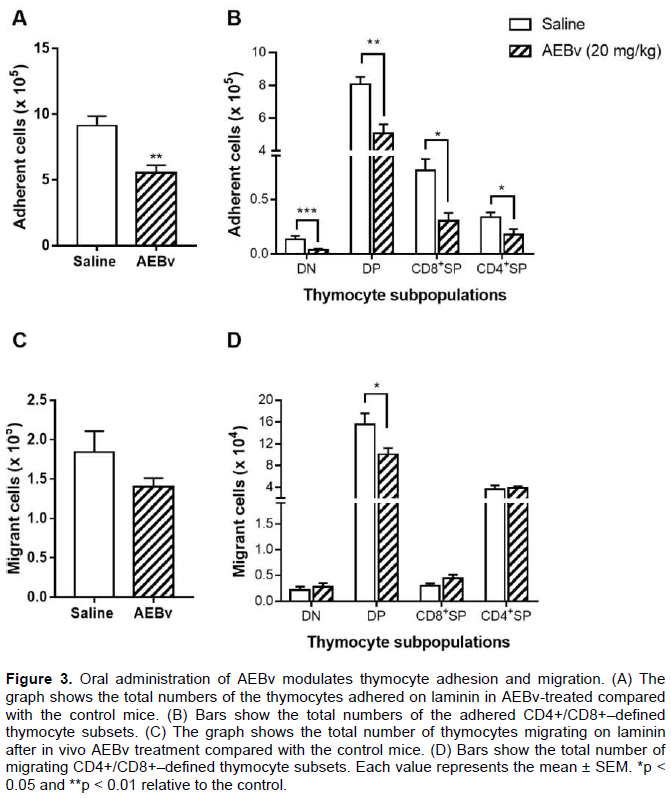
Effect of AEBv on thymocyte adhesion and migration
The results showed above indicated that the AEBv treatment caused changes in the thymic micro-environment, thus, it was investigated whether thymocytes from AEBv-treated mice could exhibit modifications in their adhesive and migratory properties. When the thymocytes from the animals treated with 20 mg/kg dose of AEBv were seeded on the laminin substrate, a significant decrease in the number of adhering thymocytes was found compared with that of the saline-treated mice (Figure 3A). Characterization of this adhesion in each thymocyte subpopulation showed that, compared with the cells from the saline-treated mice, treatment with AEBv caused 73, 37, 60, and 50% reduction in the adhesion of DN, DP, CD4SP, and CD8SP cells, respectively (Figure 3B). Since the adhesion process is required for thymocyte migration during the intrathymic development, the next evaluation was whether the AEBv treatment could also affect their ability to migrate. Using transwell chambers precoated with laminin it was observed a slight decrease in the total number of migrating thymocytes obtained from AEBv-treated mice (Figure 3C). When analyzing the thymocyte subpopulations, only DP thymocytes from AEBv-treated mice had their migratory activity decreased (Figure 3D).
