Full Length Research Paper

ABSTRACT
Cognitive decline induced by oxidative brain damage is the critical pathological hallmark of Alzheimer’s disease. Studies have shown that individual administration of Boswellia serrata Roxb (BS), Zingiber officinale Roscoe (ZO) and Ginkgo biloba L. (GB) extracts improved memory and learning through a different mechanism of actions. This study aims to compare the individual effects of each extract with their co-administration on memory impairment induced by scopolamine in mice. Memory dysfunction was induced by a single dose of scopolamine (1 mg/kg, i.p) and extracts were administered intraperitoneally in different doses for one week. Memory performance of the mice was evaluated using the object recognition task (ORT) and passive avoidance test (PAT). The outcomes from ORT demonstrated that, ZO and GB extracts at 200 mg/kg and BS extract just in combination group significantly enhanced (by 95%) the memory loss induced by scopolamine (P < 0.05). On the other hand, PAT results revealed that BS extract at 60 and 90 mg/kg, ZO and GB extracts at 200 mg/kg and their combinations noticeably improved the latency time (by 80%). Although in PAT, co-administration of extracts was more effective than either alone doses in augmenting of the memory function, ORT results showed no considerable differences.
Key words: Boswellia serrata, Zingiber officinale, Ginkgo biloba, extracts, memory, scopolamine.
INTRODUCTION
Alzheimer's disease (AD) is an irreversible progressive neurodegenerative disease that is characterized by loss of memory, accompanied by behavioral conflicts with a huge impact on daily activities (Barker et al., 2002; Wilson et al., 2012). AD pathology includes oxidative stress, neuroinflammation and neuronal death, which consequently leads to memory impairment (Kanninen et al., 2011; Hampel et al., 2019; Sivera et al., 2019).
At present, there is no suitable cure for dementia-related disorders and existing medications come with many adverse effects without providing satisfactory improvements. Plant-derived compounds have played an important part as a source in formulating synthetic drugs.
Boswellia serrata Roxb (Burseraceae) (BS) is a genus of trees known for its aromatic resins especially triterpenes that are antioxidant agents due to their inhibitory activity of the synthesis of endogenous leukotrienes (Sayed and El Sayed, 2016; Iram et al., 2017; Byler and Setzer, 2018). In addition to this, essential oils of this plant have a relaxing effect on vascular smooth muscles, especially cerebral arteries, and eliminate vascular spasm which leads to a better blood supply to brain tissues and cells (Gomaa et al., 2019).
Zingiber officinale Roscoe (Zingiberaceae) (ZO) contains several bioactive phytochemicals, mainly sesquiterpenes and monoterpenes as well as volatile oils (da Silveira Vasconcelos et al., 2019). Previous studies have confirmed that ginger root extract alters the expression of precursor genes in proteins that induce various oxidative reactions (Azam et al., 2014). Furthermore, the extract of this plant has anti-hypoxic and prevention of cerebral ischemia activity due to its ability to cross the blood-brain barrier (Tung et al., 2017).
The flavonoids and terpenoids of Ginkgo biloba L. (Ginkgoaceae) (GB) have different neuroprotective effects (Singh et al., 2019). Antioxidant and free radical scavenger properties of ginkgo extract are mainly related to its flavonoids (Wu et al., 2016). Terpenoids on the other hand, have been shown to reduce ischemic neurotoxicity and prevent glutamate-induced toxic irritability (Li et al., 2017). Besides, the extract of this plant disrupts beta-amyloid production and prevents amyloid-induced neurotoxicity (Verma et al., 2020).
Various animal models have been developed to evaluate dementia based on their diverse pathophysiological basis (Newman et al., 2017). Scopolamine, a nonselective antimuscarinic agent, leads to progressive impairment of learning and memory principally by blocking central cholinergic signaling (Muhammad et al., 2019). It is a well-known phenomenon that reactive oxygen species (ROS) generated by scopolamine results in oxidative stress, a critical factor that results in AD-like dementia (Ko et al., 2018; Skalicka-Wozniak et al., 2018).
As stated above, there is a considerable body of evidence that supports the individual action of BS, ZO and GB extracts in improving memory in both normal brains and impaired ones (Jivad and Rabiei, 2014). However, the effect of simultaneous administration of these plant extracts in scopolamine-induced memory dysfunction has not been investigated before. Therefore, the present study was designed to evaluate the possible ameliorative effects of these plant extracts alone or in combination on cognitive impairment induced by scopolamine in mice.
MATERIALS AND METHODS
Scopolamine was purchased from Osve Pharmaceutical Company (Tehran, Iran) and prepared freshly before use in normal saline. ZO, GB and BS dried extracts were supplied by Goldarou Pharmaceutical Company (Isfahan, Iran) and were dissolved in Tween 80 and normal saline at a ratio of 5:95 respectively. Rivastigmine 1.5 mg capsules (Exelon, Novartis, Switzerland) were purchased from local sources and were dissolved in normal saline. Shuttle box was purchased from Teknik-Azma (Tabriz, Iran). Drugs were injected intraperitoneally (i.p) in a volume of 10 ml/kg.
Animals
Male Syrian mice were obtained from Pasture Institute, Iran (weighing 25-30 gram) and kept under a standard condition of controlled temperature (25 and lightning (12/12 light/dark cycle) in the polyacrylic cages. Mice were randomly housed in 6 per cage and were given free access to food and water. Two hours before experiments, mice were acclimated to the main environment. To avoid the diurnal cycle, all injections and tests were carried out between 8:00 AM to 1.00 P.M. All housing and procedures associated with this experiment were approved by the Animal Research Ethics Committee of Isfahan University of Medical Science (ethical approval ID: IR.MUI.RESEARCH.REC.1398.589) and performed by National Institute of Health Guide for the Care and Use of Laboratory Animals.
