Full Length Research Paper

ABSTRACT
Passiflora setacea D.C. is a wild species native to the Brazilian Cerrado, with agronomic and medicinal potentials. The cultivar BRS Pérola do Cerrado is the first registered and protected cultivar of a wild Passiflora and has become an alternative to the passion fruit market due to its desirable characteristics for food, ornamental and pharmaceutic industries. Since plant tissue culture techniques are considered important tools for large-scale production of plants and bioactive compounds, the goal of this work was to evaluate the flavonoid content and the antioxidant potential of hydroethanolic leaf extracts from in vivo and micropropagated plants of this new cultivar. Phytochemical analysis was performed by HPLC-UV-ESI-MS/TOF. Antioxidant activity was evaluated by the DPPH and the iron-chelating assays. The activities of antioxidant enzymes catalase, superoxide dismutase and ascorbate peroxidase (CAT, SOD and APX) were also determined. Two di-C-glucosyl flavonoids derivatives from apigenin were registered as major constituents in both leaf extracts analyzed and were identified as vicenin-2 and schaftoside. Despite the low antioxidant potential observed by the DPPH assay, leaf extracts from both in vivo and micropropagated plants showed high chelating capacity. Although no differences in the activity of SOD and CAT were observed, the specific activity of APX was increased in leaf extracts of micropropagated plants. These results suggest that in vitro plants can be used as an alternative for flavonoid production from cultivar BRS Pérola do Cerrado.
Key words: Flavonoid content, phytochemical analysis, antioxidant potential, leaf extracts, Passiflora setacea.
INTRODUCTION
The genus Passiflora comprises approximately 525 species, grouped into five subgenera that are found in tropical and subtropical regions. It is considered the most economically important genus of the Passifloracea family, since some species are consumed in natura or in the food industry. It also has a great ornamental potential due to the diversity of colors and shapes of its flowers (Faleiro et al., 2019). Moreover, some species are well known in folk medicine worldwide because of their pharmacological activities as sedatives, anxiolytics, analgesics, and anti-inflammatories. Several compounds, such as flavonoids, alkaloids and saponins have been identified in their fruits, leaves, flowers, stems and roots (Smruthi et al., 2021).
Passiflora setacea D.C. is a wild species native to the Brazilian Cerrado, with edible fruits, resistant to Fusarium oxysporum, Fusarium solani, Phytophthora, and to the Passion fruit woodiness virus (PWV) (Braga et al., 2006; Pereira et al., 2019). It is also used in folk medicine in the treatment of insomnia (Carvalho et al., 2018). The cultivar BRS Pérola do Cerrado, launched in 2013 by Embrapa (Brazilian Agricultural Research Corporation), is the first registered (RNC Nº 21714) and protected cultivar of a wild Passiflora species (SNPC Certificate Nº 20120197) (Viana et al., 2016). Its fruits are rich in mineral salts as well as phenolic compounds and proanthocyanidins, which are associated to antioxidant activity (Carvalho et al., 2018). Hence, it has become an alternative to the passion fruit market due to its desirable characteristics for food, ornamental and pharmaceutic industries (Faleiro et al., 2018).
Plant tissue culture techniques are considered important tools for large-scale production of bioactive compounds, since in vitro cultures allow high plant multiplication rates and the modulation of both morphogenic and biosynthetic capacities by modifying physical and chemical parameters (Chandran et al., 2020). Several authors have reported in vitro production of bioactive substances in Passiflora species, including flavonoids, in micropropagated plants of Passiflora caerulea L., Passiflora incarnata L. (Ozarowski and Thiem, 2013) and Passiflora foetida L. and Passiflora suberosa L. (Simão et al., 2018).
The stressful in vitro conditions may induce the accumulation of reactive oxygen species (ROS), leading to lipid peroxidation, protein oxidation and even cell death (Pashkovskiy et al., 2018). Although ROS formation is considered a normal physiological process and an important cell signaling element, their overproduction may exceed the antioxidant capacity of the cells, resulting in oxidative stress (Kim et al., 2017). In order to maintain basal levels of ROS and protect against oxidative stress, plant cells exhibit both non-enzymatic and enzymatic defenses (Yu et al., 2017).
Non-enzymatic antioxidant systems include flavonoids, which are commonly found in leaves and fruits of Passiflora spp. and have been used as chemical markers in the genus, due to their structural diversity and chemical stability (Gosmann et al., 2011). Most of them are classified as C-glucolsyl flavonoids derived from apigenin and luteolin, and frequently found as isomers (Ozarowski et al., 2018). Some authors have already reported the presence of the flavonoids orientin, isoorientin, vitexin, vitexin-2´´-O-rhaminoside and isovitexin, as well as acid ascorbic, polyamines, terpenes and carotenoids, in leaf and fruit pulp extracts from P. setacea and in its cultivar BRS Pérola do Cerrado (Santana et al., 2015; Bomtempo et al., 2016; Gomes et al., 2017; Wosch et al., 2017; Carvalho et al., 2018; Sanchez et al., 2020; da Rosa et al., 2021).
