Full Length Research Paper

ABSTRACT
Lectins are proteins present in all living beings capable of interacting specifically and reversibly to carbohydrates or glycoconjugates form. They stand out in this family of lectins legumes, which are of special scientific interest because they have different biological functions. This study investigated the presence of a lectin in Brazilian seeds of Mucuna, proceeding with their isolation and evaluation of its antinociceptive effects, anti-inflammatory and antihemolytic activity. Saline extracts of Mucuna pruriens were investigated for the presence of hemagglutinating activity by serial dilution and applied to Sephacryl S-200HR columns and DEAE Sephacel HiPrepFF to purify their lectin. After assessing the purity and molecular weight by polyacrylamide agarose gel electrophoresis (PAGE), the hemolytic activity on human erythrocytes was determined, as well as its antinociceptive action by models of writhing induced by acetic acid and formalin, in addition to its anti-inflammatory activity by testing the paw edema induced by carrageenan. The results indicated that Brazilian Mucuna seeds had a lectin that was purified from seeds having molecular weight of 60.0 kDa and only able to agglutinate erythrocytes of rabbit. The lectin showed an antihemolytic effect on human erythrocytes by not causing hemolysis in these cells compared to the negative control. The administration of lectin in rodents intraperitoneally inhibited the writhing by 99% (2.5 mg/kg), induced by acetic acid, as well as in the method where the formalin nociceptive stimulus was reduced by 51% in stage one and 77% in stage two of the test, using 2.5 mg/kg lectin. The anti-inflammatory activity demonstrated a decrease in paw edema induced by carrageenan in 72% better result than indomethacin control where there was a reduction of only 48% of edema. The study supports the presence of a lectin in Brazilian seeds of M. pruriens with antinociceptive, anti-inflammatory and antihemolytic activities.
Key words: Plant lectin, Mucuna pruriens, purification, antihemolytic, antinociceptive, anti-inflammatory.
INTRODUCTION
Lectins are a heterogeneous group of proteins that share an important biological propriety; they are capable of recognizing specific glycidic structures, interacting in a reversible way (Van Damme et al., 2008). They are found in plants (Silva et al., 2012), vertebrates (Yang et al., 2014), invertebrates (Matsumoto et al., 2012) and microorganisms (Wu et al., 2010), frequently observed on the cell surface or intercellular particles (Lepenies et al., 2013). Among the kingdom Plantae they are abundant in seeds, roots, fruits, leaves and flowers, and are mainly obtained from ripe seeds of legumes in which they comprise up to 15% of total protein and may present one or more molecular forms (Loris et al., 1998).
The specificity of plant lectins concerning different carbohydrates makes possible their application on pharmaceutical research such as: target-cell recognition (Athamna et al., 2006), cellular adhesion, cell interaction, cell-matrix interactions (West and Goldring, 1994), fertilization and agglutination of cells and bacteria (Santi-Gadelha et al., 2012). Although possessing strong similarity on their physicochemical properties and three-dimensional structure, these biomolecules differ on their specificity to the carbohydrate they ligate and their physiological activity (Rego et al., 2002). Lectins already displayed activity on anti-neoplasia (Silva et al., 2014), apoptotic (Zhou et al., 2014), mitogenic (Wong et al., 2010), antibacterial, antifungal and insecticide (Vandenborre et al., 2011), anti-inflammatory (Santi-Gadelha et al., 2006), immunostimulant (Leite et al., 2012), among others. They are considered strong candi-dates for therapeutic use, for they are macromolecules with noticeable resistance to unfavourable conditions like pH and temperature variations and isotonicity, with no significant alterations to their biological function (Coffey et al., 1993).
Mucuna pruriens (L) DC. commonly known as velvet bean is a tropical legume member of the family Fabaceae, widely spread through the south and southeast regions of Asia (Duke, 1981). The seeds have been subject of study especially on their nutritional/ antinutritional contents (Machuka, 2000; Siddhuraju et al., 2000; Siddhuraju et al., 1996), as in India they are a common part of the diet of ethnical groups (Pugalenthi et al., 2005). Yet, there is no literature concerning the bio-chemical and pharmacologic properties that derivate from the lectins present in their seeds. Historically, its use on popular medicine is mainly associated to the treatment of depression, mental disorders and male infertility (Tripathi and Upadhyay, 2002). The M. pruriens seeds present high levels of L-DOPA, a drug applied on the treatment of Parkinson’s, awakening growing interest on research within this species (Dhanasekaran et al., 2008; Lieu et al., 2010; Manyam et al., 2004; Yadav et al., 2013). However, most of the literatures available on the species’ derive from pharmacological studies using aqueous extracts, making specific studies on the isolated bioactive compounds extremely relevant. In the present work, we report the isolation of a lectin present in Brazilian M. pruriens seeds, as well as the evaluation of its antinociceptive, anti-inflammatory and antihemolytic activities.
METHODOLOGY
Plants
The seeds of M. pruriens were collected in the city of João Pessoa, Paraíba, Brazil.
Animal subjects
We used male adult Swiss mice, with average body weight of 25 g, provided by the vivarium of Estácio School of Medicine of Juazeiro do Norte (Estácio/FMJ). The animals were kept on cages with free access to water and food, in a disturbance-free room with constant temperature (24°C) and 12-h light/dark cycle. FMJ’s Institutional Committee of Animal Ethics approved the experimental protocol.
Erythrocytes
We obtained erythrocytes from rabbits of the Prof. George Thomas’ vivarium, annex to the Biotechnology Center of Federal University of Paraíba (UFPB). The experimental protocol to obtain the cell samples was approved by the Institutional Committee of Animal Ethics of UFPB. We obtained human ABO system erythrocytes from expired transfusion bags donated by the state’s blood bank, located in João Pessoa.