Experimental process
Animals were divided randomly into 18 groups (at least 6 in each group) including:
Group 1: Control (normal saline for one week)
Group 2: Scopolamine, (1 mg/kg in all treatments except control group)
Group 3: Positive control (rivastigmine 2 mg/kg for one week)
Groups 4 and 5: ZO extract (150 and 200 mg/kg, respectively for one week)
Groups 6 and 7: BS extract (60 and 90 mg/kg, respectively for one week)
Group 8 and 9: GB extract (100 and 200 mg/kg, respectively for one week)
Group 10, 11 and 12: Combination of ZO/BS/GB extracts at 100/45/50, 150/45/50 and 200/45/50 mg/kg respectively, for one week.
Group 13, 14 and 15: Combination of ZO/BS/GB extracts at 100/45/50, 100/60/50, 100/90/50 mg/kg respectively, for one week.
Group 16, 17 and 18: Combination ZO/BS/GB extracts at 100/45/50, 100/45/100 and 100/45/200 mg/kg respectively, for one week.
At the end of the experiment (on day 7), scopolamine was injected in every group (except control) of animals one hour before the ORT and PAT training trials. ORT and PAT training trial were performed on day 7, continued by test trials on day 8 (scopolamine was injected 1 h before test trials). Animals were sacrificed at the end of the study (Table 1).
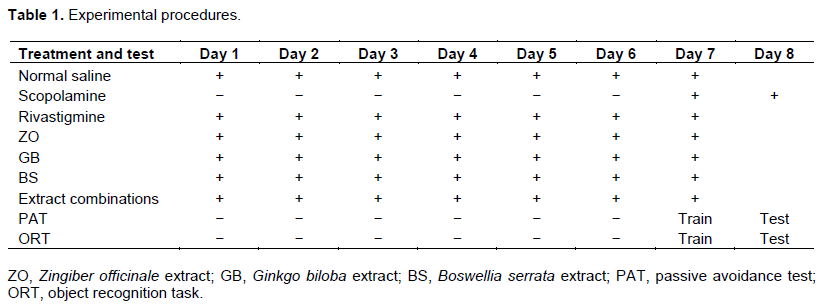
In this study, mice underwent behavioral tests after receiving scopolamine, rivastigmine and extracts of three plants in different doses separately and in combination (Receive (+), do not receive (-)).
Object recognition task (ORT)
The ORT was carried out according to Leuptow study (Lueptow, 2017), with some modifications. This test is based on the animal's natural desire to touch a new object as opposed to a familiar object. This task does not require external stimuli, reward, or punishment. The test was performed in a circular field with a diameter of 32 cm and a height of 20 cm (Figure 1). To facilitate the assessments, a video tracking camera was set on the top of the box. One hour before the training trial, mice acclimated to the empty field for 10 min and then were brought back to their cage. In the training trial, two identical objects were placed in the field within 10 cm distance from each other and 5 cm distance from the wall. The objects were made of plastic (4 cm height). Mice then were put back in the field (in the position in which their face opposes the objects) for exploration. Exploration was defined as sniffing or touching the objects with the nose or forepaws. Sitting on the object was not considered exploratory behavior. The training trial terminated in 3 min or when the mice explored two objects for at least 5 s. The animals that did not reach the exploration time of five seconds were eliminated from the study. The mice were then brought back to their cage and after 24 h the test trial was carried out by presenting a familiar object and a novel one (with the same size as the familial object and different shape). The novel object was made of glass. As soon as the mice explored two objects for 25 s or 5 min the test was completed. From several identical objects, each time a pair of objects was chosen randomly for each mouse. After each trial, all the objects and the field were cleaned with ethanol 70%. Finally, the discrimination index (DI) and the recognition index (RI) were recorded. Percentages of DI and RI were calculated as follows where N and R are the exploration time of the novel and familiar objects, respectively.
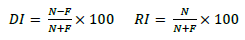
Passive avoidance test (PAT)
The test was performed using the method described by Moosavi et al. with minor modifications (Moosavi et al., 2018). Passive avoidance is a fear-motivated avoidance task used to assess short- and long-term memory in which the mice learn to refrain from stepping through a door to a dark compartment in which they were previously punished. This test was carried out in the apparatus named the shuttle box made by Teknik-Azma, Iran. The device consists of two chambers of the same size (25 × 25 × 20 cm) that are separated by a guillotine gate (6 × 7 cm) (Figure 2). One of the chambers is dark and the other one is illuminated with LED light. The floors of the dark and light chambers are made of stainless steel bars (3 mm diameter) that are spaced 1 cm apart. The bars of the darkroom are attached to the electronic supply. The animal’s position is detected by a high sensitivity photoelectric transducer. The shuttle box was controlled through a microprocessor (SB100) base controller with a touch screen. Before beginning the training pilot, mice were brought into the device to liberally get accustomed to the chambers. In the train session, rodents were placed in the bright room while the door was closed. After 10 s the guillotine gate opened and the rodent out of curiosity went through the gate and entered the darkroom. The latency of entering the brightened chamber was recorded as latency time (The rodents who did not enter the darkroom after 180 s were excluded from the study). At this time, the door sensor detected the passage of the mouse and closed the door. Then after 3 s, the shock was applied to the mouse by the pitch of the dark chamber (1 mA for 3 s). The mouse was then expelled from the device and transferred to its cage until the test phase. The test session was performed 24 h after the train session. In the test session, the rodent was placed in the light chamber while the door was closed. After 10 s the door was opened and the first time that rodent entered the darkroom was recorded as latency time. The number of crossings between chambers and the total time that rodents spent in the dark and light chambers were recorded. Test trial continued for 3 min. The latency time is the most important factor in this experiment which indicated the effect of drugs that improved training of the mouse to overcome its instinct to stay in the dark environment and the escape of light since the punishment of rodents should reverse that natural disposition. After each train and test trial, the chambers were cleaned with ethanol 70%.