The objective of this work was to evaluate the flavonoid content and the antioxidant potential of hydroethanolic leaf extracts from in vivo and micropropagated plants of P. setacea cv BRS Pérola do Cerrado.
MATERIALS AND METHODS
Plant material and culture conditions
Seeds of P. setacea cv BRS Pérola do Cerrado were gently provided by Embrapa Cerrados, Brazil. Seeds were transferred to pots containing Plantmax® substrate for germination, and in vivo-grown plants (Figure 1a) were maintained in a greenhouse for twelve months. Micropropagated plants (Figure 1b), derived from in vitro seed germination, were obtained as described by Santos-Tierno et al. (2021) and maintained by bimonthly subcultures of stem segments (3 cm) on solidified half-strength MSM medium (MSM ½) (Monteiro et al., 2000). Plants were incubated in a growth chamber at 25±2°C under a 16 h photoperiod, using a total irradiance of 46 µmol m-2 s-1 provided by cool-white fluorescent lamps (Philips F40 CW).

Extract preparation
Leaves from in vivo and micropropagated plants were lyophilized for five days, then powdered and weighed. In order to determine the best flavonoid extraction procedure, the extracts from in vivo-grown plants were initially prepared using two methodologies: i) 40% ethanol (Tedia®, Brazil) under reflux for 1 h (Birk et al., 2005), or (ii) maceration in 90% ethanol with ultrasound-assisted extraction for 15 min. After that, extracts from leaves of in vivo and micropropagated plants were prepared following the selected extraction procedure, using 1:50 plant:solvent (w/v). The solvents were then evaporated in a rotary evaporator (Marconi – M120) at 40°C, before solubilization in methanol (Tedia®, Brazil). The yield of the extracts was calculated as follows:
[extract (g)/dried sample (g)] × 100.
Qualitative HPLC-DAD-UV analysis High-Performance
High-Performance Liquid Chromatography coupled to a Diode-Array Detector (HPLC-DAD-UV) was carried out using the Shimadzu Liquid Chromatograph system according to Costa et al. (2011), with modifications. The analyses were performed using a Thermo-Scientific© Hypersil Gold RP18 column (250 mm × 4.6 mm i.d. × 5 Å particle size), at a flow rate of 1.0 mL/min and oven temperature at 25°C. Leaf extracts from in vivo plants were solubilized in methanol at a final concentration of 1.0 mg/mL and the injected volume was 10 μL. "The mobile phase consisted of solvents A (MilliQ® water acidified with 1% glacial acetic acid, pH adjusted to 3.0) and B (acetonitrile)," with the following gradient elution: 95% of A and 5% of B (0 - 2 min); 95 - 80% of A and 5 -20% of B (2 - 30 min); 80% of A and 20% of B (30 - 40 min). All solvents were HPLC grade (Tedia®, Brazil). The UV absorption was monitored at 340 nm.
HPLC-UV-ESI-MS/TOF analysis
Extracts of leaves from in vivo and micropropagated plants were also analyzed by High-Performance Liquid Chromatography coupled to UV detector and coupled to Time-of-Flight Mass Spectrometer Detector (MicrOTOF II Mass Spectrometer, Bruker Daltonics, MA, USA) equipped with electrospray ionization (200°C - 9 μL/min - 4 psi) (HPLC-UV-ESI-MS/TOF). The analysis was performed using the same column and conditions described for HPLC-DAD-UV. Electrospray ion source (ESI)-MS spectra were acquired in both positive and negative ion modes and recorded in the range of m/z 50 to 1000, using nitrogen as the nebulizing gas (400 L/h), at 250°C. The ionization energy applied was 10 eV for both positive and negative modes. UV spectral data were recorded at 340 nm. Compounds identification was carried out by comparing the exact experimental mass of the pseudomolecular ion [M-H]- with those of the MassBank (High Quality Mass Bank Database, available at https://massbank.eu/MassBank/), as well as UV spectrum, elution order and considering flavonoids previously identified in Passiflora genus.
Determination of antioxidant activity
The antioxidant potential of leaf extracts was determined by the 2,2-diphenyl-1-picrylhydrazyl (DPPH) radical scavenging capacity assay (Sánchez-Moreno et al., 1998), and the iron chelating assay was carried out according to Chew et al. (2009).
The extract concentrations (g/L) required for quenching 50% of the initial DPPH radicals or quelating 50% of the Fe2+ (EC50), were determined graphically. The same procedure was carried out with quercetin (0.01 to 0.15 g/L) and ethylenediamine tetraacetic acid (EDTA) (0.001 to 0.03 g/L) solutions that were used as standard.
Evaluation of antioxidant enzymes activity
Protein extracts were prepared from fresh leaves (300 mg), according to Azevedo et al. (1998). The activities of the antioxidant enzymes superoxide dismutase (SOD), catalase (CAT) and ascorbate peroxidase (APX) were determined according to Vianna et al. (2019).
Statistical analysis
The assays for determining the antioxidant potential and activities of antioxidant enzymes were carried out in triplicates, in two independent experiments. Statistical evaluation of experimental data was performed by analysis of variance (ANOVA), followed by the post-test Tukey-Kramer for comparing all pairs of columns (0.05% significance level), using GraphPad Instat (GraphPad Software Inc., San Diego, CA).
