Full Length Research Paper

ABSTRACT
Leishmaniasis is the second-largest parasitic killer in the world (after malaria). Currently, there are no effective vaccines or therapeutic interventions thus creating a gap for new alternative remedies in the management of the disease. Medicinal plants offer this attractive option as antileishmanial agents since they are relatively safe, locally available and affordable. The study assessed antileishmanial activity of methanol, hexane, dichloromethane and ethyl acetate extracts, in addition to the aqueous phase of Bidens pilosa, Tithonia diversifolia and Sonchus oleraceus on Leishmania major parasites as well as their phytochemical composition. The phytochemical screening of the plants showed presence of alkaloids, saponins, terpenoids, anthraquinones, glycosides, tannins and flavonoids. Cytotoxicity studies on Vero cells showed a low toxicity with IC50 more than 0.12 mg/ml. L. major viability was determined at various concentrations of the extracts and standard (Pentostam) using MTT reagent and results displayed as IC50. The extracts from S. oleraceus, B. pilosa, and T. diversifolia displayed IC50 values from 0.03 to 4.89 mg/ml on Leishmania parasites. Among the three plants extracts, dichloromethane extract had the highest activity with IC50 0.03 mg/ml. As indicated, all the extracts demonstrated a great potential in inhibiting the growth of Leishmania promatigotes with low toxicity making them good candidates for compound isolation and drug manufacture.
Key words: Vero cells, MTT, cytotoxicity, promastigotes, Sonchus oleraceus, Bidens pilosa, Tithonia diversifolia
INTRODUCTION
Leishmaniasis refers to a broad spectrum disease brought by intracellular Leishmania parasites. The disease is transmitted by bites from infected female Phlebotominae sand fly (Mutoro et al., 2018b; De Souza et al., 2016) and results into various types ranging from self-recuperating lesions, muco-cutaneous and fatal visceral form of the disease (Mutoro et al., 2018a). This parasite has been known to kill thousands and debilitates millions of people each year (De Queiroz et al., 2014). Sadly, it remains one of the most neglected diseases with new cases at seven hundred thousand to one million (700,000- 1,000,000) and twenty thousand to thirty thousand (20,000-30,000) annual deaths for the various forms of the disease (WHO, 2018). According to World Health Organization (WHO)’s report in 2017, Leishmaniasis is found in 88 countries; Latin America, Africa and other regions associated with low income. Majority of the new cases of the disease occurrence are found in South Sudan, Brazil, Sudan Somalia and Kenya. Burza et al. (2018), record that over 50% of the people suffering from the disease in India, Sudan and Brazil are children. According to Al-Hajj et al. (2018), Leishmaniasis remains a neglected tropical disease despite increased disease burden and death cases as there are no effective vaccines. Treatment of the disease has relied on pentavalent antimonials which have various drawbacks majorly the adverse side effects on users (Oryan and Akbari, 2016), high procurement cost (Gutierrez et al., 2016), long periods of administration (Clemente et al., 2018) and development of resistance by the parasites (Ponte-Sucre et al., 2017).
Though many experimental procedures have been investigated for Leishmania species, use of natural products remains stand-alone due to variety of chemical constituents present which work synergistically on multiple-target bringing about therapeutic effect (Bastos et al., 2016). This is collaborated by Mutoro et al. (2018b), in which the growth of Leishmania major parasites was inhibited by Plumbago capensis and Solanum nigrum extracts combined. Plants therefore may offer an alternative remedy as they are affordable, less toxic and easily available. The investigations of plants belonging to family of Leguminosae, Rosaceae, Aloaceae, Compositae, Myrtaceae, Barberidaceae, Boraginaceae, Lamiceae, Solaneceae, Juglandaceae, Asteraceae, Fabaceae etc have proven to be effective against Leishmaniasis (Da Silva et al., 2018; Bahmani et al., 2015; Serakta et al., 2013). Kinuthia et al. (2013) reported that Moringa stenopetala, Callistemon citrinus and Allium sativum plants are effective in elimination of L. major. More than twenty- six (26) Asteraceae plant families have demonstrated usefulness in allergies, burns; examples are Giardia lamblia and Leishmania species (Ivanescu et al., 2015).
The findings by Shamsi et al. (2018) implicated Matricaria chamomilla, Echinacea purpurea, Artemisia sieberi and Lavandula spica to be effective against Leishmaniasis. In Kenya, Warbugia ugandensis has been used by healers in management of Leishmaniasis (Ngure et al., 2009). These findings offer hope that natural products with antiparasitic activity could aid as templates for the manufacture of antileishmanial drugs. In the current work, the activity of B. pilosa, T. diversifolia and S. oleraceus; methanol, hexane, dichloromethane, ethyl acetate extracts and aqueous phase on L. major parasites was investigated. The plant has been used ethnobotanically to treat Leishmaniasis but has not been scientifically rationalized.
MATERIALS AND METHODS
Plant collection and extraction
Fresh aerial parts of S. oleraceus (Figure 1a), T. diversifolia (Figure 1b) and B. pilosa (Figure 1c) were collected from the Egerton University, Botanical garden (0°22'11.0"S, 35°55'58.0"E) with the help of a taxonomist for identification; they were assigned voucher number SK 214, SK 215 and SK 216 respectively . The botanical garden was chosen due to its richness in a wide variety of indigenous plant and its proximity to the laboratory where the analysis was performed. The aerial parts of S. oleraceus, T. diversifolia and B. pilosa were collected in September 2017 and processed as described by King’ondu et al. (2009).