Statistics
Statistical analysis for multiple comparisons was conducted by using one-way ANOVA followed by the Tukey POST HOC test for a series of behavior tests with GraphPad Prism software. The data values were defined to be statistically significant at P < 0.05. All data are expressed as mean ± SEM. Student t-Test was used to compare two groups in some experiments.
RESULTS
Object recognition task
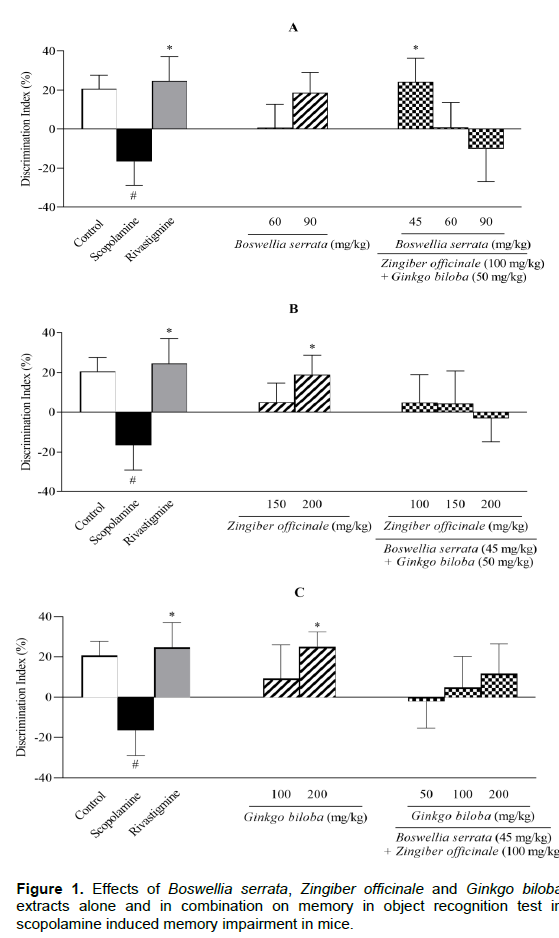
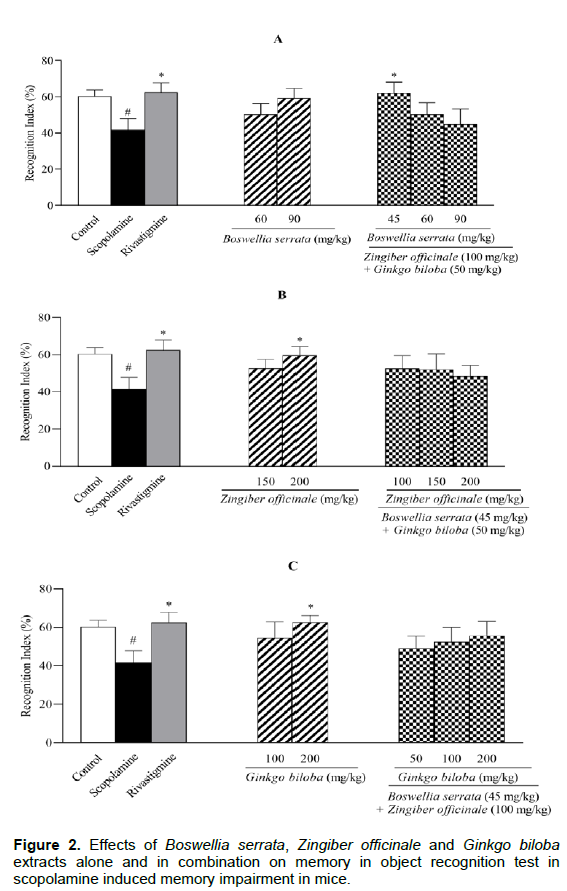
A single dose of scopolamine at 1 mg/kg significantly decreased the discrimination index (DI) (P < 0.05 compared to control group; Figure 1). Rivastigmine at 2 mg/kg reversed the scopolamine action by returning the DI to the control level (P < 0.05). Although BS extract (60 or 90 mg/kg), ZO extract (150 mg/kg) and GB extract (100 mg/kg) increased the DI by 40%, neither were significantly different from scopolamine group. At maximum dose of 200 mg/kg, ZO and GB extracts significantly reversed the scopolamine action (P < 0.05, Figures 1B and 1C). Among the combinational administration of extracts, only ZO/BS/GB at 100/45/50 mg/kg managed to better the scopolamine effects (P < 0.05, Figure 1A). The results of recognition index (RI) were very similar to DI (Figures 2A, B and C). The extent to which the memory was altered in ORT was nearly identical for mono- and combinational extract-treatments.