RESULTS AND DISCUSSION
In this work, in order to determine the best flavonoid extraction procedure for P. setacea cv BRS Pérola do Cerrado, two methods were evaluated using leaves from in vivo-grown plants. Preliminary HPLC-DAD-UV analysis revealed more peaks when the extract was prepared with 90% ethanol using the ultrasound-assisted method in comparison to the extract prepared with 40% ethanol under reflux (Figure 2). Four compounds, with retention times ranging from 43 to 58 min and typical UV-vis spectra of carotenoids, were exclusively detected in the extracts prepared with the ultrasound-assisted method. This result could be associated with the higher levels of ethanol used by the extraction process, in addition to the propagation of ultrasonic waves, which disorganize the cell wall and cause leaking of cellular contents (Gunathilake et al., 2017; Castañeda-Valbuena et al., 2021).
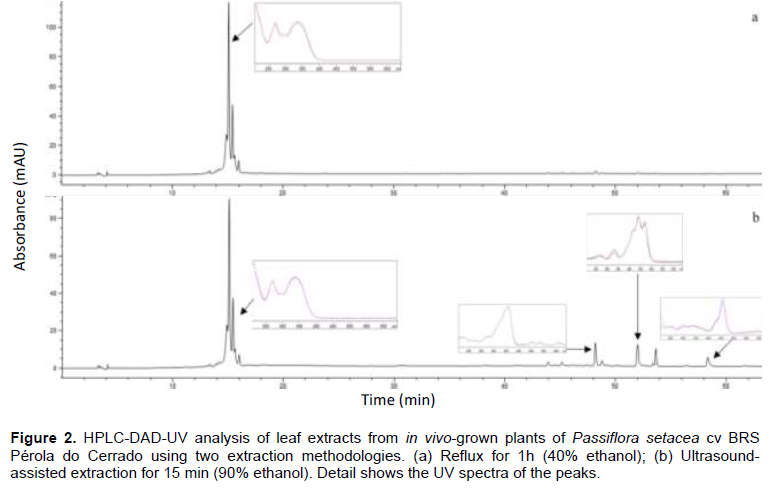
Considering the extraction efficiency and the shorter maceration period required (15 min), the ultrasound-assisted method was selected for the subsequent analyses. The yields of the leaf extracts from both in vivo and micropropagated plants obtained by the ultrasound-assisted method were 14.80 and 13.38%, respectively.
Compounds identification in leaf extracts from both in vivo and micropropagated plants performed by HPLC-UV-ESI-MS/TOF analyses revealed similar chromatographic profiles. The exact masses of the pseudomolecular ions [M-H]- were compared with literature data as described in the experimental and afforded compounds 1 to 8 (Table 1 and Figure 3). Sucrose and digalacturonic acid, commonly found in plant extracts and in the culture medium, were identified in the beginning of the chromatographic run. Two major constituents were registered in both leaf extracts: compound 6 at Rt 13.1 min (vicenin-2; [M-H]- = 593.1951; C27H29O15) and compound 7 at Rt 13.4 min (schaftoside; [M-H]- = 563.1824; C26H27O14). Other apigenin-C-glucosyl-derivatives were detected in minor amounts. Apigenin-6-C-glucoside or isovitexin (Rt = 13.9 min, M-H- = 431.1289; C21H19O10) was detected in both leaf extracts. On the other hand, apigenin-6-C-glucoside-7-O-glucoside (Rt = 11.5 min; [M-H]- = 593.1958; C27H29O15) and apigenin-8-C-glucoside-7-O-glucoside (Rt = 12.8 min; [M-H]- = 593.1943; C27H29O15) were only detected in leaves from micropropagated plants. The modulation of bioactive compounds production in in vitro systems was also observed for Poliomintha glabrescens (García-Pérez et al., 2011), Salvia dolomitica (Bassolino et al., 2015), Agave salmiana (Puente-Garza et al., 2017) and Kaempferia parviflora (Park et al., 2021).