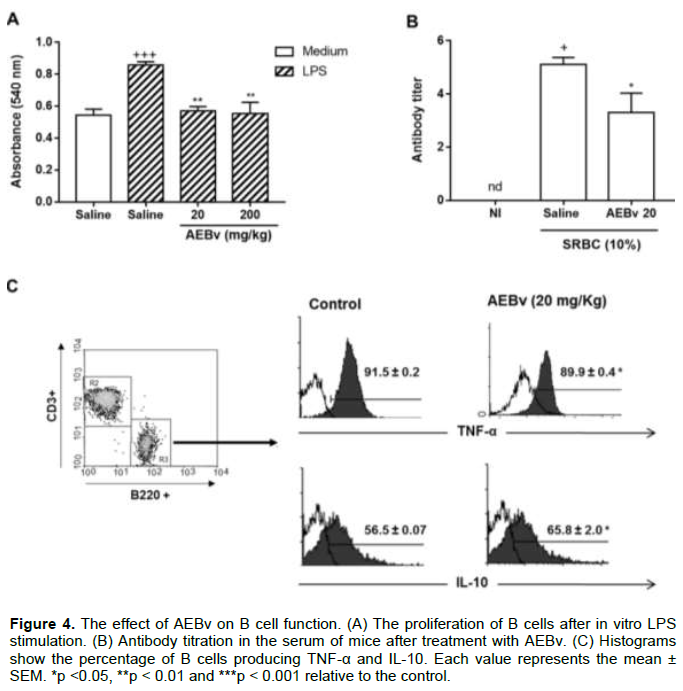
Effect of AEBv on B lymphocyte functions
To address whether AEBv treatment affected other lymphocyte populations, B cells were analyzed in terms of proliferation, production of antibodies, and TNF-α and IL-10 production. As illustrated in Figure 4A, B lymphocytes of the animals treated with saline exhibited an intense proliferative response when stimulated with LPS in vitro in comparison with unstimulated cells. However, when B lymphocytes of the animals treated with 20 or 200 mg/kg of AEBv were exposed to the same LPS stimulation, the proliferative response of B cells was not observed (Figure 4A). Next, to assess whether the antibody production was compromised, the SRBC-specific antibody titer was measured by hemagglutination test. Figure 4B shows that treatment with 20 mg/kg AEBv caused a significant suppression in SRBC-antibody titers compared with the control animals. The basal levels of cytokines in B cells as analyzed by intracellular staining. As shown in Figure 4C, treatment with AEBv induced a significant change in the basal cytokine levels in these cells, evidenced by the reduction in the percentage of TNF-α positive cells and an increase in the percentage of IL-10 positive B cells.
Preliminary toxicity evaluation
Oral administration of AEBv at 20 mg/kg did not produce any mortality. The extract did not produce significant changes in behavior throughout the observation. During the 7 days of treatment, the average daily body weight of the animals treated with 20 mg/kg AEBv (23.0 ± 1.2 g) did not significantly change relative to the average daily body weight of the saline-treated mice (24.8 ± 1.4 g).
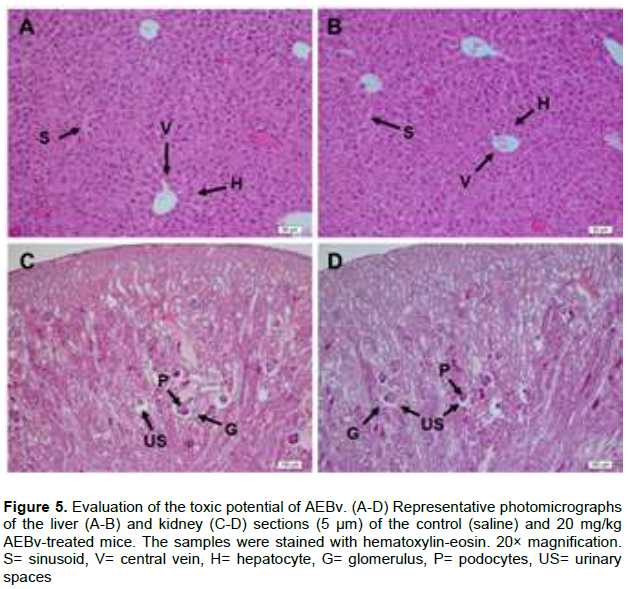
Histopathological analysis of the liver sections from the animals treated with saline (Figure 5A) or with 20 mg/kg AEBv (Figure 5B) showed normal appearance of the central vein and hepatic sinusoids with normal radiating hepatocytes. The kidney sections showed no significant microscopic difference between the mice treated with 20 mg/kg AEBv (Figure 5D) and those treated with saline (Figure 5C). In all the animals, the kidney sections revealed a normal glomerular structure, Bowman’s capsule lined with an outer parietal layer and inner visceral layer/podocytes and preserved urinary spaces (Figure 5C and D).
