ABSTRACT
Incidence of endocrine disruptors on gonadal development is the cause of many anomalies in the male reproductive system (MRS). Ginkgo biloba is an endocrine disruptor with affinity for β estrogen receptors, which exert an important role in the development of MRS. This study evaluated the effects of G. biloba on the reproductive system of F1 males born to mothers exposed to G. biloba during fetogenesis. Pregnant Wistar rats received, by gavage, 25, 50 or 100 mg/kg/day of G. biloba from the 16th to 20th day of pregnancy. The testicular descent, morphology of the glans penis, sperm concentration, organ weights, testosterone serum concentration, and the structural organization of testicular and epididymal tissues remained unchanged. However, there was an increase in the number of abnormal sperm in all G. biloba-treated groups. These results indicate that G. biloba does not promote maternal toxicity and does not interfere with the fetal and postnatal development and age of puberty onset, but induces the increase of sperm abnormalities.
Key words: Ginkgo biloba, sperm abnormality, fetogenesis, estrogen receptor, rat.
Estrogen plays an important role in testicular development and spermatogenesis. Its effect is modulated via nuclear estrogen receptors (ERs), which have two isoforms: estrogen receptor α (ERα) and estrogen receptor β (ERβ) (Lucas et al., 2010; Hess et al., 2011; Chimento et al., 2014). A new ER - "G protein- coupled estrogen receptor 1" (RSPG) (Prossnitz and Arteburn, 2015) has been identified in germ and somatic cells of rat testes and was reported to mediate the action of estrogen during testicular development and spermatogenesis by triggering rapid signaling events via different pathways (Lucas et al., 2010; Chimento et al., 2014). Knockout male mice strains for estrogen receptors (ERαKO but not ERβKO) or aromatase (ArKO) were shown to exhibit impaired reproductive capacity, reduced
sperm motility, infertility, abnormal spermatogenesis, and dysmorphic testicular structure, indicating the importance of estrogen to male reproductive functions in adults (Carreau and Levallet, 2000; Habert et al., 2006; Hamilton et al., 2014). Furthermore, during fetal development, estrogen deficiency has been reported to increase the number of germ cells and steroidogenic activity (Delbès et al., 2006).
The incidence of abnormalities in the male reproductive system is increasing and can be related to changes induced by endocrine disruptors (EDs) on fetal and gonadal development (Joensen et al., 2008; Divall, 2013). Among the EDs, there are pesticides, plastic-derived materials and natural phytoestrogens, whose presence has already been described in semen, amniotic fluid, umbilical cord and maternal milk (Safe et al., 2010; Jefferson and Williams, 2011; Jones et al., 2014). Phytoestrogens disrupt the reproductive system by mimicking the action of endogenous estrogen when they bind to ERs due to its structural similarity to estradiol (17β-estradiol or E2) (Waring et al., 2008; Patisul and Adewale, 2009; Zhao and Mu, 2011).
The extract of Ginkgo biloba (EGb) has been used for centuries in Asia and Europe for the prevention of several disorders (Sierpina et al., 2003; Dekosky et al., 2008), including dementia, Alzheimer's, and cognitive impairment. It has also been reported to increase cerebral blood flow (Sierpina et al., 2003; Sun et al., 2007; Weinmann, 2010).
The standard extract (EGb 761) has 24% of phytoestrogens, such as kaempferol, quercetin, and ishorhamnetin, which are the main components of the extract (Oh and Chung, 2004). EGb 716 displays estrogenic activity as well as antiestrogenic activity that depend on the concentration of E2 and EGb on the estrogen receptor and its ability to reduce E2 levels by stimulation of catabolism and synthesis inhibition (Oh and Chung, 2006).
Studies related to the reproductive toxicity and developmental toxicity of EGb 761 are scarce. Treatment of mouse blastocysts with gingkolides has been shown to induce apoptosis, decrease the number of cells, decrease post-implantation development, and increase the index of deaths of early blastocysts (Chan, 2005). In the investigation of the mouse oocyte maturation, fertilization, and subsequent embryonic development, oocytes obtained from EGb-treated females following in vitro cultivation and fertilization exhibited decreased oocyte maturation, fertilization and embryo development rate as well as increased embryonic resorption and reduced placental and fetal weight (Chan, 2005). The EGb embryo-fetal developmental toxicity has already been addressed in rats and mice. For instance, administration of EGb by gavage to mice during the 6th to the 15th day of pregnancy did not increase the incidence of malformations and variations or delay in the fetus development (Koch et al., 2013), but intrauterine growth retardation in rat fetuses was noted when EGb was given during the post implantation period (Pinto et al., 2007).
Moreover, the assessment of the postnatal toxicity in the offspring born to female rats treated with EGb during the lactation period did not show alterations in the offspring’s physical, sensory and motor development (de Faria et al., 2008). However, no evidence of effect on the reproductive system of adult F1 male offspring born to mothers treated with EGb in critical periods of development of the male reproductive system was found in the literature.