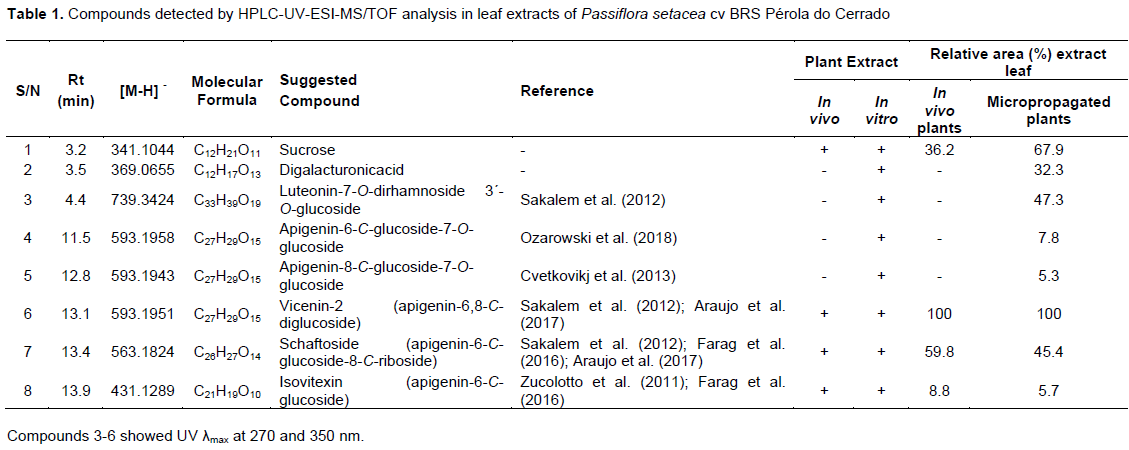
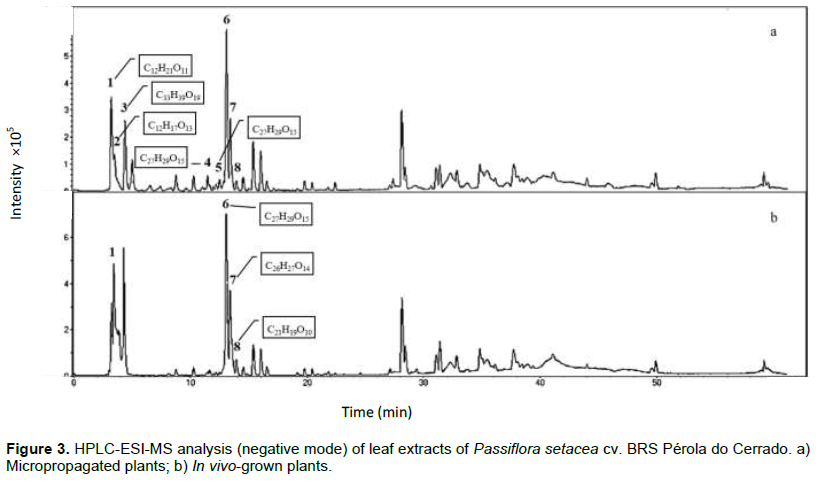
The two major constituents from both leaf extracts studied here, vicenin-2 and schaftoside, have already been identified in Passiflora spp. (Araújo et al., 2017; Farag et al., 2016; Sakalem et al., 2012; Zucolloto et al., 2011). Vicenin-2 has been detected in extracts from pericarp of Passiflora edulis (Sena et al., 2009; Zucolotto et al., 2009) and extracts from leaves and fruit pulp of Passiflora tripartite (Zucolotto et al., 2011; Simirgiotis et al., 2013). Schaftoside has been reported as the major constituent of P. incarnata (Abourashed et al., 2002) and in leaf extracts of Passiflora mucronata (Da Silva et al., 2018). Both flavonoids have been associated with sedative, anxiolytic and analgesic activities (Sakalem et al., 2012). In addition, vicenin-2 has been related to antiplatelet and anticoagulant activities (Lee and Bae, 2015), whereas schaftoside has been recently reported to be effective against the coronavirus SARS-CoV-2 (Yalç?n et al., 2021).
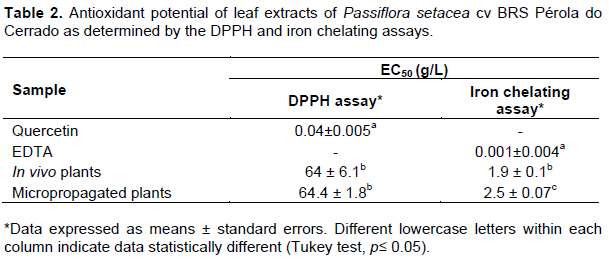
The antioxidant potential of extracts from leaves of in vivo and micropropagated plants of P. setacea cv BRS Pérola do Cerrado was also evaluated using two distinct techniques, the DPPH and iron chelation assays. Although several reports have studied the antioxidant potential of Passiflora species by the DPPH assay (Silva et al., 2013), the use of different and complementary techniques must be taken into consideration since plant-derived compounds may show distinct mechanisms of action (Santos-Sanchéz et al., 2019).
The DPPH radical scavenging capacity assay revealed a low antioxidant potential, with EC50 values of 64 ± 6.1 and 64.4 ± 1.8 g/L in extracts from in vivo and micropropagated plants, respectively (Table 2). On the other hand, these extracts showed high iron chelating capacity, with significantly low EC50 values. Higher chelating capacity was observed in leaves from in vivo-grown plants when compared with micropropagated plants (EC50 values 1.9 ± 0.1 and 2.5 ± 0.1 g/L, respectively) (Table 2). Since flavonoids with 5-hydroxy-4-keto-2,3-double bond are able to chelate iron ions (Mlad?nka et al., 2011), it is possible that this difference can be explained by the action of the apigenin type flavonoids that were found in this study (Table 1). Similar results were observed in root and rhizome extracts of Nardostachys chinensis and Valeriana officinalis (Wang et al., 2010a), root extracts of Periploca sepium (Wang et al., 2010b) and buds extracts of Brassica oleracea L. (Köksal and Gülçin, 2008).