DISCUSSION
Previous studies have demonstrated that the B. virgilioides stem bark extract (AEBv) suppresses inflammatory responses by both non-allergic and allergic stimuli (Thomazzi et al., 2010; Silva et al., 2010; Barros et al., 2010; Silva, 2016). Nevertheless, its immunomodulatory potential had not been characterized so far. Here, the present results provide evidence that AEBv treatment causes a modulatory effect on the function of thymocytes and B lymphocytes.
In this study, both 7-consecutive-day treatments of 20 and 200 mg/kg AEBv caused an evident thymic weight loss, accompanied by a reduction in absolute thymocyte numbers and an increase in the intercellular space, causing a visible reduction in the cortical region. These findings indicate a possible immunosuppressive effect of the extract. Previous studies with extracts from other medicinal plants that belong to the Fabaceae family have also demonstrated such a potent immunosuppressive effect (Hueza et al., 2007), including a reduction in the number of circulating T lymphocytes (Meng et al., 2009) and intense thymus atrophy accompanied by histological changes (Toro et al., 1992).
Interestingly, the AEBv treatment decreased the double-positive (DP; CD4+CD8+) and single-positive (SP; CD4+ and CD8+), but not the double-negative (DN; CD4-CD8-) cell numbers. This observation clearly indicates that AEBv-induced thymic atrophy was not the result of suppressed trafficking of early lymphoid precursors from the bone marrow to the thymus or due to death of the DN cells. At the same time, these findings suggest that the decline in the DP and SP thymocyte subpopulations may be due to a lack of developmental stimulus for the cells after the DN stage or because of changes in the structural integrity of the organ. It is important to point that the pattern induced by AEBv on the thymocyte subpopulations is distinct from those induced by classical anti-inflammatory drugs, such as steroid medications, since corticosteroid treatments induce thymic atrophy by inducing apoptosis in all thymocyte populations and also in thymic stromal/epithelial cells (Lima et al., 2012). This differential effect of AEBv, affecting later thymocyte maturation, may be due to constituents present in the extract, especially the flavonoids. In fact, previous reports have shown that flavonoids induce marked thymic atrophy accompanied by decreases in relative percentages of DP, CD4SP, and CD8SP thymocytes (Yellayi et al., 2002). In addition, it has been demonstrated that isoflavonoids obtained from B. virgilioides show immunosuppressive activity by inhibiting mitogen-induced T lymphocyte proliferation in vitro (Velozo et al., 1999), reinforcing the immunosuppressive effect of AEBv on the thymus.
It is well established that changes in the composition of the extracellular matrix (ECM) are critical throughout the intrathymic T-cell maturation process (Savino et al., 2015). In the thymus, the ECM forms a structural network composed of molecules that show high binding activity with each other and receptors expressed on the cell surface. Among these ECM components, laminin is heterogeneously distributed in the thymic parenchyma and plays an important role in thymocyte development (Kim et al., 2000). It has been demonstrated that laminin acts in a synergic manner with growth factors and chemokines in the control of intrathymic T-cell migration (Savino et al., 2015). Here, the results confirm previous studies that have shown laminin deposition in the cortical and medullary regions of the thymic lobules (Lannes-Vieira et al., 1991; Ribeiro-Carvalho et al., 2007; Smaniotto et al., 2005). In addition, we showed, for the first time, that the daily consumption of AEBv caused an increase in the deposition of laminin throughout the thymic parenchyma. It is important to emphasize that the augmented deposition of laminin can regulate lymphocyte migration (Simon and Bromberg, 2017), and this may contribute to the immunosuppressive effect of AEBv. However, future studies are required to determine what laminin-producing cell types are sensitive to treatment with AEBv.