Briefly, the plant parts were chopped into pieces, dried under a shade for 21 days and finally ground to powder in an electric mill. Approximately, 500 g of powdered plant was soaked in methanol overnight at room temperature. The filtrate obtained through Whatman No.1 filter paper was evaporated in rotary evaporator (BUCHI R-200) at 30-35°C under reduced pressure. The methanol extract was mixed with water and partitioned sequentially with hexane, dichloromethane and ethyl acetate to obtain their respective extracts after evaporation.
Phytochemical analysis
Phytochemical screening for alkaloids, glycosides, saponins, tannins, flavonoids, terpenoids and anthraquinones was done employing the methods of Santhi and Sengottuvel (2016) and Verma et al. (2018).
Preparation of test compounds
Stock solutions of methanol, hexane, dichloromethane, ethyl acetate and aqueous phase of B. pilosa, T. diversifolia and S. oleraceus were made in 5 ml Schneider’s Drosophila insect medium in readiness for assays. Each stock solution was filtered through 0.22 µm filter flasks in a laminar flow hood. Some plant extracts that did not dissolve in the medium were dissolved in 1% dimethyl Sulfoxide (DMSO) to prevent solvent carry-over. All the prepared plant extracts were stored at 4°C and retrieved later for use.
Cytotoxicity on Vero cells
The cytotoxicity assay on Vero cells was conducted by the 3-(4,5-Dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide (MTT) (Sigma®) protocol (Wabwoba et al., 2010). The quantity of insoluble formazan obtained from reduction of MTT reagent was proportional to number of viable cells. Vero cells were cultured and maintained in Minimum Essential Medium (MEM) (ATCC® 30-2003™) and supplemented with 10% Fetal bovine serum (FBS). Vero cells were harvested by trypsinization and pooled in 50 ml vials. A hundred microliters (100 µl) of the cell suspension was placed into two wells of rows A-H in a 96- flat-bottomed microtiter plate at a concentration of 1×106 cells per ml of the culture medium per well and incubated at 37 o C in 5% CO2 to facilitate plating. MEM was gently aspirated off and 200 µl of concentration (10 mg/ml) of the extracts was placed into two wells before dilutions of 2.0×103, 1.0×103, 5.0×102, 2.5×102, 1.25×102, and 6.25×101 were carried out. The tests were carried out in duplicate. The Vero cells and test extracts were then incubated at 37°C for 48 h in 5% CO2. Ten micro-litres (10 µl) of MTT was added to each well and further incubated for 3 h until a purple precipitate (formazan) was observed. The medium with MTT was aspirated off and the formazan solubilized with 100 μL of DMSO left to settle for 10 min. The absorbance reading for each well was measured using a micro-titer plate reader at wavelength of 562 nm. The number of viable cells was calculated at each concentration, according to Wang et al. (2006)’s formula: [(A/B) × 100], where A was the absorbance value for treated cells and B was the absorbance value for untreated cells. The extracts were considered cytotoxic if IC50 was less than 30 µg/ml (IC 50 ≤ 30 µg/ml) and non-cytotoxic if otherwise as stated by Sudha and Masilamani (2012).
Parasite preparation
The L. major strain (IDUB/KE/94=NLB-144) was sourced from Institute of Primate Research (IPR), Kenya. The parasites were grown to stationary phase at 25°C in Schneider’s Drosophila insect medium supplemented with 20% heat inactivated fetal bovine serum (FBS)-HYCLONE® USA), 100 µg/ml penicillin and 500 U/ml streptomycin (Hendricks and Wright, 1979), and 250 µg/ml 5-fluorocytosine arabinoside (Kimber et al., 1981). The stationary-phase meta-cyclic stage promastigote were obtained by centrifugation at 1500 × g and 4°C for 15 min. The meta-cyclic promastigote was used for analysis.
Anti-promastigote assays
The in vitro tests were done according to Mikus and Steverding (2000), using Alamar Blue bioassay. L. major meta-cyclic promastigote at the log phase (1×106 parasites per ml) of the culture medium was grown in a flat bottomed 24-well plate at 25°C for 48 h. Aliquots (50 µl) of the promastigotes were transferred into two wells of rows A-H in a 96-flat bottomed microtiter plate and further incubated for 24 h at 27°C. Two hundred microliters (200 µl) of (10 mg/ml) the plant extracts were added before serial dilutions of 10×103, 5×103, 2.5×102, 1.25×102, and 6.25×101 were done. The control wells contained L. major promastigotes in medium whereas the blank wells had the medium alone and positive control contained pentostam (Glaxo Operations (UK) Limited, Barnard Castle, UK) at concentration of 100 ug/ml. Each crude extract was tested in duplicate. After incubation for 24 h, the plates were removed and 10 µl of MTT was added into each well and further incubated for 4 h period. The medium and MTT were aspirated off, then 100 µl of Dimethyl sulfoxide (DMSO) was added and the plates shaken for 5 min. Optical densities readings were measured at 562 nm using micro titer reader. A decrease in fluorescence representing inhibition was deduced as percent of control culture fluorescence. Optical densities reading was used to generate 50% inhibitory concentration (IC50) for the different plants using the Alamar blue bioassay program.