The activity of enzymes associated with the cellular antioxidant system, namely SOD, CAT and APX, was also evaluated in the leaf extracts. SOD catalyzes the dismutation of the superoxide radical into molecular oxygen and hydrogen peroxide and thus is considered an important defense against free radicals, whereas CAT and APX are responsible for the degradation of hydrogen peroxide mainly in peroxisomes and chloroplasts, respectively (Kim et al., 2017). In this study, although no statistical differences were observed in the specific activity of SOD and CAT extracted from both leaf tissues (Figure 4a and b), there was a significant increase in APX activity in leaves from micropropagated plants (Figure 4c). These results might reflect the stressful conditions imposed by the in vitro environment, which led to an increase of hydrogen peroxide concentration and, thus, of the APX activity aiming its degradation. Moreover, considering that APX is the only enzyme responsible for maintaining low hydrogen peroxide levels in the chloroplasts, it is possible that its high activity rates, when compared with CAT activity, is associated with specific damages to the photosynthetic apparatus induced by in vitro conditions (Škodová-Sveráková et al., 2020).
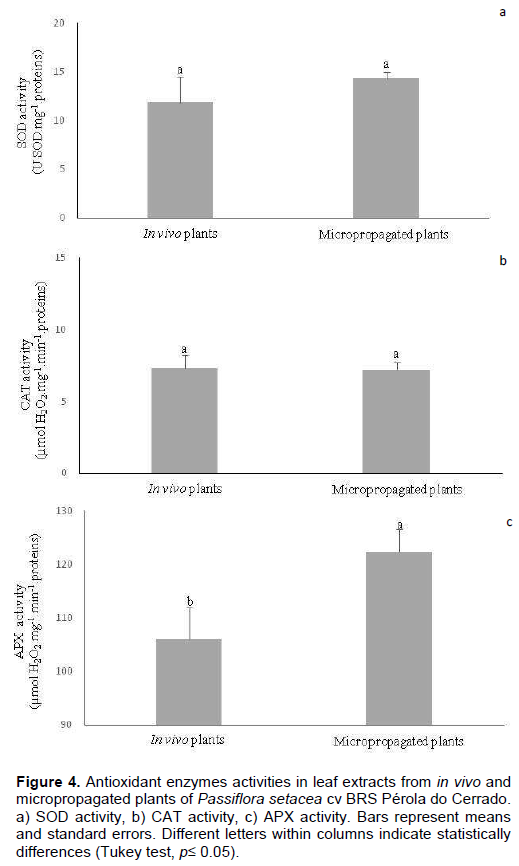
Hydrogen peroxide is not considered a highly reactive ROS. In high concentrations, however, it can interact with metallic ions, such as Fe+3, giving rise to hydroxyl radicals (OH•), which are considered one of the most relevant ROS, due to their potential to cause cell membrane damages and the absence of specific enzymatic mechanisms for their degradation (Kaczmarczyk et al., 2012; Sharma et al., 2012; Demidchik, 2015). Therefore, considering the increase in APX activity and the high iron chelating capacity observed here, it seems that both enzymatic and non-enzymatic antioxidant systems are acting together in order to reduce OH• formation (Hasanuzzaman et al., 2020).
In conclusion, the chromatographic analyses of leafextracts from both in vivo and micropropagated plants of P. setacea cv BRS Pérola do Cerrado described here detected the presence of vicenin-2 and schaftoside in higher content in comparison to the other flavonoids.
The antioxidant potential of the extracts was also evaluated, and despite the low capacity observed by the DPPH assay, a high antioxidant activity was accessed by the iron chelating assay, corroborating the importance of distinct and complementary assays in order to access the antioxidant potential of plant-derived compounds. These results suggest that in vitro plants can be used as an alternative for flavonoid production from cultivar BRS Pérola do Cerrado.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGMENTS
This work was supported by Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ) [Grant Number E-26/010.001840/2019], and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) [Grant Number 310238/2018-8]. This study was also funded by Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES) [Finance Code 001].