Lectin purification (MPLEC)
The whole seeds of M. pruriens were triturated and homogenized (1:10 m/v) via constant agitation during 3 h at 25°C, in NaCl 0.15 M. The extract obtained was then centrifuged for 20 min at 5000 rotation per minute (RPM) and 4°C. The supernatant went through water dialysis (remove NaCl 0,15 M) where the albumins (soluble in water and soluble in saline solutions) and the globulins (insoluble in water but soluble in saline solutions) fractions were separated using centrifugation under the same conditions before. The lyophilized albumin fraction was solubilized (100 mg/ml) in NaCl 0.15 M and applied in a molecular exclusion column Sephacryl S-200 HR HiPrep 26/60 with volume of 320 ml (GE Healthcare) and constant flow rate of 1.3 ml/min linked to AKTAprime plus system (GE Healthcare), the eluted product was monitored at 280 nm. The active peak (with hemagglutinant activity) obtained from the mole-cular exclusion, water-dialyzed, lyophilized and resuspended (40 mg/ml) in Tris-HCl 0.025 M pH 7.6 was applied to a 1 ml ionic-exchange column DEAE Sephacel HiPrep FF 16/10 (GE Healthcare) linked to a AKTAprime plus system (GE Healthcare). The protein fraction was eluted on Tris-HCl 0.025 M pH 7.6 with NaCl 0 to 1 M saline buffer gradient with constant flow rate of 1 ml/min and monitored at 280 nm. The protein was labelled MPLEC (Mucuna pruriens seed lectin). The purification’s electrophoretic protein profile was determined by electrophoresis in polyacrylamide gel according to Laemmli (1970), at non-reductive conditions (PAGE), with the absence of beta-mercaptoethanol and sodium dodecyle sulphate (SDS), with 3% concentration gel and 12.5% separation.
Hemagglutinating activity and inhibition essays
We determined the MPLEC’s hemagglutinating activity (UH) using human ABO and rabbit erythrocytes divided into two groups, processed and not processed by enzymatic treatment of trypsin, bromelain and papain according to Correia and Coelho (1995). UH was defined as the lowest dilution capable of causing hemagglutination. The hemagglutination inhibition essays to determine the lectin specificity were performed in the presence of glycoprotein (fucoidan, fetuin, mucin and λ-carrageenan) at initial concentration of 5 mg/ml and simple sugar carbohydrates (N-acetyl-D-glucosamine, D-glucose, saccharose, L-sorbose, D-fructose, α-lactose, arabinose, xylose, maltose, galactose, D-fucose, methyl-α-D-glucopyranoside, D-trehalose, mannose) at initial concentration of 0.5 M
Protein dosage
The soluble protein dosage was determined according to Bradford (1976), using bovine serum albumin as standard.
Molecular mass determination using PAGE
We determined apparent molecular mass using relative mobility (Mr = distance traversed by the protein compared to the distance traversed by the known molecular mass protein indicators: B phosphorylase 97.0 kDa; BSA, 66.0 kDa; ovalbumin 45.0 kDa; carbonic anhydrase 30.0 kDa; trypsin inhibitor 20.1 kDa; alpha-lactalbumin 14.4 kDa) compared to the relative mobility of MPLEC on PAGE (with the absence of beta-mercaptoethanol and SDS).
Hemolytic activity on human erythrocytes
We followed the protocol by Rangel (1997). Using human erythrocytes of the ABO system diluted in NaCl 0.15 M to achieve a 0.5% suspension, exposed to the lectin at 10, 100 and 1000 mg/ml concentrations. As a positive control, we applied the chemical hemolysant Triton X, and as negative NaCl 0.15 M. The solutions were submitted to agitation at 100 RPM for an hour at 25°C and centrifuged at 2000 RPM for 5 min. We read the liberated hemoglobin on a spectrophotometer at 540 nm. The lectin-induced hemolysis was calculated comparing to the Triton X treatment (100% hemolysis), and compared to the negative control.
Antinociceptive activity
Acetic acid-induced abdominal contortion model
Followed as determined by Vander Wende and Margolin (1956) for rats and modified by Koster et al. (1959)for mice. 30 min after administrating the lectin (MPLEC) via intraperitoneal injection (1 and 2.5 mg/kg), each mouse received an intraperitoneal injection of acetic acid 0.8% diluted in saline (0.1 mg/10 g). During the following 30 min, we counted the abdominal contortions presented by each animal. As positive control a group received Indometacine (Indom – 10 mg/kg, intraperitoneal) a standard analgesic drug, and as negative control we used saline (0.1 ml/10 g). All groups were constituted by seven animals treated independently.
Formalin model
This is done according to Hunskaar and Hole (1987) method. 30 min after administrating the lectin (MPLEC) via intraperitoneal injection (1 and 2.5 mg/kg), each mouse received 40 µl of formalin 1% (v/v, intraplantar) on the hind right foot. After, we registered the time the animal spent licking the injected foot during the initial 5 min (1st phase, neurogenic) and 20 min following (2nd phase, inflammatory). As positive control, a group was treated with Morphine (Mor – 5 mg/kg, intraperitoneal) a standard analgesic drug, and as negative control a group was treated with saline (0.1 ml/10 g). To verify the participation of opioid receptors on the pharmacological effect we used naxolone (Nalox – 2 mg/kg, subcutaneous), an opioid receptor’s antagonist, 15 min before treatment with the lectin of morphine. All groups were constituted by seven animals treated independently.
Anti-inflammatory activity
Carrageenan-induced foot edema model
This was done according to Landucci (1995) method. 30 min after administrating the lectin (MPLEC) via intraperitoneal injection (1 and 2.5 mg/kg), each mouse received an intraplantar injection of 40 µl/foot of a solution of carrageenan 1% p/v on the left hind foot. Foot volume was measured by a plethysmograph (Ugo Basile, Italy), 1, 2, 3, 4, and 24 h after the treatment. Edema volume was calculated by the difference of initial and final foot volume. As positive control we used Indometacin (Indom – 10 mg/kg, intraperitoneal), a standard anti-inflammatory drug, and as a negative control we used a group treated with saline (0.1 ml/10 g, intraperitoneal). All groups were constituted by seven animals treated independently.
Statistical analysis
Results were presented by mean ± standard error of the mean (SEM). To detect statistical difference we applied the analysis of variance (ANOVA), followed by a Turkey’s test, where p < 0.05 (* or #), p < 0.01 (** or ##), p < 0.001 (*** or ###) and p < 0.0001 (**** or ####) were considered statistically significant.
RESULTS AND DISCUSSION
Hemagglutinating activity and carbohydrate inhibition
The albumin fraction of the Mucuna pruriens seeds proved capable of agglutinate rabbit erythrocytes both treated and not treated with proteolytic enzymes (4 UH/ml), but no human erythrocytes agglutination was observed under the same conditions. Mo and Goldstein (1994) observed that the lectin obtained from seeds of M. derringiana was capable of agglutinate rabbit erythro-cytes treated with trypsin. Obochi et al (2007), when characterizing a lectin present in the seeds of M. sloanei observed agglutinating activity using native human ABO system erythrocytes; the same lectin later purified by Teixeira-Sá (2009), who reported human and rabbit ery-throcytes’ agglutination, both treated and not treated with bromelain. Opposing our findings on the hemagglutinating activity, Udedibie and Carlini (1998) reported absence of lectins in Brazilian M. prurensis seeds tested using pig, human and rabbit erythrocytes.