Data analysis
The IC50 values were determined using Alamar blue program V2. The data presented for activity were analyzed using SAS system. All the statistical comparisons were done with Ryan –Einof-Gabriel-Wels multiple ranges tests and assigned at P≤ 0.05 level of significance.
RESULTS
Phytochemical tests
Phytochemical profiling of S. oleraceus (Figure 1a), T. diversifolia (Figure 1b) and B. pilosa (Figure 1c) extracts showed that alkaloids, saponins, flavonoids, glycosides, tannins, terpenoids and anthraquinones were present. The tests were based on color change and observation of precipitation and scored as + for presence and – for absence as shown in Table 1.
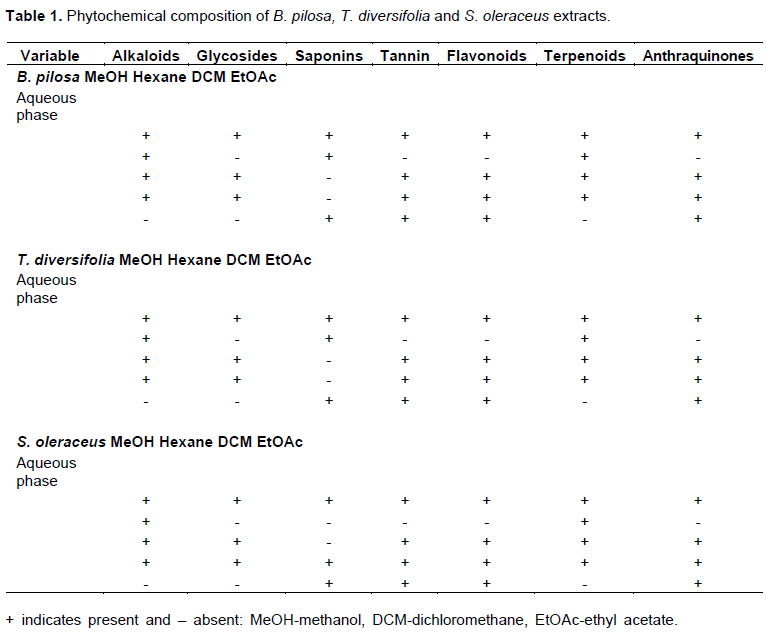
Antileishmanial activity of plants extracts
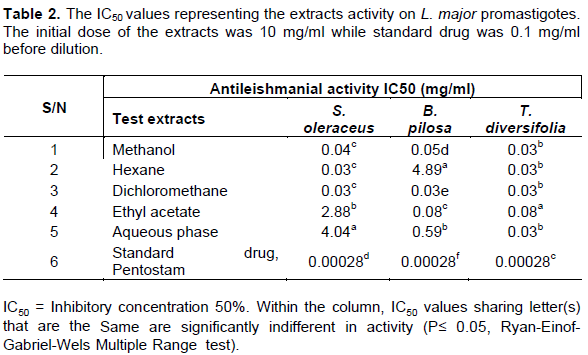
The extracts from S. oleraceus, B. pilosa, and T. diversifolia on Leishmania parasites displayed IC50 values from 0.03 to 4.89 mg/ml (Table 2). The extracts showed a concentration dependent activity which reciprocated to a high activity (a low IC50) of the extracts. S. oleraceus; methanol, hexane, dichloromethane extracts showed significant anti-leishmanial activity (0.04, 0.03, 0.03 and 0.03 mg/ml, respectively) in comparison to ethyl acetate and aqueous phase (2.88 and 4.04 mg/ml, respectively). B. pilosa; methanol, dichloromethane and ethyl acetate extracts had significant activity (0.05, 0.03 and 0.08 mg/ml, respectively) when compared to hexane extract and aqueous phase (0.59 and 4.89 mg/ml, respectively). Whereas, T. diversifolia methanol, hexane, dichloromethane, ethyl acetate extracts and aqueous phase have significant antileishmanbial activity (0.03, 0.03, 0.03, 0.08 and 0.03 mg/ml, respectively). Across plants extracts, dichloromethane registered the highest activity (0.03 mg/ml). The standard drug, pentostam at 0.1 mg/ml showed an IC50 value of 0.00028 mg /ml as shown in Table 2.