REFERENCES
|
Abourashed E, Vanderplank J, Khan I (2002). High-Speed extraction and HPLC fingerprinting of medicinal plants -I. Application to Passiflora flavonoids. Pharmaceutical Biology 40(2):81-91. |
|
|
Araújo MH, Silva ICV, Oliveira PF, Barreto ARR, Konno TUP, Esteves FA, Barth T, Aguiar FA, Lopes PL, Dermenjian RK, Guimarães DO, Leal ICR, Lasunskaia EB, Muzitano MF (2017). Biological activities and phytochemical profile of Passiflora mucronata from the Brazilian resting. Brazilian Journal of Pharmacognosy 27:702-710. |
|
|
Azevedo RA, Alas RM, Smith RJ, Lea PJ (1998). Response of antioxidant enzymes to transfer from elevated carbon dioxide to air and ozone fumigation, in the leaves and roots of wild-type and a catalase-deficient mutant of barley. Physiologia Plantarum 104(2):280-292. |
|
|
Bassolino L, Giacomelli E, Giovanelli S, Pistelli L, Cassetti A, Damonte G, Ruffoni B (2015) Tissue culture and aromatic profile in Salvia dolomitica Codd. Plant Cell, Tissue and Organ Culture 121(1):83-95. |
|
|
Birk CD, Provensi G, Gosmann G, Reginatto FH, Schenkel EP (2005). TLC fingerprint of flavonoids and saponins from Passiflora species. Journal of Liquid Chromatography Related Technologies 28(14):2285-2291. |
|
|
Bomtempo LL, Costa AM, Lima H, Engeseth N, Gloria MBA (2016) Bioactive amines in Passiflora are affected by species and fruit development. Food Research International 89(1):733-738. |
|
|
Braga MF, Santos EC, Junqueira NTV, Sousa AATC, Faleiro FG, Rezende LN, Junqueira KP (2006). Cutting rooting of three wild Passiflora species. Revista Brasileira de Fruticultura 28(2):284-288. |
|
|
Carvalho MVO, Oliveira LL, Costa AM (2018). Effect of training system and climate conditions on phytochemicals of Passiflora setacea, a wild Passiflora from Brazilian savannah. Food Chemistry 266:350-358. |
|
|
Castañeda-Valbuena D, Ayora-Talavera T, Luján-Hidalgo C, Álvarez-Gutiérrez P, Martínez-Galero N, Meza-Gordillo R (2021) Ultrasound extraction conditions effect on antioxidant capacity of mango by-product extracts. Food and Bioproducts Processing 127:212-224. |
|
|
Chandran H, Meena M, Barupal T, Sharma K (2020). Plant tissue culture as a perpetual source for production of industrially important bioactive compounds. Biotechnology Reports 26:e00450. |
|
|
Chew YL, Goh JK, Lim YY (2009). Assessment of in vitro antioxidant capacity and polyphenolic composition of selected medicinal herbs from Leguminosae family in Peninsular Malaysia. Food Chemistry 116(1):13-18. |
|
|
Costa GM, Ortmann CF, Schenkel EP, Reginatto FH (2011) Na HPLC-DAD method to quantification of main phenolic compounds from leaves of Cecropia species. Journal of the Brazilian Chemical Society 22(6):1096-1102. |
|
|
Cvetkovikj I, Stefkov G, Acevska J, Stanoeva JP, Karapandzova M, Stefova M, Dimitrovska A, Kulevanova S (2013). Polyphenolic characterization and chromatographic methods for fast assessment of culinary Salvia species from South East Europe. Journal of Chromatography A 1282:38-45. |
|
|
Da Rosa LC, Siqueira MRP, Paumgartten FJR, Pacheco G, Mansur E, Moreira DL (2021). Development and validation of a new method to quantify vitexin-2''-O-rhamnoside on Passiflora L. extracts. Journal of Medicinal Plants Research 15(1):45-55. |
|
|
Da Silva ICV, Kalu?erovi? GN, De Oliveira PF, Guimarães DO, Quaresma CH, Porzel A, Muzitano MF, Wessjohann LA, Leal ICR (2018). Apoptosis caused by triterpenes and phytosterols and antioxidant activity of an enriched flavonoid extract from Passiflora mucronata. Anti-Cancer Agents in Medicinal Chemistry 18(10):1405-1416. |
|
|
Demidchik V (2015). Mechanisms of oxidative stress in plants: From classical chemistry to cell biology. Environmental and Experimental Botany 109:212-228. |
|
|
Faleiro FG, Junqueira NTV, Jesus ON, Costa AM (2018). Avanços e perspectivas do melhoramento genético de Passifloras no Brasil. In Congreso Latino-americano de Pasifloras, 2º, Neiva, Huila, Colômbia pp. 12-23. |
|
|
Morera MP, Costa AM, Faleiro FG, Carlosama AR, Carranza C (Eds.) Maracujá: dos recursos genéticos ao desenvolvimento tecnológico. Brasília, DF: ProImpress pp.81-93. |
|
|
Faleiro FG, Junqueira NTV, Junghans TG, Jesus ON, Miranda D, Otoni WC (2019). Advances in passionfruit (Passiflora spp.) propagation. Revista Brasileira de Fruticultura 41(2):e-155. |
|
|
Farag MA, Otify A, Porzel A, Michel CG, Elsayed A, Wessjohann LA (2016). Comparative metabolite profiling and fingerprinting of genus Passiflora leaves using a multiplex approach of UPLC-MS and NMR analyzed by chemometric tools. Analytical Bioanalytical Chemistry 408(12):3125-3143. |