Fetogenesis is a sensitive and critical period for the development of the male reproductive system, since it establishes the onset of gametogenesis and steroidogenesis (Delbès et al., 2006; Jones et al., 2014). The gonocytes, for instance, proliferate until the 17th post-coital day in rats, remaining quiescent until the 3rd postnatal day (Delbès et al., 2006).
Considering the possibility of using EGb during pregnancy and its effect on the male reproductive system, this study aimed to evaluate the effects of the aqueous extract of G. biloba on the reproductive system of male Wistar rats after maternal exposure during fetogenesis.
Forty female Wistar rats (Rattus norvegicus Berkenhout, 1769) obtained from the vivarium of the Reproduction Biology Center at the Federal University of Juiz de Fora (CBR/UFJF) were used. The animals were 3 months old, nulliparous, and weighed around 175 g.
Pregnant females were housed individually in polypropylene cages (49×34×16 cm) kept in acclimatized shelves (ALESCO®) with airflow, and a controlled temperature of 22 ± 2°C and a 12 h light/dark photoperiod. They were fed on rat chow pellets NUVILAB CR1® (Nuvital Nutrients Ltda., Colombo/PR) and received filtered water ad libitum. The experimental protocol was approved by the Ethics Committee in Animal Experimentation of the Federal University of Juiz de Fora (protocol number 081/2012).
Extract of Ginkgo biloba
G. biloba extract (EGb) was provided by JR Pharma. A quality control test was carried out by Gemini Industry Pharmaceutical Drugs Ltd. and showed the presence of 24% of flavonoids, 6% ginkgolides, and 2.2% of bilobalides.
Experimental design
The guideline for toxicity studies recommends the use of at least three test groups and a control group (OECD 414, 2001). Therefore, in the present study, pregnant females were randomly distributed into four groups, containing 10 animals each, and were treated by gavage, once daily, as follows: control group (C), 1 ml of distilled water, and three groups exposed to Egb (T.Egb) at the dose levels of 25 mg/kg/day (T.Egb 25); 50 mg/kg/day (T.Egb 50) and 100 mg/kg/day (T.Egb 100). The lowest dose was calculated using the body surface area (BSA) as a conversion factor of a daily therapeutic dose from human to rat (FDA, 2005; Reagan-Shaw et al., 2007). The two other doses - 50 and 100 mg/kg/day, correspond to twice and four times the initial recommended dose and follow the guidelines of toxicological studies (OECD, 1995).
The first day of gestation was determined by the presence of sperm in the vaginal smear and treatment was performed during fetogenesis (from the 16th to the 20th day of pregnancy), a time period in which the gonocytes remain quiescent (Delbès et al., 2006). The date of birth was considered the first postnatal day (PND 1) and 72 h after birth, the litters were reduced to four pups per mother (two males and two females) that remained under observation until weaning at PND 25. After weaning, only male pups remained under observation until the day of euthanasia at PND 65. The female pups were used in other studies.
Maternal variables
Female rats were observed daily for the occurrence of clinical signs of toxicity, such as changes in locomotor activity, weight loss, piloerection, diarrhea, stereotypies, and death (OECD, 2009).
Food intake throughout pregnancy was assessed by the difference between a pre-established amount offered to the animals and what was left 24 h later. Body weight was measured every three days from the first day until the beginning of the treatments on the 16th day of pregnancy. During this period, the body weight was measured daily until birth. After delivery, the females were weighed on PND 4, which corresponded to the day of sexing of pups to reduce the number of pups per mother.
Variables of F1 generation
Before weaning, the pups were weighed separately by sex at PND 4, 10, 15, and 20. During the experimental procedure, the occurrence of deaths or cannibalization of pups by the mother was noted. After weaning, all male pups were weighed every five days until PND 65, when the animals were then anesthetized with intraperitoneal xylazine (10 mg/kg) and ketamine (90 mg/kg) (Wolfensohn and Lloyd, 1994) for blood collection by cardiac puncture, following euthanasia by cervical dislocation (CONCEA, 2013).
Testicular descent and differentiation of the glans penis morphology
The animals were monitored daily from DPN 15 until the testicular descent was completed. From DPN 25, the morphological differentiation of the glans penis was also monitored (Yamasaki et al., 2001).
Sperm assessment
After euthanasia, sperm were collected from the epidydimal secretion of the right epididymis cauda. The number of sperm was counted using a hemocytometer with improved double Neubauer ruling in a light microscope BX41TF model (Olympus Tokyo, Japan) under 100x magnification. The total number of spermatozoa was obtained by averaging two counts, corresponding to the upper and lower field in the Neubauer chamber (Seed et al., 1996).
Smears of spermatozoa were stained according to the Shorr method (WHO, 2010) for analysis of sperm morphology in the light microscope BX41TF (Olympus Tokyo, Japan) under 100x magnification. The spermatozoa were classified as normal (intact) or abnormal when displaying: amorphous head (loss of hook shape); no head and/or tail defects (coiled or broken tails and middle piece abnormalities). The evaluation of sperm morphology was made according to the criteria used by Seed et al. (1996) and Perreault and Cancel (2001). Four hundred sperm per animal from 10 animals per group were chosen at random for analysis. The procedure was performed in duplicate per animal.