Cytotoxic effects of extracts
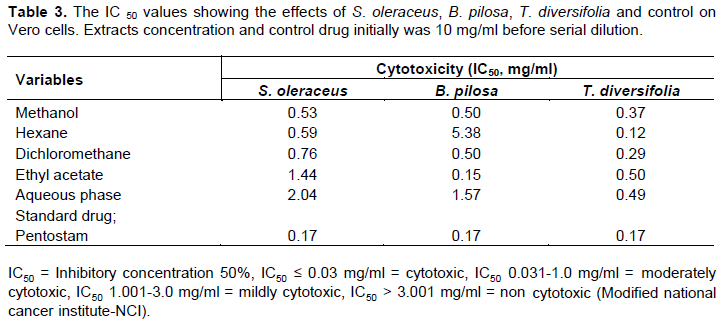
Extracts from the S. oleraceus, B. pilosa and T. diversifolia on Vero cells elicited IC50 from 0.12-5.38 mg/ml (Table 3). Criteria for scoring the cytotoxic effects on Vero cells was based on IC50 values from the National Cancer Institute (NCI) for extracts (Sudha and Masilamani, 2012): IC50 ≤ 0.03 mg/ml = cytotoxic, IC50 0.031-1.0 mg/ml = moderately cytotoxic, IC50 1.001-3.0 mg/ml = mildly cytotoxic, IC50 > 3.001 mg/ml = non cytotoxic. S. oleraceus: methanol, hexane and dichloromethane showed moderate toxicity (0.53, 0.59 and 0.76 mg/ml, respectively), while ethyl acetate and aqueous phase showed mild toxicity (1.44 and 2.04 mg/ml, respectively). B. pilosa; methanol, dichloromethane and ethyl acetate elicited moderate toxicity (0.50, 0.50 and 0.15 mg/ml respectively). While aqueous phase had mild toxicity (1.57 mg/ml) and hexane was non cytotoxic (5.38 mg/ml). Whereas T. diversifolia; methanol, hexane and dichloromethane, ethyl acetate and aqueous phase elicited moderate toxicity (0.37, 0.12, 0.29, 0.50 and 0.49 mg/ml respectively). The standard drug, pentostam exhibited IC50 values at 0.17 mg/ml (moderately toxic).
DISCUSSION
Phytochemical analysis
Qualitative phytochemical analysis of B. pilosa, S. oleraceus and T. diversifolia extracts revealed some differences in the constituents tested. The plants contained flavonoids, terpenoids, tannins, saponins, glycosides, alkaloids and anthraquinones. Presence of these phytonutrients could be linked to the activity displayed by the plants. The alkaloids present form the main constituents in herbs and used as remedy for Leishmaniasis (Ashok et al., 2015). For instance, harmalin from Peganum harmala has been shown to cure Leishmaniasis by intercalating DNA and interfering with aromatic amino acids metabolism (Di Giorgio et al., 2004; Chan-Bacab et al., 2001). According to Lage et al. (2015), flavonoids have shown significant in vitro activity against Leishmania parasites by altering the parasite ultrastructure and mitochondria functions. They cause depolarization of membrane potential as well as do not interfere with release of free radicals by macrophages. Moreover, flavonoids and terpenoids from Asteraceae have been reported to poses antileishmanial activity (Sujongya et al., 2018). Whereas the glycoside interferes with unusual type IB topoisomerase enzyme of L. donovani causing in vitro and in vivo death (Saha et al., 2013). Anthraquinones were present in the extracts and have shown selective activity on L. major parasites (Chan-Bacab et al., 2001). The chemical constituents present suggests the importance of these plants and therefore supports basis for ethnobotanical use as well as in chemical synthesis of drugs used in modern medicine.
Cytotoxic effects on Vero cells
The plants displayed mild to moderate cytotoxicity on Vero cells. Generally, an increase in concentration of the extracts resulted in more toxicity on L. major parasites thus inhibiting their growth. Although pentavalent antimonials are preferred line for Leishmaniasis treatment, they have major draw backs on users (Monzote, 2009) and the current study confirmed that pentostam exhibited moderate toxicity. B. pilosa aqueous phase elicited mild toxicity while methanol, dichloromethane and ethyl acetate extracts showed moderate cytotoxic effects with a low IC50 value for activity. The cytotoxic effects of the plant’s extracts could be responsible for antileishmanial property (Mans et al., 2016). The findings of Alvarez et al. (1996) demonstrated B. pilosa leaves to have significant carcinogenic action on colon cancer (HCT-15). This compliments our study in which the extracts showed moderate toxicity (Table 3). Whereas the hexane extract was non cytotoxic on Vero cells and studies by Shandukani et al. (2018) confirmed the plant to have no effect on cell lines after treatment exposure. T. diversifolia extracts had moderate cytotoxicity with high leishmanicidal activity (Low IC50 values). Studies by De Toledo et al. (2014) showed that presence of sesquiterpene lactones, major class of compounds present in T. diversifolia correlated with the cytotoxic effect on Vero cells. The plant’s cytotoxicity has been facilitated by addition reaction of thiol moieties present in the carbonyls of lactone structures (Guillermo et al., 2016; Ivanescu et al., 2015). Therefore, the cytotoxic compounds present could be responsible for in vitro antileishmanial activity while causing low Vero cell viability. Zhao et al. (2012) affirmed that most of the compounds derived from T. diversifolia were nontoxic to differentiated 3T3-L1 adipocytes at the concentration tested. S. oleraceus; methanol, hexane, dichloromethane had moderate toxicity while ethyl acetate extracts and aqueous phase were mildly toxic. Interestingly, among the five extracts of S. oleraceus, toxicity reduced as polarity of solvent increased from hexane to aqueous phase. Moreover, studies by Mc Dowell et al. (2011) of S. oleraceus extracts on HepG2 cells at 100 mg/ml showed to be non-toxic. This was indicator that S. oleraceus had more polar compounds therefore least cytotoxic effects on Vero cells which corresponded to low antileishmanial activity.