|
|
García-Pérez E, Gutiérrez-Uribe JA, García-Lara S (2011). Luteolin contente and antioxidant activity in micropropagated plants of Polimintha glabrescens (Gray). Plant Cell, Tissue and Organ Culture 108(3):521-527. |
|
|
Gomes S, Portugal L, Anjos J, Jesus O, Oliveira E, David J, David J (2017). Accelerated solvent extraction of phenolic compounds exploiting a Box-Behnken design and quantification of five flavonoids by HPLC-DAD in Passiflora species. Microchemical Journal 132:28-35. |
|
|
Gosmann G, Provensi G, Comunello LN, Rates SMK (2011). Composição química e aspectos farmacológicos de espécies de Passiflora L. (Passifloraceae). Brazilian Journal of Biosciences 9(1):88-99. |
|
|
Gunathilake KDP, Ranaweera KKDS, Rupasinghe HPV (2017). Optimization of phenolics and carotenoids extraction from leaves of Passiflora edulis using response surface methods. Asian Journal of Biotechnology and Bioresource Technology 1(1):1-12. |
|
|
Hasanuzzaman M, Bhuyan MHM, Zulfiqar F, Raza A, Mohsin SM, Mahmud JA, Fujita M, Fotopoulos V (2020). Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 9(8) 681. |
|
|
Kaczmarczyk A, Funnekotter B, Menon A, Phang PY, Al-Hanbali A, Bunn E, Mancera RL (2012). Current Issues in Plant Cryopreservation. In. Katkov I (Ed.), Current Frontiers in Cryobiology. London: IntechOpen pp. 417-438. |
|
|
Kim Y-H, Khan AL, Waqas M, Lee IJ (2017). Silicon regulates antioxidant activities of crop plants under abiotic-induced oxidative stress: a review. Frontiers in Plant Science 8:510. |
|
|
Köksal E, Gülçin I (2008). Antioxidant activity of cauliflower (Brassica oleracea L.). Turkish Journal of Agriculture and Forestry 32(1):65-78. |
|
|
Lee W, Bae JS (2015). Antithrombotic and antiplatelet activities of vicenin-2. Blood Coagulation and Fibrinolysis 26(6):628-34. |
|
|
Mlad?nka P, Macáková K, Filipský T, Zatloukalová L, Jahodá? L, Bovicelli P, Silvestri IP, Hrdina R, Saso L (2011). In vitro analysis of iron chelating activity of flavonoids. Journal of Inorganic Biochemistry 105(5):693-701. |
|
|
Monteiro ABC, Higashi EN, Gonçalves NA, Rodriguez APM (2000). A novel approach for the definition of the inorganic medium components for micropropagation of yellow passion fruit (Passiflora edulis Sims. f. flavicarpa Deg.). In Vitro Cellular and Development-Plant 36(6):527-531. |
|
|
Ozarowski M, Piasecka A, Paszel-Jaworska A, Siqueira Chaves D, Romaniuk A, Rybczynska M, Gryszczy?ska A, Sawikowska A, Kachlicki P, Mikolajczak P, Seremak-Mrozikiewicz A, Klejewski A, Thiem B (2018). Comparison of bioactive compounds content in leaf extracts of Passiflora incarnata, P. caerulea and P. alata and in vitro cytotoxic potential on leukemia cell lines. Brazilian Journal of Pharmacognosy 28:179-191. |
|
|
Ozarowski M, Thiem B (2013). Progress in micropropagation of Passiflora spp. to produce medicinal plants: a mini-review. Brazilian Journal of Pharmacognosy 23(6):937-947. |
|
|
Park H-Y, Kim K-S, Ak G, Zengin G, Cziáky Z, Jek? J, Adaikalam K, Song K, Kim D-H, Sivanesan I (2021). Establishment of a rapid micropropagation system for Kaempferia parviflora Wall. Ex Baker: phytochemical analysis of leaf extracts and evaluation of biological activities Plants 10(4):698. |
|
|
Pashkovskiy PP, Soshinkova TN, Korolkova DV, Kartashov AV, Zlobin IE, Lyubimov VY, Kreslavski VD, Kuznetsov VV (2018). The effect of light quality on the pro-/antioxidant balance, activity of photosystem II, and expression of light-dependent genes in Eutrema salsugineum callus cells. Photosynthesis Research 136(2):199-214. |
|
|
Pereira PPA, Lima LKS, Soares TL, Laranjeira FF, Jesus ON, Girardi EA (2019). Initial vegetative growth and survival analysis for the assessment of Fuisarium wilt resistance in Passiflora spp. Crop Protection 121:195-203. |
|
|
Puente-Garza CA, García-Lara S, Gutiérrez-Uribe JA (2017). Enhancement of saponins and flavonols by micropropagation of Agave salmiana. Industrial Crops and Products 105:225-230. |
|
|
Sakalem ME, Negri G, Tobach R (2012). Chemical composition of hydroethanolic extracts from five species of the Passiflora genus. Brazilian Journal of Pharmacognosy 22(6):1219-1232. |
|
|
Sanchez BAO, Celestino SMC, Gloria MBA, Celestino IC, Lozada MIO, Junior DAS, Alencar ER, Oliveira LL (2020). Pasteurization of passion fruit Passiflora setacea pulp to optimize bioactive compounds retention. Food Chemistry X:6-100084. |
|
|
Sánchez-Moreno C, Larrauri JA, Saura-Calixto FA (1998). Procedure to measure the antiradical efficiency of polyphenols. Journal of the Science of Food Agriculture 76(2):270-276. |