Weight of organs
Laparotomy was performed after sperm collection for the removal and weighing of the testicles, left epididymis, seminal vesicle (without secretions), kidneys, liver, spleen, and pituitary. All organs were weighed on a precision scale model AV313CP (Ohaus®, Brazil).
Testicular and epididymal histomorphometric analysis and epithelium height
The testis and epididymis were fixed in Bouin. After routine histological processing, they were sectioned at 5 μm thickness in a rotation microtome (Microm®), and were stained with hematoxylin/ eosin and Gomori’s trichrome, respectively. Later, the sections were examined to determine the volumetric proportion of tubular and intertubular testicular compartments, tubular diameter and epithelial height of the seminiferous tubule, and the epitheliumheight of the epididymal duct.
Diameter and epithelial height of the seminiferous tubule: Twenty cross-sections of the seminiferous tubules (most circular ones), from five animals per group, were captured in camera AxioCam (Zeiss) attached to the microscope Primo Star (Zeiss) with 10x objective lens. The greatest and the smallest diameters of each tubule were determined and the average of the two diameters was taken as the final diameter of each measured tubule.
For the calculation of the seminiferous epithelium height, four measures per tubule were made at the same place where the lines for the diameter measurement were positioned. The final height of the seminiferous epithelium comprised the average of these four measurements.
Epithelium height of epididymal duct: Twenty cross sections of the head, body, and tail segments of the epididymis (most circular ones) per animal (n = 4 animals/group) were made as mentioned earlier. Four measures of the duct epithelium height were taken and the average of these values was considered the height of the epithelium of each tubule.
Serum concentration of testosterone
The collected blood was centrifuged at 3500 rpm for 10 min in a RC-3 - Sorval® centrifuge. The serum was aliquoted and stored at -80°C. Thereafter, the serum concentration of testosterone was determined through ELISA immunoassay, using the microplate reader apparatus ASYS HITECH GMBH, and the competitive ELISA kit (Cayman Chemicals®).
Statistical analysis
Homocedastic and normal distribution data were analyzed by one way analysis of variance (ANOVA) followed by post hoc Dunnett test. The measurement data of the testicular diameter were transformed into log base 10 for homogenization. For heteroscedastic and non-normal distribution data, the Kruskal-Wallis-test followed by the Monte Carlo test was used. Values with p<0.05 were considered statistically different. For multiple comparisons, the Monte Carlo test was applied with α = 0.01.
Pregnant animals showed no clinical signs of toxicity during treatment. The average food intake and weight gain of the animals exposed to EGb (T.EGb 25, T.EGb 50, T.EGb 100) and the F1 generation were not significantly different during the experimental procedure when compared with control animals (results not shown).
Testicular descent was completed around DPN 26 in all groups, coinciding with the onset of differentiation of the glans penis morphology. The end of differentiation occurred around DPN 45, which corresponds to the pubertal period, and did not vary among treated and control animals.
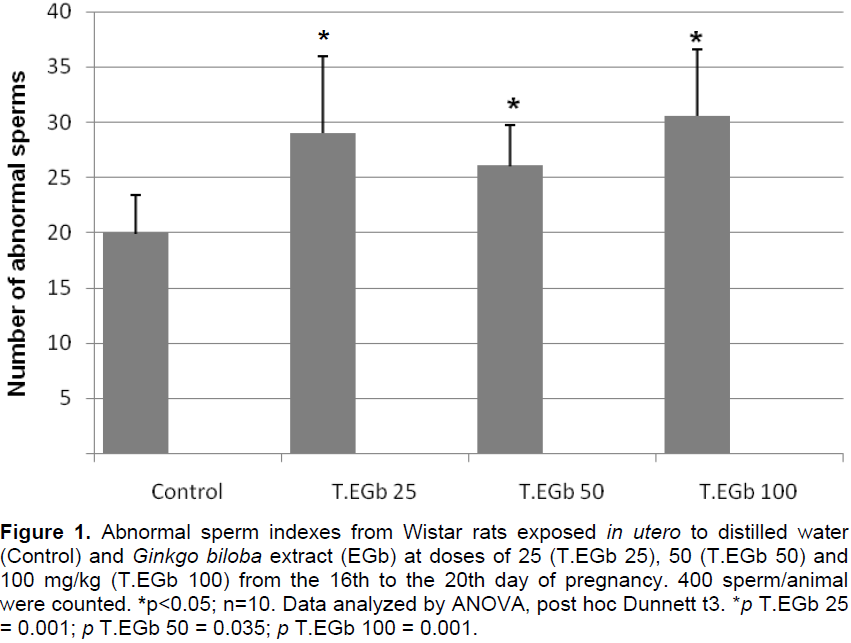
No significant difference in the amount of sperm found in the epididymal tail secretion was observed, however, there was an increase in the number of abnormal sperm in all treated groups in comparison with the control group (Figure 1).