The detection of lectins in M. pruriens seeds seems to be a highly variable factor, being a result of determinant ecological and climatic conditions, as observed on Brazilian and Nigerian M. pruriens seeds var. Utilis (Udedibie and Carlini, 1998). The stable tropical climate in India seems to be favourable to a better expression of lectins in Mucuna seeds (Siddhuraju et al., 2000, 1996), contrasting with the diverse Brazilian climate range, its large territory and geographical physiognomy variation cause significant climatic variation throughout the year, not contributing to an ideal expression of these proteins.
The lectin’s specificity to carbohydrates reported that only the λ-carrageenan was capable of inhibititig the hemagglutinating activity at the concentration of 39.06 µg/ml. Other glycoproteins and simple-sugar carbohy-drates tested could not inhibit the MPLEC activity even at the maximum concentration used (2500 µg/ml for glycoproteins and 250 nM for simple sugars). The results obtained differ from the observed on M. sloanei and M. derringiana seeds’ lectins, both presenting specificity to D-galactose and its derivations (Mo and Goldstein, 1994; Teixeira-Sá et al., 2009). λ-Carrageenan is a sulfated polysaccharide extracted from sea algae, normally constituted by esters of potassium, sodium, calcium, magnesium and ammonium sulfate and repeated units of β-D-galactose-2-sulfate-(1→4)-α-D-galactose-2,6-disulfate. Although reports of the lectins’ specificity to sulphated carbohydrates are scarce, Toda and collaborators (1981) had already observed that Solanum tuberosum and Triticum vulgaris lectins (WGA) are capable to interact with keratan sulfate, a sulfated glycosaminoglycan constituted by repeated units of (1→3)-β-D-galactosyl-(1→4)-β-D-N-acetylglucosamine-6-sulfate.
Due to the probable existence of carrageenan on cell membranes, the MPLEC hemagglutinant activity’s inhibi-tion probably takes place indirectly, through the action of the galactose residuals, hence presenting similarity to the carbohydrate linking of other lectins previously purified. The sulfate groups and the molecule size appear to have great importance on the lectin interaction, as MPLEC did not link to a sepharose CL 4B resin (a galactose polymer – data not demonstrated) nor had its hemagglutinating activity inhibited on the presence of D-galactose. Another unusual fact was observed on the lectins of S. tuberosum and T. vulgaris that demonstrated an enhancing on their inhibition sensibility when the sulfate groups of keratan were removed (Toda et al., 1981).
We highlight the importance of this characteristic observed in MPLEC, as the literature does not report any other vegetal lectin capable of interaction with the carrageenan glycoprotein. Some lectins may display low affinity to monosaccharides or disaccharides due to their active site’s spatial conformation, that demand in some cases more complex molecules such as glycoproteins, which favour an enhanced chemical interaction between the lectin and the carbohydrate (Gabius et al., 2011). The proposition is observed in many lectins isolated from legumes such as the agglutinins from Phaseolus coccineus (Chen et al., 2009), Acacia constricta and Phaseolus vulgaris (Guzman-Partida et al., 2004).
Lectin purification and PAGE
We submitted the albumin protein fraction to molecular exclusion chromatography, resulting in the elution of three distinct protein peaks (PI-ME, PII-ME, PIII-ME); only PI-ME displayed hemagglutination activity on rabbit erythrocytes (Figure 1A). The eluted active peak from the molecular exclusion column went through ionic exchange column, obtaining a peak of protein that was not retained by the matrix and had no agglutinant activity (PI-IE) and a peak of protein retained by the matrix and with agglutinant activity (PII-IE). The latter was eluted in a concentration of 0.85 M of NaCl (Figure 1B). The protein content and specific activity of the purification process can be seen in Table 1. It can be seen that the purifI-cation process resulted in a yield of 80% and an isolation of 32.98 times compared to the total protein of saline extract. These results indicate that the process performed is highly viable in both financial parameters as in final yield. The electrophoretic profile analysis for the active peak obtained by the ionic exchange on PAGE revealed a protein with approximate molecular weight of 60.0 kDa (at native conditions) and a protein pattern apparently pure compared to the albumin fraction of its origin seeds (Figure 2A and B).
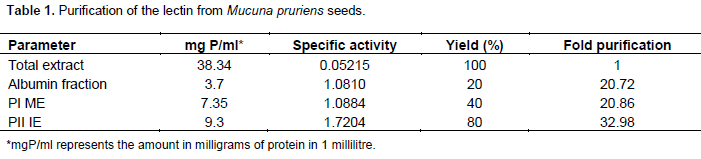
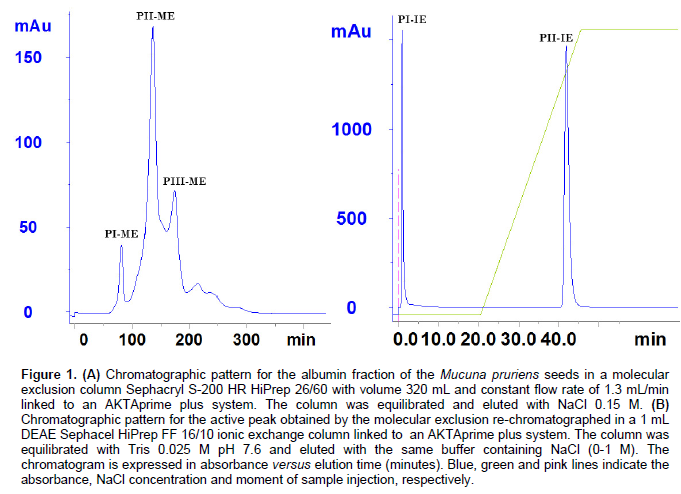
The result presents similarity to what was observed on the lectin obtained from M. sloanei, a protein with ~65.6 kDa molecular weight estimated by molecular filtration chromatography on Superdex 75-HR column (at native conditions) and two bands by SDS-PAGE (36 and 34 kDa) (Teixeira-Sá et al., 2009). There was low similarity with the high molecular weight of the M. derringiana lectin, that presents 90.0 kDa (Mo and Goldstein, 1994), in native conditions. The results presented reflect im-portant characteristics of the Mucuna lectins, displaying high molecular weight, an atypical characteristic for legume lectins, as well as (direct or indirect) affinity for galactose and its structural derivations, hence confirming what was proposed by several authors on the homology of lectins from vegetal species of the same genus and family (Sharon and Lis, 1990). The homology presented in lectins is observed and confirmed by the lectins from the subtribe Diocleinae, in which studies have already been carried out, especially on species of Canavalia, Dioclea and Cratylia. These lectins present specificity for the monosaccharides glycose and mannose, and possess high level of similarity in their amino acid sequences and three-dimensional structure (Loris et al., 2004), yet, they have displayed different biological activit-ies, differing as well on potency and efficiency for the same activities (Cavada et al., 2001; Moreno et al., 2004).