All groups except control were treated by scopolamine (1 mg/kg). Memory performance was measured by discrimination index (DI) = ((N−F/N+F) × 100) and recognition index (RI) = ((N/N+F) × 100). Results are expressed as mean ± S.E.M (n = 6) and were analyzed by one way ANOVA followed by Tukey POST HOC test and Student t-Test. #P < 0.05 or $P < 0 .01 compared to control group and *P < 0.05 or **P < 0.01 compared to scopolamine group.
Passive avoidance test
The effect of plant extracts on latency time (LT) is shown in Figure 3. Injection of scopolamine resulted in a notable reduction in LT (compared to control group) which was fully reversed by rivastigmine (P < 0.05). A single injection of BS extract (60 or 90 mg/kg) significantly increased the LT by 80% (Figure 3A). In contrast to BS extract, only higher doses of ZO and GB extracts (200 mg/kg) significantly reversed the memory deficit induced by scopolamine (P < 0.05). In combinational treatments, three plant extracts at their minimum effective doses were administered. Administration of ZO/BS/GB extracts at 100/45/50 mg/kg resulted in the increase of the LT to values close to control data (P < 0.01, Figure 3A). Similarly, increase in LT was observed when ZO/BS/GB extracts was combined at different doses of 150/45/50 and 200/45/50 mg/kg (P < 0.05, Figure 3B). Administration of ZO/BS/GB extracts at 100/45/100 and 100/45/200 mg/kg also resulted in more than 80% improvement in LT (P < 0.05, Figure 3C).
All groups except control were treated by scopolamine (1 mg/kg). Memory performance was measured by latency time of entering the dark chamber for the first time in the test session which performed 24 h after train session. Results are expressed as mean ± S.E.M (n = 6) and were analyzed by one way ANOVA followed by Tukey POST HOC test and Student t-Test. #P < 0.05 or $P < 0.01 compared to control group and *P < 0.05 or **< 0.01 compared to scopolamine group.
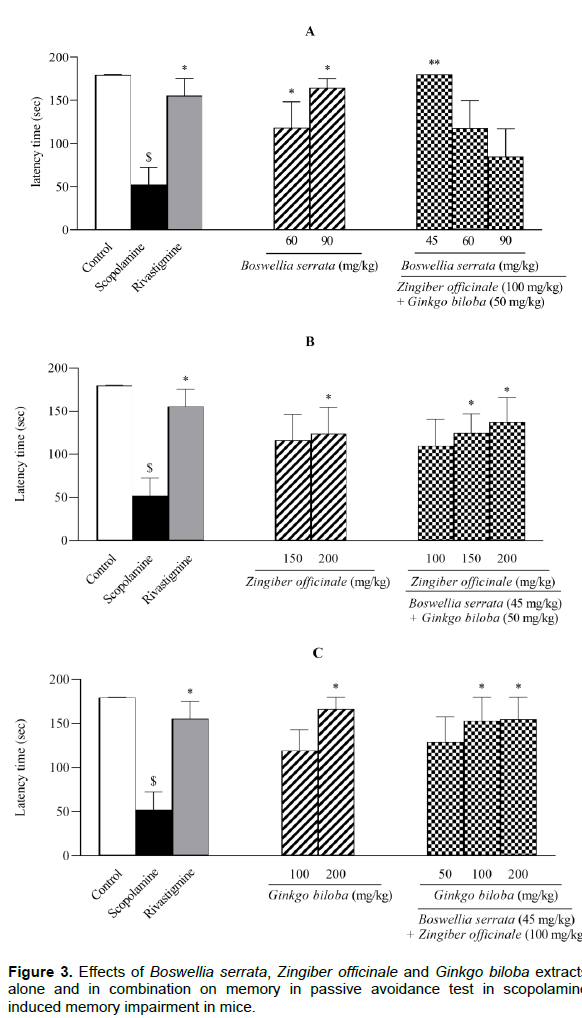
The number of crossings (NC) between the dark and light chambers in the PAT displayed in Figure 4. The NC in scopolamine treated animals was significantly higher than the control group (P < 0.01). Rivastigmine reversed the effect of scopolamine to the control level (P < 0.05). \
Treatment with BS extract (90 mg/kg) significantly decreased the NC to 30% of scopolamine group values (P < 0.05); whereas lower dose of BS extract (60 mg/kg) was ineffective in changing the NC (Figure 4A). The co-administrations of ZO/BS/GB extracts at 100/45/50 and 100/60/50 mg/kg resulted in a significant reduction in the NC (P < 0.01 and P < 0.05 respectively, Figure 4A). Surprisingly, the NC values were almost doubled when ZO/BS/GB extracts was administered at 100/90/50 mg/kg (Figure 4A). Although single injection of ZO extract at 150 or 200 mg/kg and its combination as ZO/BS/GB at 100/45/50 and 150/45/50 mg/kg did not noticeably decline the NC, a significant decrease in NC was found in combination of ZO/BS/GB extracts at 200/45/50 mg/kg in comparison with scopolamine group (P < 0.05, Figure 4B). Furthermore, treated groups of GB extract (200 mg/kg) and its combination of ZO/BS/GB at 100/45/100 and 100/45/200 mg/kg reversed the scopolamine effect and reduced the NC to the control level (P < 0.01); however, GB extract at 100 mg/kg and its combination as ZO/BS/GB at 100/45/50 mg/kg could not accomplish such effect on the NC significantly (Figure 4C).