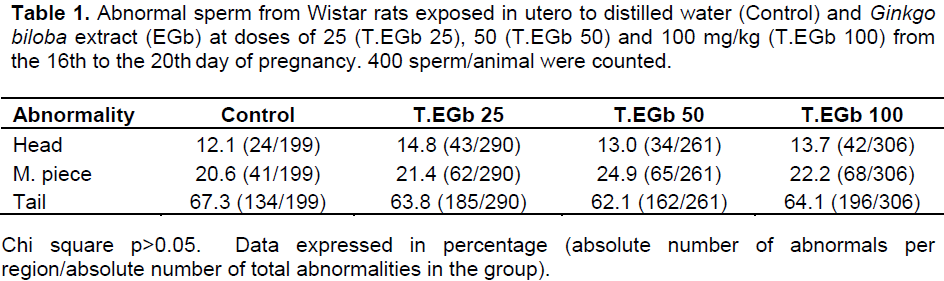
Sperm abnormalities were detected on the head, middle piece, and tail, but the obtained values did not differ significantly among the groups. Broken tail and amorphous head appeared as the most common defects (Table 1).
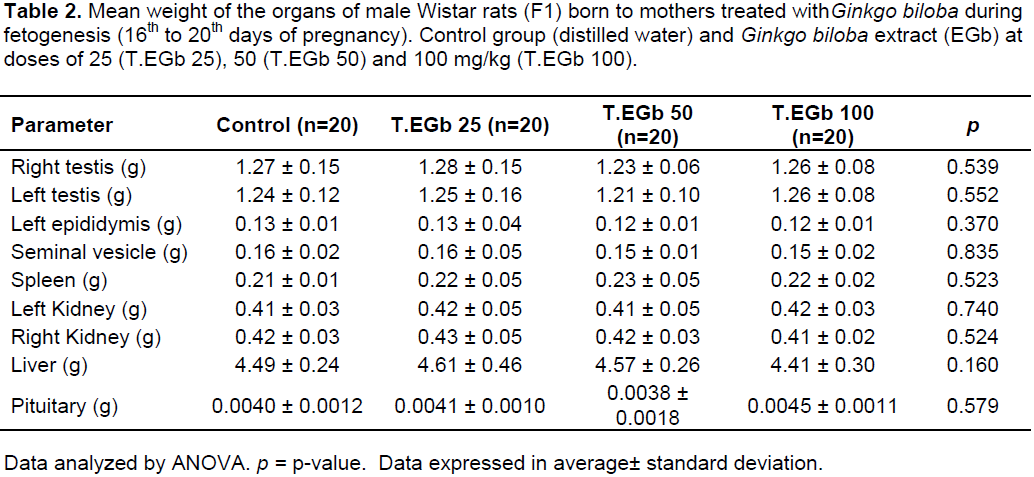
The weight of organs of the reproductive system, the seminal vesicle, pituitary gland, and other organs analyzed were not significantly different between control and treated rats (Table 2), indicating no systemic toxicity due to treatment with G. biloba.
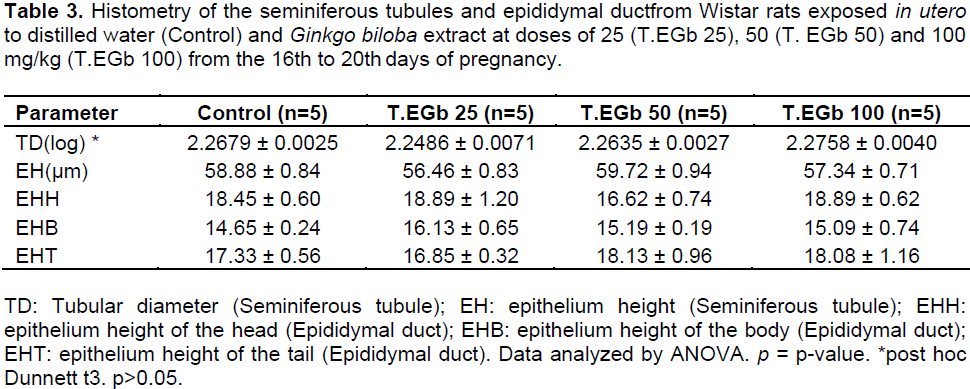
The diameter and height of the epithelium of the seminiferous tubules, and the volumetric proportion of the tubular and intertubular testicular compartments did not differ significantly between the experimental groups (Table 3).