Hemolytic activity on human erythrocytes
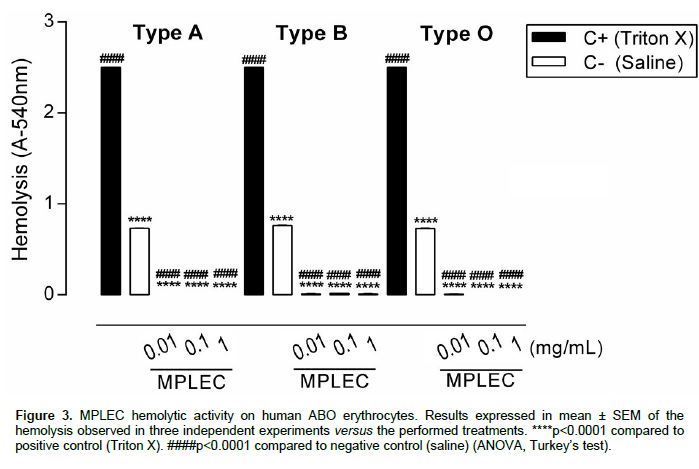
The M. pruriens lectins did not present hemolytic activity
on human erythrocytes. The hemolytic levels observed were lower than the ones demonstrated by saline (negative control), indicating that the lectin probably protected all types of erythrocytes from the natural hemolysis suffered during the experiment (Figure 3). The hemolytic activity profile for erythrocytes is measured based on studies by Rangel et al. (1997), where a substance is considered with low hemolytic effect when it causes the liberation of 0.0 to 40% of the haemoglobin compared to a substance with high hemolytic capacity (over 80% of hemoglobin liberation), such as Triton X. Similar results were found by Leite and collaborators (2012), who evaluated the hemolytic activity of a lectin-like protein present in Clitoria fairchildiana seeds, also registering very low hemolytic effects for human erythro-cytes. In cases like those, it is necessary to evaluate the risk-and-benefit factor for potential pharmacological application. We highlight the importance of these results, as if MPLEC becomes a candidate to therapeutic use, it would only be useful if there were no significant damage to the biological system, especially fundamental structures, such as erythrocytes.
Antinociceptive activity
Acetic acid-induced abdominal contortion model
MPLEC administration by intraperitoneal application (1 and 2.5 mg/kg) 30 min before the acetic acid inhibited the animals’ abdominal contortions (91 and 99% of contortion inhibition, respectively) (Figure 4). The acetic acid is a chemical agent known for its irritant action caused by the ion acetate provoking intracellular acidification, resulting in epithelial cell lesion (Zelitlin and Norris, 1983), inducing the secretion of mediators such as prostaglandins (Ikeda et al., 2001), that activate chemo-sensitive nociceptors causing peripheral inflammatory pain (Julius and Basbaum, 2001). Although it is a test of low specificity, as many non-analgesic drugs (antihistamines, Central Nervous System stimulants, serotoninergic antagonists, neuroleptics and others) may also inhibit the acetic acid-induced contortions (Rates and Barros, 1994), its use as a pharmacological screening procedure still constitutes one of the main models for analgesic activity identification due to its simplicity, execution haste and reduced costs.
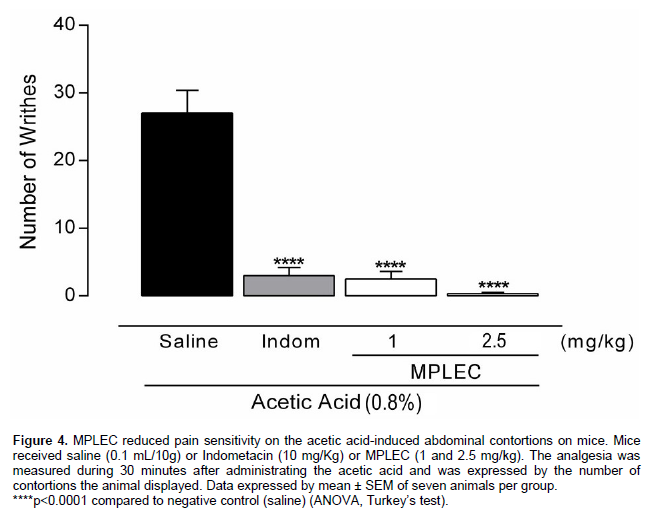
The results obtained by the animal treatment with MPLEC displayed a remarkable pharmacological efficiency compared to the non-steroid anti-inflammatory Indometacin used as test control, as the concentration of 2.5 mg/kg of lectin practically abolished the animals’ contortions. The referred experimental model has been widely applied to verify analgesic activity promoted by lectins present in vegetal species (Holanda et al., 2009) and algae (Silva et al., 2010), as observed in a lectin-like protein purified from Clitoria fairchildiana seeds, in which dose-dependent antinociceptive activity was also reported (Leite et al., 2012). The lectinic site seems to be involved significantly on the lectin-promoted antinocicep-tive activity, as reported in the analgesic agglutinins isolated from C. boliviana seeds and the red algae Pterocladiella capilacea, in which previous association of these lectins with their specific carbohydrates (glycose and mucin) blocked the protein from exert its function against the painful stimulus, resulting in no contortion inhibition (Figueiredo et al., 2009; Silva et al., 2010).
Formalin method
Systemic administration of MPLEC (1 and 2.5 mg/kg) 30 min before the formalin demonstrated significant antinociceptive effect on the foot-licking timespan only at the second lectin concentration tested (2.5 mg/kg), both on initial phase (51% inhibition) and late phase (77% inhibition). As expected, the opioid morphine (5 mg/kg) reduced the nociception significantly in both phases (51% on phase 1, 88% on phase 2), the activity being completely
reversed with the previous administration of the antagonist opioid receptor naloxone (2 mg/kg). No antinociceptive activity reversion observed at the test’s initial phase with previous naxolone administration on the active lectin concentration, but at the late phase partial reversion was observed, yet not significant (Figure 5). The formalin test is considered a more specific model to analgesic drug test, as analgesic drugs present different responses on foot licking timing according to the test phase: the first five minutes (neurogenic phase) are related to drugs that act on opioid receptors system, occurring direct nociceptor stimulation with secretion of substance P, being a Central Nervous System stimulus. The following 20 minutes (inflammatory phase) are linked to anti-inflammatory action (Tjølsen et al., 1992) when chemical mediators, such as amino acids (Tjølsen and Hole, 1997), PGE2 (Malmberg and Yaksh, 1995), histamine (Gaertner et al., 1999), and others are involved, constituting a peripheral nervous system stimulus. It is known that opioid drugs inhibit both phases similarly, and the anti-inflammatories act mainly in the second phase (Hunskaar and Hole, 1987).