Anti-leishmanial activity
Majority of world population depend solely on herbs for their remedies against numerous infectious diseases (WHO, 2017). Therefore, the advent of synthetic drugs led to neglect of natural products as source of antimicrobials (Ezatpour et al., 2015). However, the management of Leishmaniasis using these drugs have posed challenges such as adverse side effects on users, cost, requirement for long periods of administration and parasite resistance. Moreover, there are no effective vaccines available to cure the disease (Al-Hajj et al., 2018). In this work, antileishmanial activities of B. pilosa, T. diversifolia and S. oleraceus; methanol, hexane, dichloromethane, ethyl acetate extracts and aqueous phase was investigated on L. major. Consequently, the results showed the plants had significant antileishmanial activity and inhibited the growth of L. major parasites. The flavonoids and phenolic compounds present in B. pilosa confer the plant with pharmacological activity (Cortés-Rojas et al., 2013). Interestingly, the plant has been used traditionally to cure illness as pain, diabetes, edema and inflammation (Lima et al., 2011). Other studies conducted have implicated B. pilosa, 1-phenyl-hepta-1,3,5-triyne to have antimalarial activity and cancer inhibitor (Kumari et al., 2009). The presence of flavonoids, abundant class of compounds in B. pilosa (Lima et al., 2011), promotes formation of cleavable complex as well as induces apoptosis through arrest of G1 phase (Mittra et al., 2000). This in turn inhibits growth of Leishmania parasites and subsequent low IC50 values.
According to Al-Hajj et al. (2018), flavonoids inhibit DNA topoisomerase I activity of L. donovani enzyme responsible for relegation step in parasite growth. In Leishmania donovani parasites, presence of flavonoids causes reversal of multi-drug resistance through binding to glycoprotein like transporter and inhibits synthesis of heat shock proteins responsible for evasion of host immunity (Hamarsheh et al., 2017). Therefore, a lower IC50 for activity, a better antileishmanial activity for B. pilosa; hexane, dichloromethane, ethyl acetate extracts and aqueous phase. According to Makwali et al. (2012), presence of saponins in the B. pilosa extracts complemented the activity of the flavonoids by acting on the parasite membrane inducing a drop in membrane potential. Our study was further complemented by Delmas et al. (2000), in which alpha- Hederin and other saponins isolated had antileishmanial activity. Therefore, in contrast to a single therapy, the various bioactive compounds present in the extract led to a synergy (Makwali et al., 2012) as it brought concept of structure-function approach in killing the parasites. Moreover, studies by Mutoro et al. (2018b) demonstrated that combinational approach in treatment reduces toxicity, lowers parasite resistance and optimizes drug efficacies.
T. diversifolia extracts showed activity against Leishmania parasites, in which dichloromethane showed strongest antileishmanial activity. The report herein was corroborated by De Toledo et al. (2014), in which dichloromethane extract from T. diversifolia had strong activity against Leishmania parasites. Notably, the dichloromethane extract becomes concentrated with large quantities of sesquiterpenes compounds from T. diversifolia glandular where they are manufactured and stored during successive partitioning (De Toledo et al., 2014). Additionally, T. diversifolia dichloromethane treated promastigotes lost distinct traits such as change in body morphology and loss of motility apparatus. Studies by Cortes-Selva et al. (2005), demonstrated the ability of sesquiterpenes to reverse multidrug resistance in Multi-Drug Resistance Leishmania tropica lines.
S. oleraceus has been used by different cultures to avert various diseases such as diarrhea, pneumonia and hepatitis (McDowell et al., 2011). Our study showed activity against Leishmania parasites thus validating the use of plant. Both dichloromethane and hexane extract had the highest activity. This could be attributed to polyphenols and flavonoids present (Table 1). Our study is in agreement with Jimoh et al. (2011) who showed that phenols were responsible for pharmacological activity.
On the other hand, activity decreased from ethyl acetate to aqueous phase, an indicator that bioactive compounds in S. oleraceus mainly remained in hexane and dichloromethane extracts, thus significant antileishmanial activity. The findings by Jimoh et al. (2011) revealed S. oleraceus methanol to have high flavonoid contents compared to aqueous phase. Moreover, the ethyl acetate extract registered a low activity. This could be as a result of low concentration of bioactive constituents. In summary, the activity of the various extracts on Leishmania parasites could be linked to the cell biology, where the chemical composition of the cell comprises ether lipids (LPGs, GPI-anchored metalloprotease, gp63, etc) which gives the parasite ability to evade host immunity and virulence (Weingartner, 2012). The availability of chemical constituents in the three plants leads to attachment of the corresponding groups in the parasite cell membrane. This attachment causes disruption of the Leishmania cell membrane and leakage of parasite contents leading to eventual death.
CONCLUSION
The findings of our study demonstrated that B. pilosa, T. diversifolia and S. oleraceus; methanol, hexane, dichloromethane, ethyl acetate extracts and aqueous phase had antileishmanial activity against L. major parasites. Therefore, with these promising results, the plants can be explored as alternative source of drugs for the management of Leishmaniasis as they are easily accessible and safe. Moreover, the results obtained provide basis for scientific rationalization of B. pilosa, T. diversifolia and S. oleraceus for use in pharmaceutical drug developments.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGMENTS
The authors are grateful to Professor S. Kariuki, a taxonomist (Department of Biological Science, Egerton University) for identification of plant species and Kenya Medical Research Institute (KEMRI). This work was funded by ASAFEM Project.