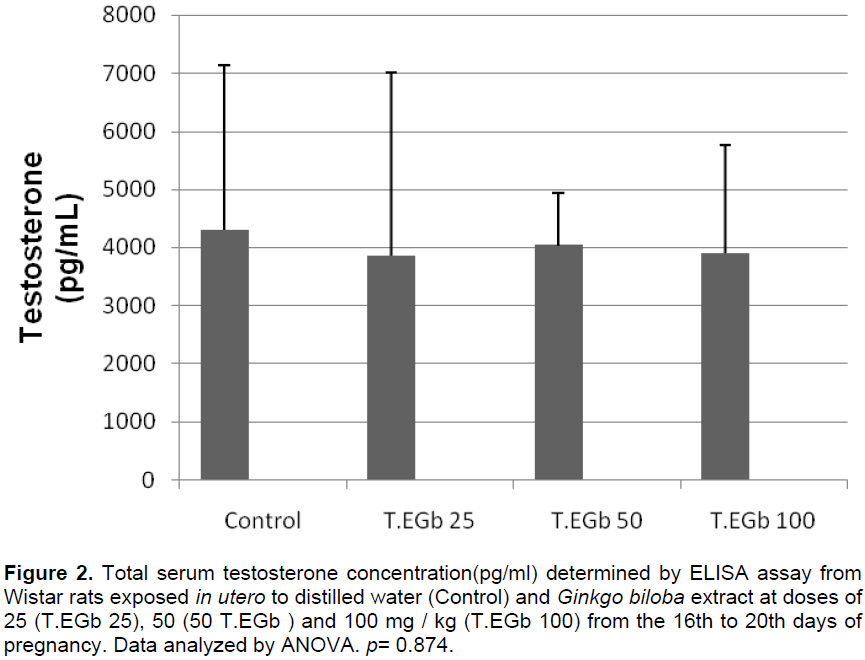
The serum testosterone concentration was largely variable, but there was no significant difference between treated and control values (Figure 2).
Maternal toxicity is an important factor related to embryo-fetal and postnatal development (Khera, 1985). Therefore, whenever the developmental toxicity is the subject of interest in a particular study, it is necessary to eliminate confounding factors, that is, it should be determined if the toxicity in the postnatal development is due to maternal toxicity or a direct effect of the tested substance on the offspring.
Maternal toxicity is observed clinically when there are changes in body weight (Chernoff et al., 2008), reduced locomotion, diarrhea, piloerection and decreased water and food consumption and deaths (Christian, 2001). In this study, none of these clinical signs were noted.
Although it is not known yet if EGb or any of its active constituents can pass through the placenta barrier, the intrauterine growth retardation found in fetuses of rats treated during fetogenesis may be an indicator that the extract or some of its components can reach and affect the fetuses (Pinto et al., 2007).
The variables analyzed in the generation of F1 males in this study cannot be compared to others in the literature, because to the best of our knowledge, this is the first study that evaluated the influence of EGb on male offspring. The F1 generation showed no significant change in body weight or in the weight of the organs examined. This suggests absence of systemic toxicity, since such changes may indicate lesions in the organs systems that lead to physiological impairment (Sellers et al., 2007). These data are also corroborated by the similar growth observed in the litter of the treated and control groups.
In the assessment of reproductive and developmental toxicity, preputial separation and glan penis morphology are among the endpoints in the investigation of abnormal maturation caused by androgenic or antiandrogenic compounds (Stoker et al., 2000; Yamasaki et al., 2001). Yamazaki and collaborators (2001) identified three stages in the normal growth of the rat glans penis which begins with a gradual alteration on the surface of the glans penis from a protuding os penis structure to a W shape (PND 30 - 35), and a subsequent change of the W shape to a flattened surface (PND 39 – 44). Apparently, EGb did not affect the physiological processes related to puberty since both testicular descent and the three stages of differentiation of the glans penis were similar among the groups and similar to the description provided by Yamasaki et al. (2001). These parameters are regarded as the animal’s puberty indicators (Neill et al., 2006) used for the study of endocrine disruptors (Yamasaki et al., 2001). Testicular descent is a prerequisite for the production of mature spermatozoa and can be affected by the action of compounds with estrogenic activity (Klonisch et al., 2004). Although evidence suggests that in utero exposure to such compounds may inhibit testicular descent (Klonisch et al., 2004), in this study such evidence was not observed.
The testicles weight is associated with the number of germ cells present in this organ and the spermatogenic activity (Russell et al., 1990). The weight of the testis did not differ significantly between the control and EGb-treated groups, which is suggestive of absence of changes in the morphology of the seminiferous tubules and sperm production. Similar results were found in studies with adult rats and mice, which showed no changes in the weight of the testis after oral administration of EGb (Al-Yahya et al., 2006; Yeh et al., 2008).
There is a positive correlation between the spermatogenic activity and the height of the seminiferous tubules and the seminiferous epithelium (França and Russell, 1998). Moreover, the diameter of the seminiferous tubule can also providerelevant information about possible alterations in spermatogenesis (Unsal et al., 2006). No changes in the height of the epithelium of the seminiferous tubule were observed in the EGB-treated groups, suggesting that EGb did not change the testicular histologic structure and the spermatogenesis process.
In the epididymis, the epithelium height is a parameter generally used to assess the epithelial secretory and absorptive activity (Hermo and Robaire, 2002). Each region of the epididymis has distinct functions; in particular, the tail compartment is responsible for sperm storage and phagocytosis of abnormal sperm (Robaire and Viger, 1995). This study did not identify significant changes in the height of the epididymal duct epithelium, but showed increased number of abnormal spermatozoa in all groups treated with EGb, which may suggest that the phagocytic process was hindered by the treatment or the number of abnormal spermatozoa superior to the phagocytic capacity of the organ.