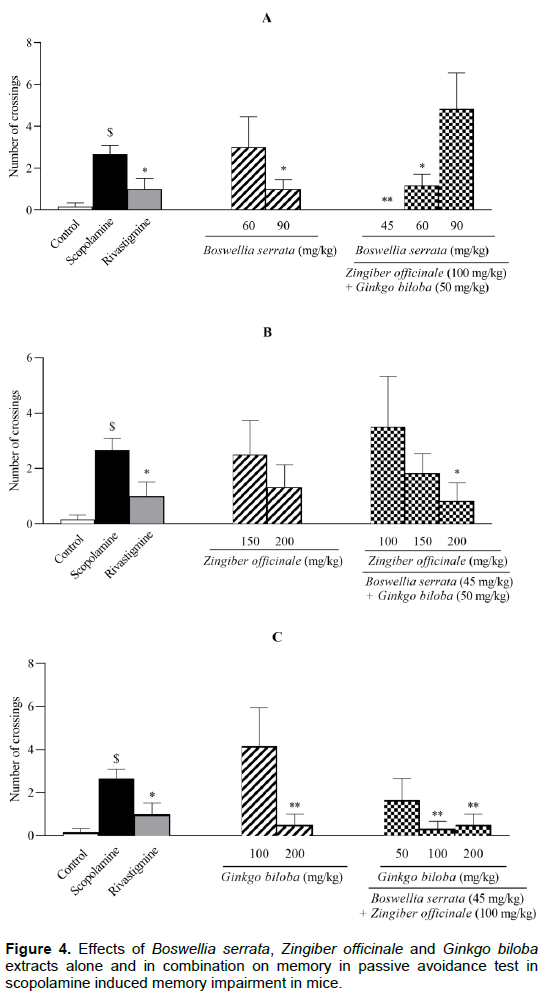
All groups except control were treated by scopolamine (1 mg/kg). Memory performance was measured by number of crossings between the dark and light chambers in the test session which performed 24 h after train session. Results are expressed as mean ± S.E.M (n = 6) and were analyzed by one way ANOVA followed by Tukey POST HOC test and Student t-Test. #P < 0.05 or $P < 0.01 compared to control group and *P < 0.05 or **P < 0.01 compared to scopolamine group.
Figure 5 demonstrated the total amount of time that rodents spent in the dark chamber (DT) in PAT. Administration of scopolamine resulted in the increase of the time was spent by animal in the dark chamber (P < 0.01, compared to control group) and as expected rivastigmine reversed this time to control values (P < 0.05). Administration of BS extract at single dose of 90 mg/kg and its combination as ZO/BS/GB at 100/45/50 and 100/60/50 mg/kg considerably decreased the DT (P < 0.05 and P < 0.01 respectively, Figure 5A). On the other hand, when the dose of BS extract was lowered to 60 mg/kg or co-administrated with other two extracts as ZO/BS/GB (100/90/50 mg/kg), resulted in an increase of 50% compared to scopolamine group in the time was spent by mice in dark chamber (Figure 5A). Moreover, ZO extract at 150 or 200 mg/kg and its combination as ZO/BS/GB (150/45/50 mg/kg) did not reduce the DT; however, ZO/BS/GB extracts at 100/45/50 and 200/45/50 mg/kg decreased that time significantly (P < 0.05, Figure 5B). Among single administrations of the GB extract, dose of 100 mg/kg could not reverse the effect of scopolamine considerably though, GB extract at 200 mg/kg represented notable contrast with scopolamine group (P < 0.05); as well all its combinations at different doses could meaningfully diminish the DT to the control level (P < 0.01, Figure 5C).
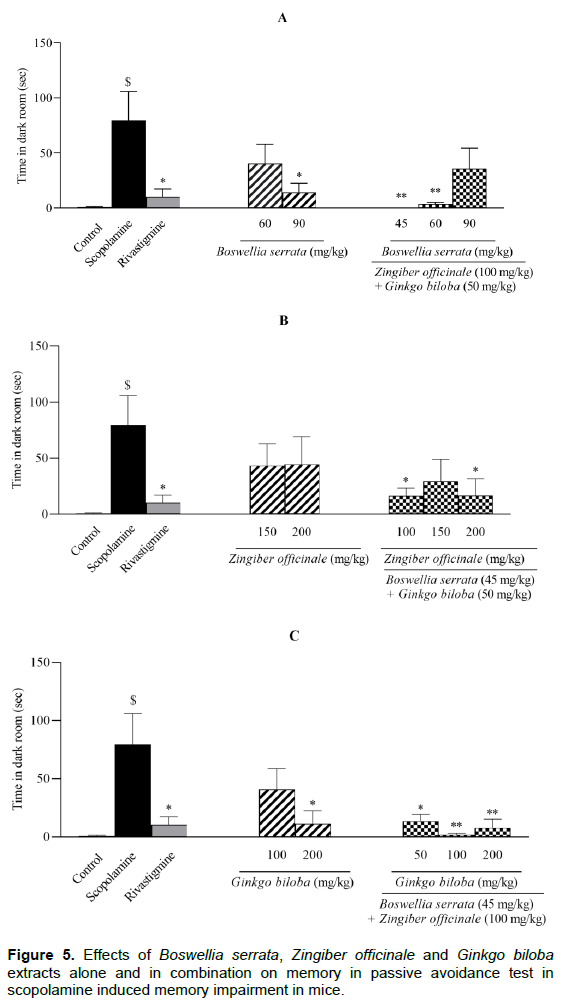
All groups except control were treated by scopolamine (1 mg/kg). Memory performance was measured by total time spent in the dark chamber in the test session which performed 24 h after training session. Results are expressed as mean ± S.E.M (n = 6) and were analyzed by one way ANOVA followed by Tukey POST HOC test and Student t-Test. #P < 0.05 or $P < 0.01 compared to control group and *P < 0.05 or **P < 0.01 compared to scopolamine group.