It is well described that interactions with ECM components, including laminin, allow adhesion and migration of thymocytes, thereby enabling thymocyte differentiation into more mature subpopulations in the thymus (Ocampo et al., 2008; Savino et al, 2002; Pezzi et al., 2016). In fact, within the thymus microenvironment, the laminin receptor VLA-6 (integrin α6β1) participates in cell adhesion and migration events (Golbert et al., 2013). In our experiments, oral administration of the AEBv did not affect VLA-6 expression in thymocytes. The augmented expression of laminin, but not VLA-6, in thymocytes may impair their physiology to the point that it affects the events necessary for their differentiation, such as adhesion and migration. Similar results have been obtained in thymocytes of mice with high levels of growth hormone (Smaniotto et al., 2005), suggesting that the mechanisms controlling laminin expression in thymocytes are not the same as those involved in VLA-6 expression. To substantiate this hypothesis, future studies are needed to determine the signalling pathways involved in these molecular events.
Considering the changes in laminin deposition and given that intrathymic microenvironment influences thymocyte maturation by affecting adhesion and migration (Savino et al., 2004), the effect of AEBv on thymocyte adhesion and migration was determined. The data revealed that all the thymocyte subpopulations obtained from AEBv-treated mice had decreased adhesive capacity for laminin. Similar cell adhesion results have also been observed in previous studies, in which lymphocyte adhesion was found to be attenuated after treatment with plant-derived products (Rathee et al., 2009). It is worth to mention that cell adhesion plays an essential role in the migration of developing thymocytes through the different thymic compartments (Hu et al., 2015). The results showed that only the DP thymocyte subpopulation from AEBv-treated mice had their migratory ability suppressed. Indeed, there is a broad consensus that thymocyte subpopulations are differentially sensitive to stimuli in a way that DP thymocytes are more susceptible to death than other thymocyte subpopulations (Petrie, 2003). Further studies are required to understand the factors governing the selective DP thymocyte migration in the context of AEBv treatment.
To identify whether treatment with AEBv also affected other lymphocyte populations, the analysis was extended to evaluate the effects of this extract on B-lymphocyte functions. The results revealed that oral treatment with AEBv caused a marked reduction in B-cell proliferation induced by LPS. Consistent with these results, previous studies have already reported that B. virgilioides root extract can inhibit mitogen-induced B cell proliferation (Velozo et al., 1999). During the evaluation of the inflammation-related cytokine profile of B lymphocytes, it was observed that AEBv-treated mice exhibited a reduction in cells producing TNF-α while showing an increase in IL-10–positive cells. These findings indicate that consumption of AEBv may provide a balance between pro- and anti-inflammatory cytokines, which may be a promising strategy in the treatment of inflammatory diseases. The AEBv also decreased the anti-SRBC antibody production. Therefore, it is possible that AEBv may have inhibitory effects on immune-mediated inflammatory diseases in which antibody production is present such as systemic erythematosus lupus (Schelleken et al., 1998; Saha et al., 2011) and asthma (Murdoch and Lioyd, 2010).
Preliminary phytochemical screening of AEBv showed the presence of flavonoids, saponins, and condensed tannins, with low presence of steroids (Silva et al., 2016). It has been shown that oral administration of apple-condensed tannins may ameliorate rheumatoid arthritis by suppressing Th17 cell functions (Nakamura et al., 2015). The pharmacologic and clinical relevance of these data may be important. These results show that AEBv can affect various stages of thymocyte differentiation, and probably, alter the functions of circulating lymphocytes, indicating that AEBv may exert an inhibitory effect specifically on their inflammatory response as in the case with the allergic reactions (Silva et al., 2016).