Serum testosterone levels are essential for the production of sperm as well as the development of androgen-dependent organs (Grumbach and Conte, 1992). No significant changes were observed in the serum concentration levels of testosterone in the EGB-treated groups when compared with control values. The similar serum testosterone concentrations corroborate the absence of changes in the weights of testosterone-dependent organs, such as theepididymis and seminal vesicle, and the indicators of puberty (testicular descent and morphology of the penis glans).
Although it has been shown that estrogen deficiency during fetal development increased the number of germ cells (Delbès et al., 2006), the obtained results showed that EGb did not alter the sperm concentration, but increased the number of abnormal sperm in all treated groups. The present study did not investigate the causes of these abnormalities. During fetogenesis, the testis begins the gametogenesis and steroidogenesis processes (Delbès et al., 2006; Jones et al., 2014): in the rat, the gonocytes proliferate until the 17th day of pregnancy, remaining quiescent until DPN 3 (Delbès et al., 2006), and from the 16th day of pregnancy, they begin to express the ERβ receptors (Saunders et al., 1998). Considering the administration of EGb to pregnant rats from the 16th to the 20th day of gestation and that EGb has estrogenic or antiestrogenic activity depending on the concentration of E2 and EGb on the estrogen receptor (Oh and Chung, 2006), it is possible to assume that EGB might have interfered with gene expression mechanisms of gonocytes during the period of quiescence, which may have led to the production of abnormal gametes.
In conclusion, the exposure of Wistar rats to EGb during fetogenesis, a critical period of the male reproductive system development, did not seem to cause maternal toxicity or alterations on fetal and postnatal development, and on the onset of puberty. Nevertheless, an increased number of abnormal sperm in the adult offspring was noted.
The authors have not declared any conflict of interests.
This work received the support of Rede Mineira de Bioterismo/FAPEMIG 31/11 and TOXIFAR 26/11 and CNPq (“Conselho Nacional de Desenvolvimento Científico e Tecnológico - Brasil"). The authors are thankful to Kyle Caçador Florence and Sergio Caçador Florence for assisting with the English version of the article, to Leonardo Toshio Oshio for teaching the techniques used in the work methodology, and to João Evangelista de Paula Reis for the supply of EGb.
REFERENCES
|
Al-Yahya AA, Al-Majed AA, Al-Bekairi AM, Al-Shabanah OA, Qureshi S (2006). Studies on the reproductive, cytological and biochemical toxicity of Ginkgo biloba in Swiss albino mice. J. Ethnopharmacol. 107(2):222-8.
crossref
|
|
|
|
Carreau S, Levallet J (2000). Testicular estrogens and male reproduction. Physiology 15(4):195-198.
|
|
|
|
Chan WH (2005). Ginkgolides induce apoptosis and decrease cell numbers in mouse blastocysts. Biochem. Biophys. Res. Commun. 338(2):1263-7.
crossref
|
|
|
|
Chernoff N, Rogers EH, Gage MI, Francis BM (2008). The relationship of maternal and fetal toxicity in developmental toxicology bioassays with notes on the biological significance of the "no observed adverse effect level". Reprod. Toxicol. 25(2):192-202.
crossref
|
|
|
|
Chimento A, Sirianni R, Casaburi I, Pezzi V (2014). GPER signaling in spermatogenesis and testicular tumors. Front. Endocrinol. (Lausanne). 5:1-9.
crossref
|
|
|
|
Christian MS (2001). Test Methods for assessing female reproductive and development toxicology. In: Hayes AW (eds.), Principles and methods of toxicology. Philadelphia: Taylor & Francis. pp. 1301-1381.
|
|
|
|
Conselho Nacional de Controle de Experimentação Animal (CONCEA) (2013). Diretrizes da prática de eutanásia do CONCEA. Brasília: Ministério da Ciência Tecnologia e Inovação. 54 p.
|
|
|
|
Dekosky ST, Williamson JD, Fitzpatrick AL, Kronmal RA, Ives DG, Saxton JA, Lopez OL, Burke G, Carlson MC, Fried LP, Kuller LH, Robbins JA, Tracy RP, Woolard NF, Dunn L, Snitz BE, Nahin RL, Furberg CD (2008). Ginkgo biloba for prevention of dementia: a randomized controlled trial. JAMA 300(19):2253-2262.
crossref
|
|
|
|
Delbès G, Levacher C, Habert R (2006). Estrogen effects on fetal and neonatal testicular development. Reproduction 132(4):527-38.
crossref
|
|
|
|
Divall AS (2013). The influence of endocrine disruptors on growth and development of children. Curr. Opin. Endocrinol. Diabetes Obes. 20(1):50-5.