DISCUSSION
Several preclinical and clinical studies have demonstrated the effectiveness of BS, ZO and GB extracts in varied animal models of memory impairment (Ebrahimpour et al., 2017; El Tabaa et al., 2017; Kim et al., 2018). However, none have examined the combinational effects of these three substances on memory functions. Therefore, the present study was aimed to compare the possible ameliorative effects of three plant extracts, namely BS, ZO and GB, alone or in combination on cognitive impairment induced by scopolamine in mice.
In this study, a single i.p injection of scopolamine led to a cognitive deficit in both models of memory assessment, namely the passive avoidance test (PAT) and object recognition task (ORT). In PAT paradigm, the scopolamine group showed an expected performance in learning and memory test, as mice took a shorter time (lower latency) to enter the dark chamber. The same phenomenon was observed in ORT since discrimination index (DI) and recognition index (RI) were significantly affected by scopolamine. Based on studies were carried out on rodents brain, scopolamine caused impairment in learning and memory through degeneration and dysfunction of cortical cholinergic neurons which are the result of several unexpressed genes related to muscarinic receptor signaling pathways, apoptosis and cell differentiation (Khakpai et al., 2012). It has also been reported that the memory impairment effect of scopolamine could be related to a high level of lipid peroxidation and a low amount of antioxidants in the mice brains (Wong-Guerra et al., 2017).
A significant decrease in DI and RI values which were observed in animals treated with scopolamine in ORT model was completely reversed by administration of rivastigmine. In PAT, administration of rivastigmine also improved the latencies that were substantially lowered by scopolamine. This is an expected effect from a drug with anticholinesterase activity. Rivastigmine has been shown
in number of studies that could improve the cognition deficit in different types of memory (immediate, long term and short term memory) and decreased the acetyl cholinesterase activity in the cerebral cortex, hippocampus, and striatum (Yanev et al., 2015; Gawel et al., 2016; Gothwal et al., 2019; Ray et al., 2020).
Single administration of BS, ZO and GB extracts for one week, dose-dependently improved short-term memory (STM) and long-term memory (LTM) deficit in both paradigms. In ORT, the effect of different doses of combined extracts (except the combination of ZO/BS/GB at 100/45/50 mg/kg) was no more effective than either alone injections which indicated a lack of synergistic or even additive effects. On the other hand, in the PAT model, the combination of extracts represented more valuable effects in ameliorating the latency time. In addition, combinations of BS, GB and ZO extracts at lower doses represented a more considerable decrease in numbers of crossings which specified the synergistic effect. When the total time that rodents spent in the dark chamber was studied, rivastigmine significantly reversed the scopolamine effects and the co-administration of the extracts enhanced memory more excellently than each agent alone.
The results that have been obtained from the individual administrations of extracts matched with previous reports. Memory enhancement by BS, ZO and GB extracts has been proved that related to several factors. BS extract affected memory function through several mechanisms such as cholinergic pathway, reducing free radical, decrease cerebral oxidative stress and decline glutamate uptake (Rajabian et al., 2016). ZO extract has been shown to alter the expression of precursor genes in proteins that induce various oxidative reactions. Furthermore, the extract of this plant represented antioxidant and anti-hypoxic activities and also could prevent cerebral ischemic because of its ability to cross the blood-brain barrier (Okesola et al., 2019). Additionally, GB extract could reduce ischemic neurotoxicity and prevent glutamate-induced toxic irritability. The extract of this plant has been evidenced properties such as disrupting beta-amyloid production and prevention of amyloid-induced neurotoxicity which are more specifically related to memory dysfunction (Ribeiro et al., 2016).
Despite the wide range of actions by BS, ZO and GB extracts, the co-administration of these three substances revealed little difference from individual injections in the ORT model. In contrast, more effective improvement in memory function was observed in PAT model after combinational therapy. The reason for observing such differences in these two models could be due to several factors. Firstly, this might be due to the different types of memory that are assessed in these two behavioral experiments. A suggested categorization of memory divides it based on various criteria such as its duration (STM vs. LTM), content (explicit vs. implicit) and motive (appetitive/reward vs. aversive) (Barros et al., 2003).
Results from PAT latency time expressed that STM and LTM were enhanced equally. STM is not the early phase of LTM because they process independently through parallel paths. The kind of memory that is assessed in ORT is different in motive and content from PAT; PAT evaluates the implicit memory while ORT assesses explicit one (Neto et al., 2008). In PAT, rodents learn to avoid an aversive stimulus (electric foot-shock) by inhibiting a response so this performance is related to aversive memory. Secondly, different parts of the brain might be involved in these two tests. ORT is believed to related to Para hippocampal regions (perirhinal, entorhinal and inferior temporal cortices), whereas, in PAT amygdala and insular cortex are thought to play an important role (Winters et al., 2008). Finally, according to some studies one-trial object recognition test sometimes may not be able to assess novelty. This discrimination could be construed in two ways; animals could explore each object equally because they have been recognized as the novel or as familiar. That model also could not assess the measure of the memory when the animal faced the object and so it is hard to evaluate the strength of the memory (Antunes and Biala, 2012).