|
|
Santana FC, Shinagawa FB, Araujo ES, Costa AM, Mancini-Filho J (2015). Chemical composition and antioxidant capacity of Brazilian Passiflora seed oils. Journal of Food Science 80(12):C2647-C2654. |
|
|
Santos-Sanchéz NF, Salas-Coronado R, Villanueva-Cañongo C, Hernández-Carlos B (2019). Antioxidant compounds and their antioxidant mechanism. London, UK: IntechOpen pp. 23-50. |
|
|
Santos-Tierno R, Garcia R, Fonseca E, Faleiro F, Moreira D, Pacheco G, Mansur E (2021). Light quality and explant type modulate growth, antioxidant properties and bioactive compounds production of calluses of Passiflora setacea cv BRS Pérola do Cerrado. Plant Cell, Tissue Organ Culture 147(3):635-646. |
|
|
Sena L, Zucolotto S, Reginatto F, Schenkel E, De Lima T (2009). Neuropharmacological activity of the pericarp of Passiflora edulis flavicarpa Degener: Putative involvement of C-glycosyl flavonoids. Experimental Biology and Medicine 234(8):967-975. |
|
|
Sharma P, Jha AB, Dubey RS, Pessarakdi M (2012). Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. Journal of Botany 217037. |
|
|
Silva JK, Cazarin CBB, Colomeu TC, Batista AG, Meletti LMM, Paschoal JAR, Júnior SB, Furlan MF, Reyes FGR, Augusto F, Júnior MRM, Zollner RL (2013). Antioxidant activity of aqueous extract of passion fruit (Passiflora edulis) leaves: in vitro and in vivo study. Food Research International 53(2):882-890. |
|
|
Simão MJ, Barbosa TJS, Vianna MG, Garcia R, Mansur E, Ignacio ACPR, Pacheco G (2018). A comparative study of phytoconstituents and antibacterial activity of in vitro derived materials of four Passiflora species. Annals of the Brazilian academy of sciences 90(3):2805-2813. |
|
|
Simirgiotis M, Schmeda-Hirschmann G, Borquez J, Kennelly E (2013). The Passiflora tripartita (Banana Passion) fruit: a source of bioactive flavonoid C-glycosides isolated by HSCCC and characterized by HPLC-DAD-ESI/MS/MS. Molecules 18(2):1672-1692. |
|
|
Škodová-Sveráková I, Záhonová K, Bu?ková B, Füssy Z, Yurchenko V, Lukeš J (2020). Catalase and Ascorbate Peroxidase in Euglenozoan Protists. Pathogens 9(4):317. |
|
|
Smruthi R, Divya M, Archana K, Maddaly R (2021). The active compounds of Passiflora spp and their potential medicinal uses from both in vitro and in vivo evidences. Journal of advanced Biomedical and Pharmaceutical Sciences 4(1):45-55. |
|
|
Viana ML, Costa AM, Celestino SMC (2016). Informations for the composition of nutritional table of the passion fruit pulp BRS Pérola do Cerrado. Boletim de Pesquisa e Desenvolvimento - Embrapa Cerrados 335:17-ref.18. |
|
|
Vianna MG, Garcia RO, Mansur E, Engelmann F, Pacheco G (2019). Oxidative stress during the cryopreservation of Passiflora suberosa L. shoot tips using the V-Cryo-plate technique: determination of the critical stages of the protocol. Plant Cell, Tissue and Organ Culture 139(2):369-379. |
|
|
Wang J, Liu H, Zhao J, Gao H, Zhou L, Zhilong L, Chen Y, Sui P (2010b). Antimicrobial and antioxidant activities of the root bark essential oil of Periploca sepium and its main component 2-hydroxy-4-methoxybenzaldehyde. Molecules 15(8):5807-5817. |
|
|
Wang J, Zhao J, Liu H, Zhou L, Liu Z, Wang J, Han J, Yu Z, Yang F (2010a). Chemical analysis and biological activity of the essential oils of two Valerianaceous species from China: Nardostachys chinensis and Valeriana officinalis. Molecules 15(9):6411-6422. |
|
|
Wosch L, dos Santos KC, Imig DC, Santos CAM (2017). Comparative study of Passiflora taxa leaves: II. A chromatographic profile. Brazilian Journal of Pharmacognosy 27(1):40-49. |
|
|
Yalç?n S, Yalç?nkaya S, Ercan Fn (2021). In silico detection of inhibitor potential of Passiflora compounds against SARS-Cov-2(Covid-19) main protease by using molecular docking and dynamic analyses. Journal of Molecular Structure 1240:130556. |
|
|
Yu W, Liu Y, Song L, Jacobs DF, Du X, Ying Y, Shao Q, WU J (2017). Effect of differential light quality on morphology, photosynthesis, and antioxidant enzyme activity in Camptotheca acuminata Seedlings. Journal Plant Growth Regulation 36(1):148-160. |
|
|
Zucolotto S, Reginatto F, Ramos F, Castellanos L, Duque C, Schenkel E (2011). Analysis of C-glycosyl flavonoids from south American Passiflora species by HPLC-DAD and HPLC-MS. Phytochemical Analysis 23(3):232-239. |
|
|
Zucolotto S, Goulart S, Montanher A, Reginatto F, Schenkel E, Fröde T (2009). Bioassay-guided isolation of anti-inflammatory C-glucosylflavones from Passiflora edulis. Planta Medica 75(11):1221-122. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