REFERENCES
|
Al-Hajj MM, Al-Shamahy HA, Alkhatib BY, Moharram BA (2018). In vitro antileishmanial activity against cutaneous Leishmania parasites and preliminary phytochemical analysis of four Yemen medicinal. Universal Journal of Pharmaceutical Research 3(4)1-7. |
|
|
Alvarez L, Marquina S, Villarreal ML, Alonso D, Aranda E, Delgado G (1996). Bioactive polyacetylenes from Bidens pilosa. Planta Medica 62(04):355-357. |
|
|
Ashok P, Lathiya H, Murugesan S (2015). Manzamine alkaloids as antileishmanial agents: a review. European Journal of Medicinal Chemistry 97:928-936. |
|
|
Bahmani M, Saki K, Ezatpour B, Shahsavari S, Eftekhari Z, Jelodari M, Rafieian-Kopaei M, Sepahvand R (2015). Leishmaniosis phytotherapy: Review of plants used in Iranian traditional medicine on leishmaniasis. Asian Pacific Journal of Tropical Biomedicine 5(9):695-701. |
|
|
Bastos CCC, de Ávila PHM, dos Santos Filho EX, de Ávila RI, Batista AC, Fonseca SG, Lima EM, Marreto RN, de Mendonca EF, Valadares MC (2016). Use of Bidens pilosa L. (Asteraceae) and Curcuma longa L.(Zingiberaceae) to treat intestinal mucositis in mice: toxico-pharmacological evaluations. Toxicology Reports 3:279-287. |
|
|
Burza S, Croft SL, Boelaert M (2018). Leishmaniasis. Seminar. Lancet. 392:951-970. |
|
|
Chan-Bacab MJ, Peña-Rodríguez LM (2001). Plant natural products with leishmanicidal activity. Natural Product Reports 18(6):674-688. |
|
|
Clemente WT, Mourão PHO, Aguado JM (2018). Current approaches to visceral Leishmaniasis treatment in solid organ transplant recipients. Expert Review Of Anti-Infective Therapy 16(5):391-397. . |
|
|
Cortés-Rojas DF, Chagas-Paula DA, Da Costa FB, Souza CR, Oliveira WP (2013). Bioactive compounds in Bidens pilosa L. populations: a key step in the standardization of phytopharmaceutical preparaions. Revista Brasileira de Farmacognosia 23(1):28-35. . |
|
|
Cortes-Selva F, Jimenez IA, Munoz-Martinez F, Campillo M, Bazzocchi IL, Pardo L, Ravelo AG, Castanys S, Gamarro F (2005). Dihidro-β-agarofuran sesquiterpenes: a new class of reversal agents of the multidrug resistance phenotype mediated by P-glycoprotein in the protozoan parasite Leishmania. Current Pharmaceutical Design 11(24): 3125-3139. . |
|
|
da Silva VP, Alves CCF, Miranda MLD, Bretanha LC, Balleste MP, Micke GA, Silveira EV, Martins CHG, Ambrosio MALV, de Souza Silva T, Tavares DC (2018). Chemical composition and in vitro leishmanicidal, antibacterial and cytotoxic activities of essential oils of the Myrtaceae family occurring in the Cerrado biome. Industrial Crops and Products 123:638-645. |
|
|
De Queiroz AC, Dias TDLMF, Da Matta CBB, Silva C, Agra LH, de Araújo-Júnior JX, Araújo GBD, Moura FDBP, Alexandre-Moreira MS (2014). Antileishmanial activity of medicinal plants used in endemic areas in northeastern Brazil. Evidence-Based Complementary and Alternative Medicine Volume 2014, Article ID 478290, 9pp. |
|
|
de Souza AA, dos Santos TV, Jennings YL, Ishikawa EA, da Rocha Barata I, Silva MD, Lima JA, Shaw J, Lainson R, Silveira FT (2016). Natural Leishmania (Viannia) spp. infections in phlebotomine sand flies (Diptera: Psychodidae) from the Brazilian Amazon region reveal new putative transmission cycles of American cutaneous leishmaniasis. Parasite 2016:23. |
|
|
De Toledo J, Ambrósio S, Borges C, Manfrim V, Cerri D, Cruz A, Da Costa F (2014). In vitro leishmanicidal activities of sesquiterpene lactones from Tithonia diversifolia against Leishmania braziliensis promastigotes and amastigotes. Molecules 19(5):6070-6079. |
|
|
Delmas F, Di Giorgio C, Elias R, Gasquet M, Azas N, Mshvildadze V, Dekanosidze G, Kemertelidze E, Timon-David P (2000). Antileishmanial activity of three saponins isolated from ivy, α-hederin, β-hederin and hederacolchiside A1, as compared to their action on mammalian cells cultured in vitro. Planta Medica 66(04):343-347. |
|
|
Di Giorgio C, Delmas F, Ollivier E, Elias R, Balansard G, Timon-David P (2004). In vitro activity of the β-carboline alkaloids harmane, harmine, and harmaline toward parasites of the species Leishmania infantum. Experimental Parasitology 106(3-4):67-74. |
|
|