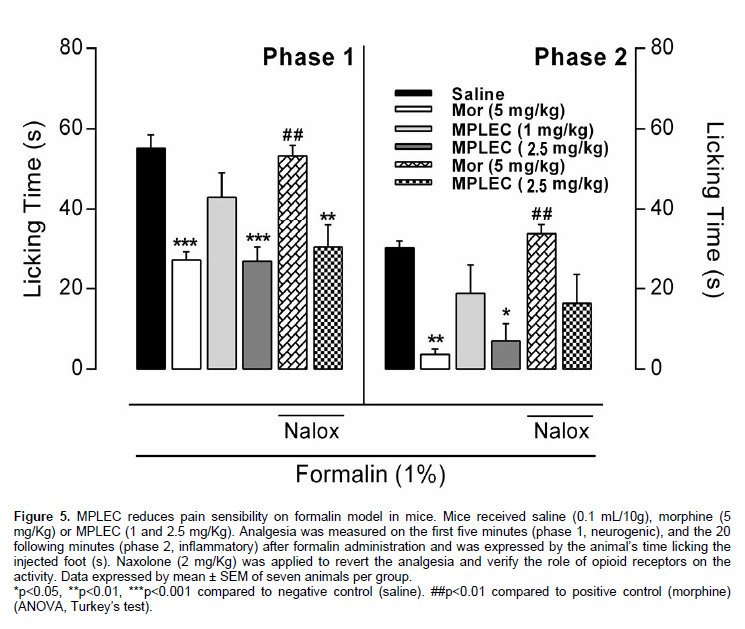
In our study, we could observe inhibitory stimuli in both phases in which the bigger concentration (2.5 mg/kg) was considered active. Probably MPLEC may be acting in a mixed action, but with a stronger anti-inflammatory characteristic as the inhibition was more efficient at the second phase of the test; suggesting that lectin can act both in peripheral and central sites. Such statement is confirmed by the fact that the antinociceptive activity is not completely reverted by naxolone, indicating that the compound is of a different receptor and not the classic opioid receptors. Similar results were observed by Vanderlei and collaborators (2010) on the antinociceptive effects of the lectin extracted from C. cupressoides where the antinociceptive activity was also only partially blocked by naxolone, suggesting a mainly peripheral effect. The effect of lectins on painful stimuli has presented wide variation of stimulated receptors. The algae lectins appear to act mainly on the peripheral nervous system (Bitencourt et al., 2008; Vanderlei et al., 2010), and the legume lectins appear to act at the Central Nervous System (de Freitas Pires et al., 2013; Figueiredo et al., 2009), although it is not a universal claim.
One class of receptor for analgesic exogenous substances, with ongoing studies, is the opioid-like receptors (ORL-1). These receptors are linked to a G protein and present connection with nociception already proven (Higgins et al., 2001; Reinscheid et al., 1995). It is known that these receptors are involved directly on the nociceptive stimuli due to the action of specific antagonists that cause the analgesia, but their action is not blocked by opioid receptor antagonists such as naxolone (Byford et al., 2007). The substances that do
not present naxolone-promoted analgesic action reversion might act directly on ORL-1 receptors; yet, more studies must be held on their specificity, applying drugs that antagonize the activity such as the ones that act on the ATP-dependent potassium channels (glibenclamide), since it is known that the ORL-1 recep-tors are sensitive to those (Armstead, 1999). Hence, if verified the MPLEC activity on this class of receptors, a new target for anesthetic and analgesic drugs is discovered.
Anti-inflammatory activity
The carrageenan caused intense edema that peaked within 3 h on the negative control group (saline) (0.27 ± 0.006) after the administration. The animals treated with MPLEC (1 and 2.5 mg/kg) by intraperitoneal administration 30 min before the carrageenan significantly reduced the edema occurrence when compared with the saline treatment. As expected, the treatment with the anti-inflammatory Indometacin (10 mg/kg), the positive control, also reduced the animals’ edemas (Figure 6A). After the derivation of the points obtained during the experiment in area over the curve distribution with arbitrary units, it was possible to determine the percentage of edema reduction observed. We registered a diminution of 72 and 64% for MPLEC 1 and 2.5 mg/kg, respectively, and 48% for the positive control Indometacin 10 mg/kg, resulting in MPLEC displaying a remarkable pharmacological response compared to the control group, taking into account the dosage applied on the test (Figure 6B). Despite the MPLEC be inhibited by this flogistic agent, a phenomenon occurs so that a higher concentration of carrageenan is necessary or equal to 39.06 mg/ml. The concentration of carrageenan paw edema used in the test was 1% w/v, and the injected volume of 40 µl of the agent in the paw of the animal. These data gave us a value of concentration of 0.0004 µg/ml which is not enough to connect the lectin, and thus block its effect resulting in an anti-inflammatory false positive effect. Thus, the end of the bioavailable fraction agent is capable of causing its pro-inflammatory effect, but is not able to inhibit the lectin injected in doses of 1 and 2.5 mg/kg.
The carrageenan is a polysaccharide extracted from algae and is considered as one of the principal chemical agents of inflammatory processes, with wide application in pharmacological screening tests that evaluate potential anti-inflammatory activity (Hajhashemi et al., 2010; Sadeghi et al., 2011; Whiteley and Dalrymple, 1998). The edema and inflammation caused by carrageenan occur in three distinct phases in which diverse chemical mediators are involved: phase one presents histamine and serotonin liberation, phase two presents cytokine liberation and phase three display the action of prostaglandins (Lo et al., 1982). As presupposed by the antinociceptice activity test, MPLEC has anti-inflammatory activity, with antiedematogenic properties.