It is important to evaluate the toxicity of a plant before its use as an herbal medicine (Saad et al., 2005). Here, histological analyses were performed on the liver and kidney to evaluate the effects of AEBv after its oral administration, since mice did not show any signs of systemic toxicity. The treatment with AEBv for 7 days did not cause death, similar to extracts from other species of the Fabaceae family (Bin-Hafeez et al, 2001, 2003), induce aggressive behavior or piloerection, change the body weight, or affect the locomotor activity of the animals. Additionally, no changes were observed in the liver and kidney tissue microarchitecture after AEBv treatment, suggesting that AEBv has low or no toxicity. Based on the outcomes of the present study, it is possible to conclude that AEBv has an immunomodulatory effect, which involves inhibition of distinct functions of both thymocytes and B lymphocytes. Thus, this study results may support this plant's popular use in the treatment of immunological diseases and provide it as a candidate for the isolation of immunomodulatory molecules.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGMENTS
This study was supported by grants from the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq, grant nº 408677/2016-3) and the Fundação de Amparo à Pesquisa do Estado de Alagoas (FAPEAL, grant nº 60030 001260/2017).
REFERENCES
|
Agra IKR, Pires LLS, Carvalho PSM, Silva-Filho EA, Smaniotto S, Barreto E (2013). Evaluation of wound healing and antimicrobial properties of aqueous extract from Bowdichia virgilioides stem barks in mice. Anais da Academia Brasileira de Ciências 85(3):945-954. |
|
|
Alburquerque KA, Guimarães RM, Almeida IF, Clemente ACS (2007). Métodos para superação da dormência em sementes de sucupira-preta (Bowdichia virgilioides Kunth). Cienc Agrotec 6:31. |
|
|
Almeida SP, Proença CEB, Sano SM, Ribeiro JF (1998). Cerrado: espécies vegetais úteis. Planaltina: Embrapa-CPAC. |
|
|
Asadi-Samani M, Bagheri N, Rafieian-Kopaei M, Shirzad H (2017). Inhibition of Th1 and Th17 Cells by Medicinal Plants and Their Derivatives: A Systematic Review. Phytotherapy Research 31(8):1128-1139. |
|
|
Azad R, Babu NK, Gupta AD, Reddanna P (2018). Evaluation of anti-inflammatory and immunomodulatory effects of Premna integrifolia extracts and assay-guided isolation of a COX-2/5-LOX dual inhibitor Fitoterapia 131:189-199. |
|
|
Barros WM, Rao VS, Silva RM, Lima JC, Martins DT (2010). Anti-inflammatory effect of the ethanolic extract from Bowdichia virgilioides H.B.K stem bark. Anais da Academia Brasileira de Ciências 82(3):609-616. |
|
|
Bin-Hafeez B, Ahmad I, Haque R, Raisuddin S (2001). Protective effect of Cassia occidentalis L. on cyclophosphamide-induced suppression of humoral immunity in mice. Journal of Ethnopharmacology 75:13-18. |
|
|
Bin-Hafeez B, Haque R, Pandey S, Sayeed I, Raisuddin S (2003). Immunomodulatory effects of fenugreek (Trigonella foenum graecum L.) extract in mice. International Immunopharmacology 3:257-265. |
|
|
Bui TT, Kwon DA, Choi DW, Jung SY, Lee SY, Piao CH, Hyeon E, Fan Y, Yeon SH, Son RH, Shon DH, Song CH, Shin HS, Chai OH (2019). Rosae multiflorae fructus extract and its four active components alleviate ovalbumin-induced allergic inflammatory responses via regulation of Th1/Th2 imbalance in BALB/c rhinitis mice. Phytomedicine 55:238-248. |