Ezatpour B, Saedi Dezaki E, Mahmoudvand H, Azadpour M, Ezzatkhah F (2015). In vitro and in vivo antileishmanial effects of Pistacia khinjuk against Leishmania tropica and Leishmania major. Evidence-Based Complementary and Alternative Medicine Volume 2015, Article ID 149707, 6 p. |
|
|
Guillermo Federico Padilla-Gonzalez, Felipe Antunes dos Santos , Fernando Batista Da Costa (2016). Sesquiterpene Lactones: More Than Protective Plant Compounds With High Toxicity. Critical Reviews in Plant Sciences 35(1):18-37. |
|
|
Gutierrez V, Seabra AB, Reguera RM, Khandare J, Calderón M (2016). New approaches from nanomedicine for treating Leishmaniasis. Chemical Society Reviews 45(1):152-168. . |
|
|
Hamarsheh O, Azmi K, Amro A, Schultheis M, Abdeen Z, Firdessa, Sawalha K, Al-Rimawi F, Yaghmour R, Moll H (2017). Antileishmanial potential of crude plant extracts derived from medicinal plants in palestine. Annals of Clinical Cytology and Pathology 3(4):1065. |
|
|
Hendricks L, Wright N (1979). Diagnosis of cutaneous Leishmaniasis by in vitro cultivation of saline aspirates in Schneider's Drosophila medium. The American Journal of Tropical Medicine and Hygiene 28(6):962-964. |
|
|
Ivanescu B, Miron A, Corciova A (2015). Sesquiterpene lactones from Artemisia genus: biological activities and methods of analysis. Journal of analytical methods in Chemistry 2015:1-21. . |
|
|
Jimoh FO, Adedapo AA, Afolayan AJ (2011). Comparison of the nutritive value, antioxidant and antibacterial activities of Sonchus asper and Sonchus oleraceus. Records of Natural Products 5(1):29. |
|
|
Kimber C, Evans D, Robinson B, Peters W (1981). Control of yeast contamination with 5-fluorocytosine in the in vitro cultivation of Leishmania spp. Annals of Tropical Medicine and Parasitology 75(4):453-454. |
|
|
King'ondu CK, Sriskandakumar T, Opembe N, Chen CH, Morey A, Suib SL, (2009). Green decomposition of organic dyes using octahedral molecular sieve Manganese Oxide catalysts. |
|
|
Kinuthia GK, Anjili CO, Gikonyo NK, Kigondu EM, Ingonga JM, Kabiru EW (2013). In vitro and in vivo activities of blends of crude aqueous extracts from Allium sativum L, Callistemon citrinus (Curtis) Skeels and Moringa stenopetala (Baker F) Cufodontis against Leishmania major. International Journal of Medicinal and Aromatic Plants 3(2):234-246. |
|
|
Kumari P, Misra K, Sisodia BS, Faridi U, Srivastava S, Luqman S, Darokar MP, Negi AS, Gupta, MM, Singh SC, Kumar JK (2009). A promising anticancer and antimalarial component from the leaves of Bidens pilosa. Planta Medica 75(01): 59-61. |
|
|
Lage PS, Chávez-Fumagalli MA, Mesquita JT, Mata LM., Fernandes SO, Cardoso VN, Soto M, Tavares CA, Leite JP, Tempone AG, Coelho EA (2015). Antileishmanial activity and evaluation of the mechanism of action of strychnobiflavone flavonoid isolated from Strychnos pseudoquina against Leishmania infantum. Parasitology Research 114(12):4625-4635. |
|
|
Lima SF, Fischer DCH, Fechine Tavares J, Sobral Slva M, Filgueiras de Athayde-Filho P, Barbosa-Filho JM (2011). Compilation of secondary metabolites from Bidens pilosa L. Molecules 16(2):1070-1102. |
|
|
Makwali JA, Wanjala FM, Kaburi JC, Ingonga J, Byrum WW, Anjili CO (2012). Combination and monotherapy of Leishmania major infection in BALB/c mice using plant extracts and herbicides. Journal of Vector Borne Diseases 49(3):123. |
|
|
Mans DRA, Beerens T, Magali I, Soekhoe RC, Schoone GJ, Oedairadjsingh K, Hasrat JA, Van Den Bogaart E, Schallig HDFH (2016). In vitro evaluation of traditionally used Surinamese medicinal plants for their potential anti-leishmanial efficacy. Journal of Ethnopharmacology 180:70-77. |
|
|
McDowell A, Thompson S, Stark M, Ou ZQ, Gould KS (2011). Antioxidant activity of puha (Sonchus oleraceus L.) as assessed by the cellular antioxidant activity (CAA) assay. Phytotherapy Research 25(12):1876-1882. . |
|
|
Mikus J, Steverding D (2000). A simple colorimetric method to screen drug cytotoxicity against Leishmania using the dye Alamar Blue. Parasitology International 48:265-269. . |
|
|
Mittra B, Saha A, Chowdhury AR, Pal C, Mandal S, Mukhopadhyay S, Bandyopadhyay S, Majumder HK (2000). Luteolin, an abundant dietary component is a potent anti-leishmanial agent that acts by inducing topoisomerase II-mediated kinetoplast DNA cleavage leading to apoptosis. Molecular Medicine 6(6):527-541. |
|
|