crossref
|
|
|
|
de Faria DE, Borges LV, Peters VM, Reis JE, Ribeiro LC, de Cássia da Silveira E Sá R, Guerra Mde O (2008). Postnatal development of pups from nursing rats treated with Ginkgo biloba. Phytother. Res. 22:185-189.
crossref
|
|
|
|
Food and Drug Administration (FDA) (2005). Guidance for Industry: Estimating the maximum safe dose in initial clinical trials for therapeutics in adult healthy volunteers. Available at: view
|
|
|
|
França LR, Russell LD (1998). The testis of domestic mammals. In: Martinezgarcia F, Regadera J (Eds). Male reproduction – a multidisciplinary overview. Madrid: Churchill Communications. pp. 198-219.
|
|
|
|
Grumbach MM, Conte FA (1992). Disorders of sex differentiation. In: Wilson JD, Foster DW (Eds). Williams Textbook of Endocrinology. Philadelphia: Saunders. pp. 853-951.
|
|
|
|
Habert R, Delbes G, Duquenne C, Livera G, Levacher C (2006). Effets des estrogènes sur le développement du testicule pendant la vie foétale et néonatale. Gynecol. Obstet. Fertil. 34(10):970-977.
crossref
|
|
|
|
Hamilton KJ, Arao Y, Korach KS (2014). Estrogen hormone physiology: reproductive findings from estrogen receptor mutant mice. Reprod. Biol. 14(1):3-8.
crossref
|
|
|
|
Hermo L, Robaire B (2002). Epididymal cell types and their functions. In The Epididymis: From Molecules to Clinical Practice. New York:
crossref
|
|
|
|
Hess RA, Fernandes SA, Gomes GR, Oliveira CA, Lazari MF, Porto CS (2011). Estrogen and its receptors in efferent ductules and epididymis. J. Androl. 32(6):600-13.
crossref
|
|
|
|
Jefferson WN, Williams CJ (2011). Circulation levels of genistein in the 1088 J. Med. Plants Res. neonate, apart from dose and route, predict future adverse female reproductive outcomes. Reprod. Toxicol. 31(3):272-279.
crossref
|
|
|
|
Joensen UN, Jorgensen N, Rajpert-De Metys E, Skakkebaek NE (2008). Testicular dysgenesis syndrome and Leydig cell function. Basic Clin. Pharmacol. Toxicol. 102:155-161.
crossref
|
|
|
|
Jones S, Boisvert A, Buong TB, Francois S, Thrane P, Culty M (2014). Disruption of rat testis development following combined in utero exposure to the phytoestrogen genistein and antiandrogenic plasticizer Di-(2-Ethylhexyl) phthalate. Biol. Reprod. 91:1-14.
crossref
|
|
|
|
Khera KS (1985). Maternal toxicity: a possible etiological factor in embryofetal deaths and fetal malformations of rodent-rabbit species. Teratology 31(1):129-53.
crossref
|
|
|
|
Klonisch T, Fowler PA, Hombach-Klonisch S (2004). Molecular and genetic regulation of testis descent and external genitalia development. Dev. Biol. 270:1-18.
crossref
|
|
|
|
Koch E, Noldner M, Leuschner J (2013). Reproductive and developmental toxicity of the Ginkgo biloba special extract EGb 761® in mice. Phytomedicine 21:90-7.
crossref
|
|
|
|
Lucas TFG, Royer C, Siu ER, Lazari MFM, Porto CS (2010). Expression and signaling of G protein-coupled estrogen receptor 1 (GPER) in rat Sertoli cells. Biol. Reprod. 83:307-317.
crossref
|
|
|
|
Neill JD, Plant TM, Pfaff DW, Challis JRG, Kretser DM, Richards JS, Wassarman PM (2006). Knobil and Neill's Physiology of Reproduction. In: Ojeda SR, Skinner MK (eds) Puberty in the rat. 3rd ed. Elsevier. pp. 2061-2126.
|
|
|
|
Organisation for Economic Co-operation and Development (OECD) (2001). Test Guideline 414. OECD Test Guideline for Testing of Chemicals. Prenatal Developmental Toxicity Study.
view
|
|
|
|
Organisation for Economic Co-operation and Development (OECD) (2009). Test Guideline 452. OECD Test Guideline for Testing of Chemicals. Chronic Toxicity Studies.
view
|
|
Oh SM, Chung KH (2006). Antiestrogenic activities of Ginkgo biloba extracts. J. Steroid Biochem. Mol. Biol. 100(4-5):167-76.
crossref
|
|
|
|
Oh SM, Chung KH (2004). Estrogenic activities of Ginkgo biloba extracts. Life Sci. 74: 1325-1335. Life Sci. 74(11):1325-35.
crossref
|
|
|
|
Patisul HB, Adewale HB (2009). Long-term effects of environmental endocrine disruptors on reproductive physiology and behavior. Front. Behav. Neurosci. 3:10.
crossref
|
|
|
|
Perreault SD, Cancel AM (2001). Significance of incorporating measures of sperm production and function into rat toxicology studies. Reproduction 121(2):207-16.