|
|
Chen F, Ye X, Yang Y, Teng T, Li X, Xu S, Ye Y (2015). Proanthocyanidins from the bark of Metasequoia glyptostroboides ameliorate allergic contact dermatitis through directly inhibiting T cells activation and Th1/Th17 responses. Phytomedicine 22(4):510-5. |
|
|
Chen LX, Qi YL, Qi Z, Gao K, Gong RZ, Shao ZJ, Liu SX, Li SS, Sun YS (2019). A Comparative Study on the Effects of Different Parts of Panax ginseng on the Immune Activity of Cyclophosphamide-Induced Immunosuppressed Mice. Molecules 20:24(6). |
|
|
Cragg GM, Newman DJ (2013). Natural products: a continuing source of novel drug leads. Biochimica et Biophysica Acta 1830(6):3670-95. |
|
|
Deharo E, Bourdy G, Quenevo C, Munoz V, Ruiz G, Sauvain M (2001). A search for natural bioactive compounds in Bolivia through a multidisciplinary approach. Part V. Evaluation of the antimalarial activity of plants used by the Tacana indians. Journal of Ethnopharmacology 77:91-98. |
|
|
Fridlender M, Kapulnik Y, Koltai H (2015). Plant derived substances with anti-cancer activity: from folklore to practice. Frontiers in Plant Science 1(6):799. |
|
|
Gao S, Wang Y, Zhang P, Dong Y, Li B (2008). Subacute oral exposure to dibromoacetic acid induced immunotoxicity and apoptosis in the spleen and thymus of the mice. Toxicological Sciences105(2):331-341. |
|
|
Garcillán B, Figgett WA, Infantino S, Lim EX, Mackay F (2018). Molecular control of B-cell homeostasis in health and malignancy. Immunology and Cell Biology 96(5):453-462. |
|
|
Golbert DC, Correa-de-Santana E, Ribeiro-Alves M, de Vasconcelos AT, Savino W (2013). ITGA6 gene silencing by RNA interference modulates the expression of a large number of cell migration-related genes in human thymic epithelial cells. BMC Genomics 14(Suppl 6):S3. |
|
|
Hu Z, Lancaster JN, Ehrlich LI (2015). The Contribution of Chemokines and Migration to the Induction of Central Tolerance in the Thymus. Frontiers in Immunology 7(6):398. |
|
|
Hueza IM, Latorre AO, Raspantini PCF, Raspantini LER, Mariano-Souza DP, Guerra JL, Górniak SL (2007). Effect of Senna occidentalis seeds on immunity in broiler chickens. Journal of Veterinary Medicine 54:179-185. |
|
|
Kallifatidis G, Hoy JJ, Lokeshwar BL (2016). Bioactive natural products for chemoprevention and treatment of castration-resistant prostate cancer. Seminars in Cancer Biology 40-41:160-169. |
|
|
Khan WN, Wright JA, Kleiman E, Boucher JC, Castro I, Clark ES (2013). B-lymphocyte tolerance and effector function in immunity and autoimmunity. Immunologic Research 57(1-3):335-353. |
|
|
Kim MG, Lee G, Lee SK, Lolkema M, Yim J, Hong SH, Schwartz RH (2000). Epithelial cell-specific laminin 5 is required for survival of early thymocytes. Journal of Immunology 165(1):192-201. |
|
|
Lannes-Vieira J, Dardenne M, Savino W (1991). Extracellular matrix componentes of the mouse thymus microenvironment Ontogenetic studies and modulation by glucocorticoids hormones. Journal of Histochemistry and Cytochemistry 39:1539-1546. |
|
|
Le CF, Kailaivasan TH, Chow SC, Abdullah Z, Ling SK, Fang CM (2017). Phytosterols isolated from Clinacanthus nutans induce immunosuppressive activity in murine cells. International Immunopharmacology 44:203-210. |
|
|