Monzote L (2009). Current treatment of leishmaniasis: a review. The Open Antimicrobial Agents Journal 1(1). |
|
|
Mutoro CN, Kinyua JK, Kariuki DW, Ingonga JM, Anjili CO (2018a). In vitro study of the efficacy of Solanum nigrum against Leishmania major. F1000Research 7(1329):1-17. http://dx.doi.org/10.12688/f1000research.15826.1 |
|
|
Mutoro CN, Kinyua J, Kariuki D, Ingonga JM, Anjili CO (2018b). Efficacy of the combination of crude extracts of Solanum nigrum and Plumbago capensis on Leishmania major. F1000Research 7(1556):1-14. . |
|
|
Ngure PK, Tonui WK, Ingonga J, Mutai C, Kigondu E, Ng'ang'a ZZ, Rukunga G, Kimutai A (2009). In vitro antileishmanial activity of extracts of Warburgia ugandensis (Canellaceae), a Kenyan medicinal plant. Journal of Medicinal Plants Research 3(2):061-066. . |
|
|
Oryan A, Akbari M (2016). Worldwide risk factors in Leishmaniasis. Asian Pacific Journal of Tropical Medicine 9(10):925-932. |
|
|
Ponte-Sucre A, Gamarro F, Dujardin JC, Barrett MP, Lopez-Velez R, Garcia-Hernandez R, Pountain AW, Mwenechanya R, Papadopoulou B (2017). Drug resistance and treatment failure in Leishmaniasis: A 21st century challenge. PLoS Neglected Tropical Diseases 11(12):e0006052. |
|
|
Santhi K, Sengottuvel R (2016). Qualitative and quantitative phytochemical analysis of Moringa concanensis Nimmo. International Journal of Current Microbiology and Applied Sciences 5(1):633-640. |
|
|
Serakta M, Djerrou Z, Mansour-Djaalab H, Kahlouche-Riachi F, Hamimed S, Trifa W, Belkhiri A, Edikra N, Pacha YH (2013). Antileishmanial activity of some plants growing in Algeria: Juglans regia, Lawsonia inermis and Salvia officinalis. African Journal of Traditional, Complementary and Alternative Medicines 10(3):427-430. |
|
|
Shamsi M, Abbasi N, Mohajer A, Hoseini M, Rafieian-Kopaei M (2018). The most important native medicinal plants effective against cutaneous Leishmaniasis in mouse. International Journal of Life Science and Pharmaresearch 8(2):1-7. |
|
|
Shandukani PD, Tshidino SC, Masoko P, Moganedi KM (2018). Antibacterial activity and in situ efficacy of Bidens pilosa Linn and Dichrostachys cinerea Wight et Arn extracts against common diarrhoea-causing waterborne bacteria. BMC Complementary and Alternative Medicine 18(1):171. |
|
|
Saha S, Mukherjee T, Chowdhury S, Mishra A, Chowdhury SR, Jaisankar P, Mukhopadhyay S, Majumder HK (2013). The lignan glycosides lyoniside and saracoside poison the unusual type IB topoisomerase of Leishmania donovani and kill the parasite both in vitro and in vivo. Biochemical Pharmacology 86(12):1673-1687. |
|
|
Sudha S, Masilamani SM (2012). Characterization of cytotoxic compound from marine sediment derived actinomycete Streptomyces avidinii strain SU4. Asian Pacific Journal of Tropical Biomedicine 2(10):770-773. |
|
|
Verma A, Dhanik J, Agarwal D, Arya NNV (2018). Qualitative phytochemical and cluster analysis of genotypic extracts of coriander leaves and seeds from Tarai and Kumaun regions of Uttarakhand, Himalayan state of India. International Journal of Chemical Studies 6(2):1566-71. |
|
|
Verma A, Dhanik J, Agarwal D, Arya NNV (2018). Qualitative phytochemical and cluster analysis of genotypic extracts of coriander leaves and seeds from Tarai and Kumaun regions of Uttarakhand, Himalayan state of India. IJCS. 6(2):1566-71. |
|
|
Wabwoba BW, Anjili CO, Ngeiywa MM, Ngure PK, Kigondu EM, Ingonga J, Makwali J (2010). Experimental chemotherapy with Allium sativum (Liliaceae) methanolic extract in rodents infected with Leishmania major and Leishmania donovani. Journal of Vector Borne Disease 47(3):160. |
|
|
Wang X, Ge J, Wang K, Qian J, Zou Y (2006). Evaluation of MTT assay for measurement of emodin-induced cytotoxicity. Assay and Drug Development Technologies 4(2):203-207. |
|
|
Weingärtner A (2012). The plasma membrane lipid asymmetry of Leishmania donovani and its relevance for phagocytosis. |
|
|
World Health Organization (WHO) (2017). Unveiling the neglect of leishmaniasis. |
|
|
World Health Organization (WHO) (2018). Fact sheet: leishmaniasis. March 2018. |
|
|
Zhao G, Li X, Chen W, Xi Z, Sun L (2012). Three new sesquiterpenes from Tithonia diversifolia and their anti-hyperglycemic activity. Fitoterapia 83(8): 1590-1597. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