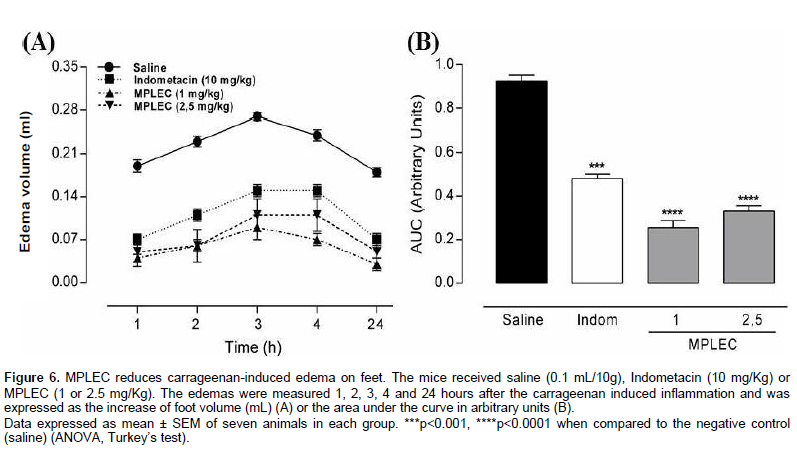
Many physiopathology mechanisms are involved in lectin-mediated anti-inflammatory process. They can inhibit neutrophil migration to the peritoneal cavity and their rolling and vascular adhesion (Mayadas and Cullere, 2005), as well as alter the pro-inflammatory cytokines production (Nunes et al., 2009). More studies are needed to clarify the probable mechanism in which MPLEC exert its function; although many legume lectins that present proven antiedematogenic activity already have their functional pathways partially known, such as the lectin-like protein from C. fairchildiana seeds and the lectin from C. grandiflora, that are capable of inhibiting neutrophil migration and alter the vascular permeability in vivo (Leite et al., 2012; Nunes et al., 2009).
CONCLUSION
A lectin from Brazilian M. pruriens seeds was isolated with 60 kDa mass, capable of agglutinating both native and enzymatically treated rabbit erythrocytes, with its activity inhibited by carrageenan. The lectin possess antinociceptive effect by mixed mechanism, anti-inflammatory and antihemolytic action. More studies are necessary to evaluate its in vivo biological action mechanism and three-dimensional structure.
ACKNOWLEDGEMENTS
The Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) for the financial support given to research. The Federal University of Paraiba and Cell and Molecular Biology Post-Graduate Program.
CONFLICT OF INTEREST
Authors have not declared any conflict of interest.
REFERENCES
|
Armstead WM (1999). Nociceptin/orphanin FQ dilates pial arteries by K+ ATP and K+ Ca2+ channel activation. Brain Res. 835(2):315-323. Crossref |
||||
|
Athamna A, Cohen D, Athamna M, Ofek I, Stavri H (2006). Rapid identification of Mycobacterium species by lectin agglutination. J. Microbiol. Methods 65:209-215. Crossref |
||||
|
Bitencourt FS, Figueiredo JG, Mota MRL, Bezerra CCR, Silvestre PP, Vale MR, Saker-Sampaio S (2008). Antinociceptive and anti-inflammatory effects of a mucin-binding agglutinin isolated from the red marine alga Hypnea cervicornis. Naunyn-Schmiedeberg's Arch. Pharmacol. 377(2):139-148. Crossref |
||||
|
Bradford MM (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72(1):248-254. Crossref |
||||
|
Byford AJ, Anderson A, Jones PS, Palin R, Houghton AK (2007). The hypnotic, lectroencephalographic, and antinociceptive properties of nonpeptide ORL1 receptor agonists after intravenous injection in rodents. Anesth. Analg. 104(1):174-179. Crossref |
||||
|
Cavada BS, Barbosa T, Arruda S, Grangeiro TB, Barral-Netto M (2001). Revisiting proteus: do minor changes in lectin structure matter in biological activity? Lessons from and potential biotechnological uses of the Diocleinae subtribe lectins. Curr. Protein Pept. Sci. 2(2):123-135. Crossref |
||||
|
Chen J, Liu B, Ji N, Zhou J, Bian H, Li C, Bao J (2009). A novel sialic acid-specific lectin from Phaseolus coccineus seeds with potent antineoplastic and antifungal activities. Phytomedicine 16(4):352-360. Crossref |
||||
|
Coffey DG, Uebersax MA, Hosfield GL, Bennink MR (1993). Thermal extrusion and alkali processing of dry beans (Phaseolus vulgaris L.). J. Food Process. Preserv. 16(6):421-431. Crossref |
||||
|
Correia MTS, Coelho LCBB (1995). Purification of a glucose/mannose specific lectin, isoform 1, from seeds of Cratylia mollis mart. (Camaratu Bean). Appl. Biochem. Biotechnol. 55:261-273. Crossref |
||||
|
De Freitas Pires A, Assreuy AMS, Lopes ÉAB, Celedônio NR, Soares CEA, Rodrigues NVFC, Cavada BS (2013). Opioidâ€like antinociceptive effects of oral administration of a lectin purified from the seeds of Canavalia brasiliensis. Fundam. Clin. Pharmacol. 27(2):201-209. Crossref |
||||
|
Dhanasekaran M, Tharakan B, Manyam BV (2008). Antiparkinson drug - Mucuna pruriens shows antioxidant and metal chelating activity. Phytother. Res. 22:6-11. Crossref |
||||
|
Duke JA (1981). Handbook of legumes of world economic importance. Plenum Press. Crossref |
||||
|
Figueiredo JG, Da Silveira Bitencourt F, Beserra IG, Teixeira CS, Luz PB, Bezerra EHS, De Alencar NMN (2009). Antinociceptive activity and toxicology of the lectin from Canavalia boliviana seeds in mice. Naunyn-Schmiedeberg's Arch. Pharmacol. 380:407-414. Crossref |
||||
|
Gabius HJ, André S, Jiménez-Barbero J, Romero A, Solís D (2011). From lectin structure to functional glycomics: principles of the sugar code. Trends Biochem. Sci. 36(6):298-313. Crossref |
||||
|
Gaertner M, Müller L, Roos JF, Cani G, Santos ARS, Niero R, Cechinel-Filho V (1999). Analgesic triterpenes from Sebastiania schottiana roots. Phytomedicine 6(1):41-44. Crossref |
||||
|
Guzman-Partida AM, Robles-Burgueno MR, Ortega-Nieblas M, Vázquez-Moreno I (2004). Purification and characterization of complex carbohydrate specific isolectins from wild legume seeds: Acacia constricta is (vinorama) highly homologous to Phaseolus vulgaris lectins. Biochimie 86(4):335-342. Crossref |
||||
|
Hajhashemi V, Sadeghi H, Minaiyan M, Movahedian A, Talebi A (2010). Central and peripheral anti-inflammatory effects of maprotiline on carrageenan-induced paw edema in rats. Inflamm. Res. 59:1053-1059. Crossref |
||||
|
Higgins GA, Grottick AJ, Ballard TM, Richards JG, Messer J, Takeshima H, Wichmann J (2001). Influence of the selective ORL1 receptor agonist, Ro64-6198, on rodent neurological function. Neuropharmacology 41:97-107. Crossref |