Lima AC, Francelin C, Ferrucci DL, Stach-Machado DR, Verinaud L (2012). Thymic alterations induced by Plasmodium berghei: expression of matrix metalloproteinases and their tissue inhibitors. Cellular Immunology 279(1):53-59. |
|
|
Ma B, Kan WL, Zhu H, Li SL, Lin G (2017). Sulfur fumigation reducing systemic exposure of ginsenosides and weakening immunomodulatory activity of ginseng. Journal of Ethnopharmacology 195:222-230. |
|
|
Macêdo MJF, Ribeiro DA, Santos MO, Macêdo DG, Macedo JGF, Almeida BV, Saraiva ME, Lacerda MNS, Souza MMA (2018). Fabaceae medicinal flora with therapeutic potential in Savanna areas in the Chapada do Araripe, Northeastern Brazil. Revista Brasileira de Farmacognosia 28:738-750. |
|
|
Mahajan SG, Mehta AA (2010). Immunosuppressive activity of ethanolic extract of seeds of Moringa oleifera Lam. in experimental immune inflammation. Journal of Ethnopharmacology 130:183-186. |
|
|
Melo FN, Navarro VR, Silva MS, Da-Cunha EV, Barbosa-Filho JM, Braz-Filho R. (2001). Bowdenol, a new 2,3-dihydrobenzofuran constituent from Bowdichia virgilioides. Natural Product Letters 15(4):261-266. |
|
|
Mendes-da-Cruz DA, Brignier AC, Asnafi V, Baleydier F, Messias CV, Lepelletier Y, Bedjaoui N, Renand A, Smaniotto S, Canioni D, Milpied P, Balabanian K, Bousso P, Leprêtre S, Bertrand Y, Dombret H, Ifrah N, Dardenne M, Macintyre E, Savino W, Hermine O (2014). Semaphorin 3F and neuropilin-2 control the migration of human T-cell precursors. PLoS ONE 9(7):e103405. |
|
|
Meng Q, Niu Y, Roubin RH, Hanrahan JR (2009). Ethnobotany, phytochemistry and pharmacology of the genus Caragana used in traditional Chinese medicine. Journal of Ethnopharmacology 124:350-368. |
|
|
Murdoch JR, Lloyd CM (2010). Chronic inflammation and asthma. Mutation Research 690(1-2):24-39. |
|
|
Nakamura K, Matsuoka H, Nakashima S, Kanda T, Nishimaki-Mogami T, Akiyama H (2015). Oral administration of apple condensed tannins delays rheumatoid arthritis development in mice via downregulation of T helper 17 (Th17) cell responses. Molecular Nutrition and Food Research 59(7):1406-1410. |
|
|
Ocampo JS, de Brito JM, Corrêa-de-Santana E, Borojevic R, Villa-Verde DM, Savino W (2008). Laminin-211 controls thymocyte--thymic epithelial cell interactions. Cellular Immunology 254(1):1-9. |
|
|
Petrie HT (2003). Cell migration and the control of post-natal T-cell lymphopoiesis in the thymus. Nature Reviews Immunology, 3(11):859-866. |
|
|
Pezzi N, Assis AF, Cotrim-Sousa LC, Lopes GS, Mosella MS, Lima DS, Bombonato-Prado KF, Passos GA (2016). Aire knockdown in medullary thymic epithelial cells affects Aire protein, deregulates cell adhesion genes and decreases thymocyte interaction. Molecular Immunology 77:157-173. |
|
|
Rathee P, Chaudhary H, Rathee S, Rathee D, Kumar V, Kohli K (2009). Mechanism of action of flavonoids as anti-inflammatory agents: a review. Inflammation and Allergy Drug Targets 8(3):229-235. |
|
|
Ribeiro-Carvalho MM, Lima-Quaresma KRF, Mouço T, Varvalho VF, Mello-Coelho V, Savino W (2007). Triiodothyronine modulates thymocyte migration. Scandinavian Journal of Immunoogy 66:17-25. |
|
|
Saad B, Azaizeh H, Said O (2005). Tradition and perspectives of arab herbal medicine: a review. Evidence-based Complementary and Alternative Medicine 2(4):475-479. |
|
|