CONCLUSION
Combination therapies could in some cases increase the probability of obtaining a new compound with better efficacy, decreased toxicity and reduced drug resistance. Both single and multi-plant extract injections in this study were effective in improving memory impairment in mice. However, the combination of BS, ZO and GB extracts implied more reliable improve of memory rather than individual extract administration in PAT model. In ORT model, nevertheless, single-drug treatments were as effective as combinational ones. Due to the facts that were discussed, it could be easily understand that the synergistic actions that are typically expected to occur by drug combinations may not arise in every case and test paradigm.
CONFLICTS OF INTERESTS
The author has not declared any conflict of interests.
ACKNOWLEDGMENT
This research was supported by the Research Department of Isfahan University of Medical Sciences. We are grateful to Mr. H.R. Sharifi Jabali for assisting in housing and feeding the animals.
REFERENCES
|
Antunes M, Biala G (2012). The novel object recognition memory: Neurobiology, test procedure, and its modifications. Cognitive Processing 13(2):93-110. |
|
|
Azam F, Amer AM, Abulifa AR, Elzwawi MM (2014). Ginger components as new leads for the design and development of novel multi-targeted anti-alzheimer's drugs: A computational investigation. Drug Design, Development And Therapy 8:2045. |
|
|
Barker WW, Luis CA, Kashuba A, Luis M, Harwood DG, Loewenstein D, Waters C, Jimison P, Shepherd E, Sevush S (2002). Relative |
|
|
frequencies of alzheimer disease, lewy body, vascular and frontotemporal dementia, and hippocampal sclerosis in the state of florida brain bank. Alzheimer Disease and Associated Disorders 16(4):203-212. |
|
|
Barros DM, Izquierdo LA, Medina JH, Izquierdo I (2003). Pharmacological findings contribute to the understanding of the main physiological mechanisms of memory retrieval. Current Drug Targets-CNS amd Neurological Disorders 2(2):81-94. |
|
|
Byler KG, Setzer WN (2018). Protein targets of frankincense: A reverse docking analysis of terpenoids from boswellia oleo-gum resins. Medicines 5(3):96. |
|
|
da Silveira Vasconcelos M, Mota EF, Gomes-Rochette NF, Nunes-Pinheiro DCS, Nabavi SM, de Melo DF (2019). Ginger (zingiber officinale roscoe). In: Nonvitamin and nonmineral nutritional supplements. Elsevier, pp. 235-239. |
|
|
Ebrahimpour S, Fazeli M, Mehri S, Taherianfard M, Hosseinzadeh H (2017). Boswellic acid improves cognitive function in a rat model through its antioxidant activity:-neuroprotective effect of boswellic acid. Journal of Pharmacopuncture 20(1):10. |
|
|
El Tabaa MM, Sokkar SS, Ramadan ES, Abd El Salam IZ, Zaid A (2017). Neuroprotective role of ginkgo biloba against cognitive deficits associated with bisphenol a exposure: An animal model study. Neurochemistry International 108:199-212. |
|
|
Gawel K, Labuz K, Gibula-Bruzda E, Jenda M, Marszalek-Grabska M, Filarowska J, Silberring J, Kotlinska JH (2016). Cholinesterase inhibitors, donepezil and rivastigmine, attenuate spatial memory and cognitive flexibility impairment induced by acute ethanol in the barnes maze task in rats. Naunyn-Schmiedeberg's Archives of Pharmacology 389(10):1059-1071. |
|
|
Gomaa AA, Makboul RM, Al-Mokhtar MA, Nicola MA (2019). Polyphenol-rich boswellia serrata gum prevents cognitive impairment and insulin resistance of diabetic rats through inhibition of gsk3β activity, oxidative stress and pro-inflammatory cytokines. Biomedicine and Pharmacotherapy 109: 281-292. |
|
|
Gothwal A, Singh H, Jain SK, Dutta A, Borah A, Gupta U (2019). Behavioral and biochemical implications of dendrimeric rivastigmine in memory-deficit and alzheimer's induced rodents. ACS Chemical Neuroscience 10(8):3789-3795. |
|
|
Hampel H, Mesulam MM, Cuello A, Khachaturian A, Vergallo A, Farlow M, Snyder P, Giacobini E, Khachaturian Z, Group CSW (2019). Revisiting the cholinergic hypothesis in alzheimer's disease: Emerging evidence from translational and clinical research. The Journal of Prevention Of Alzheimer's Disease 6(1):2-15. |
|
|
Iram F, Khan SA, Husain A (2017). Phytochemistry and potential therapeutic actions of boswellic acids: A mini-review. Asian Pacific Journal of Tropical Biomedicine 7(6):513-523. |
|
|
Jivad N, Rabiei Z (2014). A review study on medicinal plants used in the treatment of learning and memory impairments. Asian Pacific Journal of Tropical Biomedicine 4(10):780-789. |
|
|
Kanninen K, White AR, Koistinaho J, Malm T (2011). Targeting glycogen synthase kinase-3β for therapeutic benefit against oxidative stress in alzheimer's disease: Involvement of the nrf2-are pathway.International Journal of Alzheimer's Disease, 2011. |
|
|
Khakpai F, Nasehi M, Haeri-Rohani A, Eidi A, Zarrindast MR (2012). Scopolamine induced memory impairment; possible involvement of nmda receptor mechanisms of dorsal hippocampus and/or septum. Behavioural Brain Research 231(1):1-10. |