||||
|
Holanda FR, Coelho-de-Sousa AN, Assreuy A, Leal-Cardoso JH, Pires AF, do Nascimento KS, Santos CF (2009). Antinociceptive activity of lectins from Diocleinae seeds on acetic acid-induced writhing test in mice. Protein Pept. Lett. 16(9):1088-1092. Crossref |
||||
|
Hunskaar S, Hole K (1987). The formalin test in mice: dissociation be-tween inflammatory and non-inflammatory pain. Pain 30(1):103-114. Crossref |
||||
|
Ikeda Y, Ueno A, Naraba H, Oh-ishi S (2001). Involvement of vanilloid receptor VR1 and prostanoids in the acid-induced writhing responses of mice. Life Sci. 69(24):2911-2919. Crossref |
||||
|
Julius D, Basbaum AI (2001). Molecular mechanisms of nociception. Nature 413(6852):203-210. Crossref |
||||
| Koster R, Anderson M, De Beer EJ (1959). Acetic acid for analgesic screening. Fed. Proc. 18:412. | ||||
|
Laemmli U (1970). Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227:680-685. Crossref |
||||
|
Landucci ECT, Antunes E, Donato JL, Faro R, Hyslop S, Marangoni S, Nucci G (1995). Inhibition of carrageeninâ€induced rat paw oedema by crotapotin, a polypeptide complexed with phospholipase A2. Br. J. Pharmacol. 114(3):578-583. Crossref |
||||
|
Leite JFM, Assreuy AMS, Mota MRL, Bringel PHSF, e Lacerda RR, Gomes VM, Santi-Gadelha T (2012). Antinociceptive and Anti-inflammatory Effects of a Lectin-Like Substance from Clitoria fairchildiana R. Howard Seeds. Molecules 17(3):3277-3290. Crossref |
||||
|
Lepenies B, Lee J, Sonkaria S (2013). Targeting C-type lectin receptors with multivalent carbohydrate ligands. Adv. Drug Del. Rev. 65:1271-1281. Crossref |
||||
|
Lieu CA, Kunselman AR, Manyam BV, Venkiteswaran K, Subramanian T (2010). A water extract of Mucuna pruriens provides long-term amelioration of parkinsonism with reduced risk for dyskinesias. Parkinsonism Relat. Disord. 16:458-465. Crossref |
||||
|
Lo TN, Almeida AP, Beaven MA (1982). Dextran and carrageenan evoke different inflammatory responses in rat with respect to composition of infiltrates and effect of indomethacin. J. Pharmacol. Exp. Ther. 221(1):261-267. Pubmed |
||||
|
Loris R, Hamelryck T, Bouckaert J, Wyns L (1998). Legume lectin structure. Biochim. Biophys. Acta. 1383:9-36. Crossref |
||||
|
Loris R, Van Walle I, De Greve H, Beeckmans S, Deboeck F, Wyns L, Bouckaert J (2004). Structural Basis of Oligomannose Recognition by the Pterocarpus angolensis Seed Lectin. J. Mol. Biol. 335(5):1227-1240. Crossref |
||||
|
Machuka J (2000). Characterization of the seed proteins of velvet bean Mucuna pruriens from Nigeria. Food Chem. 68(4):421-427. Crossref |
||||
|
Malmberg AB, Yaksh TL (1995). Cyclooxygenase inhibition and the spinal release of prostaglandin E2 and amino acids evoked by paw formalin injection: a microdialysis study in unanesthetized rats. J. Neurosci. 15(4):2768-2776. Pubmed |
||||
|
Manyam BV, Dhanasekaran M, Hare TA (2004). Neuroprotective effects of the antiparkinson drug Mucuna pruriens. Phytother. Res. 18:706-712. Crossref |
||||
|
Matsumoto R, Fujii Y, Kawsar S, Kanaly RA, Yasumitsu H, Koide, Im CH (2012). Cytotoxicity and Glycan-Binding Properties of an 18 kDa Lectin isolated from the Marine Sponge Halichondria okadai. Toxins 4(5):323-338. Crossref |
||||
|
Mayadas TN, Cullere X (2005). Neutrophil beta integrins: Moderators of life or death decisions. Trends Immunol. 26(7):388-395. Crossref |
||||
|
Mo H, Goldstein IJ (1994). Isolation and characterization of a Forssman antigen-binding lectin from velvet bean (Mucuna derringiana) seeds. Glycoconj. J. 11(5):424-431. Crossref |
||||
|
Moreno FBMB, Delatorre P, Freitas BT, Rocha BAM, Souza EP, Faco F, Lima Filho JL (2004). Crystallization and preliminary X-ray diffraction analysis of the lectin from Canavalia gladiata seeds. Acta Crystallogr. D Biol. Crystallogr. 60(Pt 8):1493-5. Crossref |
||||
|
Nunes BS, Rensonnet NS, Dal-Secco D, Vieira SM, Cavada BS, Teixeira EH, Cunha FQ (2009). Lectin extracted from Canavalia grandiflora seeds presents potential anti-inflammatory and analgesic effects. Naunyn-Schmiedeberg's Arch. Pharmacol. 379(6):609-616. Crossref |
||||
| Obochi GO, Malu SP, Effiom EO, Bassey GA (2007). Efficacy and Suitability of Lectin from Mucuna sloanei seeds extracts as a Cell Receptor Signal Inducer. Res. J. Biol. Sci. 2(6):667-669. | ||||
|
Pugalenthi M, Vadivel V, Siddhuraju P (2005). Alternative food/feed perspectives of an underutilized legume Mucuna pruriens var. utilis-a review. Plant Foods Hum. Nutr. 60(4):201-218. Crossref |
||||
|
Rangel M, Malpezzi ELA, Susini SMM, De Freitas J (1997). Hemolytic activity in extracts of the diatom Nitzschia. Toxicon 35(2):305-309. Crossref |
||||
| Rates SMK, Barros HMT (1994). Modelos animais para a avaliação da dor: métodos para triagem de novos analgésicos. Rev. Bras. Farm. 75:31-34. | ||||
|
Rego EJL, de Carvalho DD, Marangoni S, de Oliveira B, Novello JC (2002). Lectins from seeds of Crotalaria pallida (smooth rattlebox). Phytochemistry 60(5):441-446. Crossref |
||||
|
Reinscheid RK, Nothacker HP, Bourson A, Ardati A, Henningsen RA, Bunzow JR, Civelli O (1995). Orphanin FQ: a neuropeptide that activates an opioidlike G protein-coupled receptor. Science 270(5237):792-794. Crossref |
||||
|