Saha S, Tieng A, Pepeljugoski KP, Zandamn-Goddard G, Peeva E (2011). Prolactin, Systemic Lupus Erythematosus, and Autoreactive B Cells: Lessons learnt from murine models. Clinical Reviews in Allergy and Immunology 40:8-15. |
|
|
Savino W, Mendes-da-Cruz DA, Golbert DC, Riederer I, Cotta-de-Almeida V (2015). Laminin-Mediated Interactions in Thymocyte Migration and Development. Frontiers in Immunology 6:579. |
|
|
Savino W, Mendes-da-Cruz DA, Lepletier A, Dardenne M (2016). Hormonal control of T-cell development in health and disease. Nature Reviews Endocrinology 12(2):77-89. |
|
|
Savino W, Mendes-da-Cruz DA, Silva JS, Dardenne M, Cotta-de-Almeida, V (2002). Intrathymic T cell migration: a combinatorial interplay of extracellular matrix and chemokines? Trends in Immunology 23:305-313. |
|
|
Savino W, Mendes-da-Cruz DA, Smaniotto S, Silva-Monteiro E, Villa-Verde DMS (2004). Molecular mechanisms governing thymocyte migration: combined role of chemokines and extracellular matrix. Journal of Leukocyte Biology 75:1-11. |
|
|
Schelleken GA, de Jong BAW, van den Hoogen FHJ, van de Putte LBA, van Venrooij WJ (1998). Citrulline is an essential constituent of antigenic determinants recognized by rheumatoid arthritis-specific autoantibodies. The Journal of Clinical Investigation 101:273-281. |
|
|
Silva JP, Ferro JN, Filho HS, Silva LA, Souza TP, Matos HD, Carvalho VD, Rodarte RS, Barreto E (2016). Aqueous extract of Bowdichia virgilioides stem bark inhibition of allergic inflammation in mice. Journal of Medicinal Plants Research 10:575-584. |
|
|
Silva JP, Rodarte RS, Calheiros AS, Souza CZ, Amendoeira FC, Martins MA, Silva PMR, Frutuoso V, Barreto E (2010). Antinociceptive activity of aqueous extract of Bowdichia virgilioides in mice. Journal of Medicinal Food 13:348- 351. |
|
|
Simon T, Bromberg JS (2017). Regulation of the Immune System by Laminins. Trends in Immunology 38(11):858-871. |
|
|
Smaniotto S, Mello-Coelho V, Villa-Verde DM, Pléau JM, Postel-Vinay MC, Darrdenne M, Savino W (2005). Growth hormone modulates thymocyte development in vivo through a combined action of laminin and CXC chemokine ligand 12. Endocrinology 146:3005-3017. |
|
|
Thomazzi SM, Silva CB, Silveira DCR, Vasconcellos CLC, Lira AF, Cambui EVF, Estevam CS, Antoniolli AR (2010). Antinociceptive and anti-inflammatory activities of Bowdichia virgilioides (sucupira). Journal of Ethnopharmacology 127:451-456. |
|
|
Toro F, Benshimol AL, González EM, Soyano A (1992). Spleen and thymus histology and proliferative response of splenic cells in rats fed raw and cooked Phaseolus vulgaris beans. Archivos Latinoamericanos de Nutrición 42:395-402. |
|
|
Velozo LSM, Silva BP, Bernardo RR, Parente JP (1999). Odoratin-7-O-β-D-glucopyranoside from Bowdichia virgilioides. Phytochemistry 52:1473-1477. |
|
|
Vieira LFA, Reis MDS, Brandão ARA, Viana IMMN, Silva JP, Barreto E, Smaniotto S (2013). Anxiolytic-like effect of the extract from Bowdichia virgilioides in mice. Revista Brasileira de Farmacognosia 23:680-686. |
|
|
Yellayi S, Naaz A, Szewczykowski MA, Sato T, Woods JA, Chang J, Segre M, Allred CD, Helferich WG, Cooke PS (2002). The phytoestrogen genistein induces thymic and immune changes: a human health concern? Proceeding of the National Academy of Science of the United States of America 99(11):7616-7621. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