crossref
|
|
|
|
Pinto RM, Fernandes ES, Reis JE, Peters VM, Guerra Mde O (2007). Intra-uterine growth retardation after prenatal administration of Ginkgo biloba to rats. Reprod. Toxicol. 23(4):480-5.
crossref
|
|
|
|
Prossnitz ER, Arterburn JB (2015). International union of basic and clinical pharmacology. XCVII. G Protein–coupledestrogen receptor and its pharmacologic modulators. Pharmacol. Rev. 67(3):505-540.
crossref
|
|
|
|
Reagan-Shaw S, Nihal M, Ahmad N (2007). Dose translation from animal to human studies revisited. FASEB J. 22(3):659-61.
crossref
|
|
|
|
Robaire B, Viger RS (1995). Regulation of epididymal epithelial cell functions. Biol. Reprod. 52:226-236.
crossref
|
|
|
|
Russell LD, Ettlin R, Sinha APH, Clegg ED (1990). Histological and histopathological evaluation of the testis, 1st ed. Clearwater: Cache River Press. 286 p.
|
|
|
|
Safe S, Jutoorua I, Chadalapakaa G (2010). Estrogenic endocrine disruptors: molecular characteristics and human impacts. Compr. Toxicol. 2:609-621.
crossref
|
|
|
|
Saunders P, Fisher J, Sharpe R, Millar M (1998). Expression of oestrogen receptor beta (ER beta) occurs in multiple cell types, including some germ cells, in the rat testis. J. Endocrinol. 156:13-17.
crossref
|
|
|
|
Seed J, Chapin RE, Cleeg ED, Dostal LA, Foote RH, Hurtt ME, Klinefelter GR, Makris SL, Perreault, SD, Schrader S, Seyler D, Sprano R, Treinen KA, Veeramachaneni DNR, Wise LD (1996). Methods for assessing sperm motility, morphology, and counts in the rat, rabbit, and dog: a consensus report. Reprod. Toxicol. 10: 237-244.
crossref
|
|
|
|
Sellers RS, Morton D, Michael B, Roome N, Johnson JK, Yano BL, Perry R, Schafer K (2007). Society of Toxicologic Pathology Position Paper: Organweight recommendations for toxicology studies. Toxicol. Pathol. 35:751-755.
crossref
|
|
|
|
Sierpina VS, Wollschlaeger B, Blumenthal M (2003). Ginkgo biloba. Am. Fam. Physician 68(5):923-6.
Pubmed
|
|
|
|
Stoker TE, Parks LG, Gray LE, Cooper PL (2000). Endocrine-disrupting chemicals: prepubertal exposures, and effects on sexual maturation, and thyroid function in the male rat. A focus on the EDSTAC recommendations. Crit. Rev. Toxicol. 30:197-252.
crossref
|
|
|
|
Sun BL, Yuan H, Xia ZL, Zhang SM, Wang LX (2007). Effects of extract of Ginkgo biloba on intracranial pressure, cerebral perfusion pressure, and cerebral blood flow in a rat model of subarachnoid hemorrhage. Int. J. Neurosci. 117(5):655-65.
crossref
|
|
|
|
Unsal A, Muzaffer E, Avci A, Cimentepe E, Guven C, Balbay MD, Durak I (2006). Protective role of natural antioxidant supplementation on testicular tissue after testicular torsion and detorsion. Scand. J. Urol. Nephrol. 40:17-22.
crossref
|
|
|
|
Waring RH, Ayers S, Gescher AJ, Glatt HR, Meinl W, Jarrat P, Kirk CJ, Pettitt T, Rea D, Harris RM (2008). Phytoestrogens and xenoestrogens: the contribution of diet and environment to endocrine disruption. J. Steroid Biochem. Mol. Biol. 108:213-220.
crossref
|
|
|
|
Weinmann S, Roll S, Schwarzbach C, Vauth C, Willich SN (2010). Effects of Ginkgo biloba in dementia: systematic review and meta-analysis. BMC Geriatr. 10:14.
crossref
|
|
|
|
World Health Organization (WHO) (2010). WHO laboratory manual for the examination and processing of human semen. 5. ed. Geneva: WHO Press, 287p.
|
|
|
|
Wolfensohn S, Lloyd M (1994). Handbook of laboratory animal management and welfare. New York: Oxford University Press Inc. 304p.
|
|
|
|
Yamasaki K, Sawaki M, Noda S, Muroi T, Takatsuki M (2001). Preputial separation and glans penis changes in normal growing Crj: CD (SD) IGS rats. Reprod. Toxicol. 15:533-536.
crossref
|
|
|
|
Yeh KY, Pu HF, Kaphle K, Lin SF, Wu LS, Lin JH, Tsai YF (2008). Ginkgo biloba extract enhances male copulatory behaviour and reduces serum prolactin levels in rats. Horm. Behav. 53(1):225-231.
crossref
|
|
Zhao E, Mu Q (2011). Phytoestrogen biological actions on mammalian reproductive system and cancer growth. Sci. Pharm. 79:1-20.
crossref
|