Sadeghi H, Hajhashemi V, Minaiyan M, Movahedian A, Talebi A (2011). A study on the mechanisms involving the anti-inflammatory effect of amitriptyline in carrageenan-induced paw edema in rats. Eur. J. Pharmacol. 667:396-401. Crossref |
||||
|
Santi-Gadelha T, Gadelha ACA, Aragão KS, de Oliveira CC, Lima Mota MR, Gomes RC, Cavada BS (2006). Purification and biological effects of Araucaria angustifolia (Araucariaceae) seed lectin. Biochem. Biophys. Res. Commun. 350:1050-1055. Crossref |
||||
|
Santi-Gadelha T, Rocha BAM, Gadelha CAA, Silva HC, Castellon RER, Gonçalves FJT, Cavada BS (2012). Effects of a lectin-like protein isolated from Acacia farnesiana seeds on phytopathogenic bacterial strains and root-knot nematode. Pest. Biochem. Physiol. 103:15-22. Crossref |
||||
|
Sharon N, Lis H (1990). Legume lectins--a large family of homologous proteins. Faseb J. 4(14):3198-208. Pubmed |
||||
|
Siddhuraju P, Becker K, Makkar HPS (2000). Studies on the Nutritional Composition and Antinutritional Factors of Three Different Germ-plasm Seed Materials of an Under-Utilized Tropical Legume, Mucuna p ruriens Var. U tilis. J. Agric. Food Chem. 48(12):6048-6060. Crossref |
||||
|
Siddhuraju P, Vijayakumari K, Janardhanan K (1996). Chemical com-position and protein quality of the little-known legume, velvet bean (Mucuna pruriens (L.) DC.). J. Agric. Food Chem. 44(9):2636-2641. Crossref |
||||
|
Silva LMCM, Lima V, Holanda ML, Pinheiro PG, Rodrigues JAG, Lima MEP, Benevides NMB (2010). Antinociceptive and anti-inflammatory activities of lectin from marine red alga Pterocladiella capillacea. Biol. Pharm. Bull. 33:830-835. Crossref |
||||
|
Silva MCC, De Paula CAA, Ferreira JG, Paredes-Gamero EJ, Vaz AMSF, Sampaio MU, Oliva MLV (2014). Bauhinia forficata lectin (BfL) induces cell death and inhibits integrin-mediated adhesion on MCF7 human breast cancer cells. Biochim. Biophys. Acta 1840:2262-2271. Crossref |
||||
|
Silva MCC, Santana LA, Mentele R, Ferreira RS, de Miranda A, Silva-Lucca RA, Oliva MLV (2012). Purification, primary structure and potential functions of a novel lectin from Bauhinia forficata seeds. Process. Biochem. 47(7):1049-1059. Crossref |
||||
|
Teixeira-Sá D, Reicher F, Braga RC, Beltramini LM, de Azevedo Moreira R (2009). Isolation of a lectin and a galactoxyloglucan from Mucuna sloanei seeds. Phytochemistry 70(17):1965-1972. Crossref |
||||
|
Tjølsen A, Berge OG, Hunskaar S, Rosland JH, Hole K (1992). The formalin test: an evaluation of the method. Pain 51(1):5-17. Crossref |
||||
|
Tjølsen A, Hole K (1997). Animal models of analgesia. In The pharmacology of pain. pp. 1-20. Crossref |
||||
| Toda N, Doi A, Jimbo A, Matsumoto I, Seno N (1981). Interaction of sulfated glycosaminoglycans with lectins. J. Biol. Chem. 256(11):5345-5349. | ||||
|
Tripathi YB, Upadhyay AK (2002). Effect of the alcohol extract of the seeds of Mucuna pruriens on free radicals and oxidative stress in albino rats. Phytother. Res. 16(6):534-538. Crossref |
||||
|
Udedibie ABI, Carlini CR (1998). Brazilian Mucuna pruriens seeds (velvet bean) lack hemagglutinating activity. J. Agric. Food Chem. 46(4):1450-1452. Crossref |
||||
|
Van Damme EJM, Lannoo N, Peumans WJ (2008). Plant lectins. Adv. Bot. Res. 48:107-209. Crossref |
||||
|
Vandenborre G, Smagghe G, Van Damme EJM (2011). Plant lectins as defense proteins against phytophagous insects. Phytochemistry 72(13):1538-1550. Crossref |
||||
| Vander Wende C, Margolin S (1956). Analgesic tests based upon ex-perimentally induced acute abdominal pain in rats. Fed. Proc. 15:494. | ||||
|
Vanderlei ESO, Patoilo KKNR, Lima NA, Lima APS, Rodrigues JAG, Silva LMCM, Benevides NMB (2010). Antinociceptive and anti-inflammatory activities of lectin from the marine green alga Caulerpa cupressoides. Int. Immunopharmacol. 10(9):1113-1118. Crossref |
||||
|
West I, Goldring O (1994). Lectin affinity chromatography. Mol. Biotechnol. 2:147-155. Crossref |
||||
| Whiteley P, Dalrymple S (1998). Models of Inflammation : Carrageenan-Induced Paw Edema in the Rat. Curr. Protocols Pharmacol. 5(4):541-543. | ||||
|
Wong JH, Ng TB, Cheung RCF, Ye XJ, Wang HX, Lam SK, Ngai PHK (2010). Proteins with antifungal properties and other medicinal applications from plants and mushrooms. Appl. Microbiol. Biotechnol. 87(4):1221-1235. Crossref |
||||
|
Wu AM, Gong YP, LiCC, Gilboa-Garber N (2010). Duality of the carbohydrate-recognition system of Pseudomonas aeruginosa II lectin (PA-IIL). FEBS Lett. 584(11):2371-2375. Crossref |
||||
|
Yadav SK, Prakash J, Chouhan S, Singh SP (2013). Mucuna pruriens seed extract reduces oxidative stress in nigrostriatal tissue and improves neurobehavioral activity in paraquat-induced Parkinsonian mouse model. Neurochem. Int. 62:1039-1047. Crossref |
||||
|
Yang L, Bu L, Sun W, Hu L, Zhang S (2014). Functional characterization of mannose-binding lectin in zebra fish: Implication for a lectin-dependent complement system in early embryos. Dev. Comp. Immunol. 46:314-322. Crossref |
||||
| Zelitlin IJ, Norris AA (1983). Mechanism of gastrointestinal inflammation. In A. Standstead (Ed.). Animal model of colitis. p. 70. | ||||
|
Zhou W, Gao Y, Xu S, Yang Z, Xu T (2014). Purification of a mannose-binding lectin Pinellia ternata agglutinin and its induction of apoptosis in Bel-7404 cells. Protein Exp. Purif. 93:11-17. Crossref |
||||
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