|
|
Kim CY, Seo Y, Lee C, Park GH, Jang JH (2018). Neuroprotective effect and molecular mechanism of [6]-gingerol against scopolamine-induced amnesia in c57bl/6 mice. Evidence-based Complementary and Alternative Medicine: eCAM 2018. |
|
|
Ko YH, Kim SY, Lee SY, Jang CG (2018). 6, 7, 4′-trihydroxyisoflavone, a major metabolite of daidzein, improves learning and memory via the cholinergic system and the p-creb/bdnf signaling pathway in mice. European Journal of Pharmacology 826:140-147. |
|
|
Li ZY, Chung YH, Shin EJ, Dang DK, Jeong JH, Ko SK, Nah SY, Baik TG, Jhoo JH, Ong WY (2017). Yy-1224, a terpene trilactone-strengthened ginkgo biloba, attenuates neurodegenerative changes induced by β-amyloid (1-42) or double transgenic overexpression of app and ps1 via inhibition of cyclooxygenase-2. Journal of Neuroinflammation 14(1):1-22. |
|
|
Lueptow LM (2017). Novel object recognition test for the investigation of learning and memory in mice. Journal of Visualized Experiments (126):e55718. |
|
|
Moosavi M, SoukhakLari R, Moezi L, Pirsalami F (2018). Scopolamine-induced passive avoidance memory retrieval deficit is accompanied with hippocampal mmp2, mmp-9 and mapks alteration. European Journal of Pharmacology 819:248-253. |
|
|
Muhammad T, Ali T, Ikram M, Khan A, Alam SI, Kim MO (2019). Melatonin rescue oxidative stress-mediated neuroinflammation/ neurodegeneration and memory impairment in scopolamine-induced amnesia mice model. Journal of Neuroimmune Pharmacology 14(2):278-294. |
|
|
Neto SPD, Carneiro BTS, Valentinuzzi VS, Araújo JF (2008). Dissociation of the circadian rhythm of locomotor activity in a 22 h light-dark cycle impairs passive avoidance but not object recognition memory in rats. Physiology and Behavior 94(3):523-527. |
|
|
Newman M, Kretzschmar D, Khan I, Chen M, Verdile G, Lardelli M (2017). Animal models of alzheimer's disease. In: Animal models for the study of human disease. Elsevier pp. 1031-1085. |
|
|
Okesola MA, Ajiboye BO, Oyinloye BE, Ojo OA (2019). Neuromodulatory effects of ethyl acetate fraction of zingiber officinale roscoe extract in rats with lead-induced oxidative stress. Journal of Integrative Medicine 17(2):125-131. |
|
|
Rajabian A, Boroushaki MT, Hayatdavoudi P, Sadeghnia HR (2016). Boswellia serrata protects against glutamate-induced oxidative stress and apoptosis in pc12 and n2a cells. DNA and Cell Biology 35(11):666-679. |
|
|
Ray B, Maloney B, Sambamurti K, Karnati HK, Nelson PT, Greig NH, Lahiri DK (2020). Rivastigmine modifies the α-secretase pathway and potentially early alzheimer's disease. Translational Psychiatry 10(1):1-17. |
|
|
Ribeiro ML, Moreira LM, Arçari DP, Dos Santos LF, Marques AC, Pedrazzoli Jr J, Cerutti SM (2016). Protective effects of chronic treatment with a standardized extract of ginkgo biloba l. In the prefrontal cortex and dorsal hippocampus of middle-aged rats. Behavioural Brain Research 313:144-150. |
|
|
Sayed AS, El Sayed NSED (2016). Co-administration of 3-acetyl-11-keto-beta-boswellic acid potentiates the protective effect of celecoxib in lipopolysaccharide-induced cognitive impairment in mice: Possible implication of anti-inflammatory and antiglutamatergic pathways. Journal of Molecular Neuroscience 59(1):58-67. |
|
|
Singh SK, Srivastav S, Castellani RJ, Plascencia-Villa G, Perry G (2019). Neuroprotective and antioxidant effect of ginkgo biloba extract against ad and other neurological disorders. Neurotherapeutics pp. 1-9. |
|
|
Sivera R, Delingette H, Lorenzi M, Pennec X, Ayache N, Alzheimer's Disease Neuroimaging Initiative (2019). A model of brain morphological changes related to aging and alzheimer's disease from cross-sectional assessments. NeuroImage 198:255-270. |
|
|
Skalicka-Wozniak K, Budzynska B, Biala G, Boguszewska-Czubara A (2018). Scopolamine-induced memory impairment is alleviated by xanthotoxin: Role of acetylcholinesterase and oxidative stress processes. ACS Chemical Neuroscience 9(5):1184-1194. |
|
|
Tung BT, Thu DK, Thu NTK, Hai NT (2017). Antioxidant and acetylcholinesterase inhibitory activities of ginger root (zingiber officinale roscoe) extract. Journal of Complementary and Integrative Medicine 14(4). |
|
|
Verma S, Sharma S, Ranawat P, Nehru B (2020). Modulatory effects of ginkgo biloba against amyloid aggregation through induction of heat shock proteins in aluminium induced neurotoxicity. Neurochemical Research 45(2):465-490. |
|
|
Wilson RS, Segawa E, Boyle PA, Anagnos SE, Hizel LP, Bennett DA (2012). The natural history of cognitive decline in alzheimer's disease. Psychology and Aging 27(4):1008. |
|
|
Winters BD, Saksida LM, Bussey TJ (2008). Object recognition memory: Neurobiological mechanisms of encoding, consolidation and retrieval. Neuroscience and Biobehavioral Reviews 32(5):1055-1070. |
|
|
Wong-Guerra M, Jiménez-Martin J, Pardo-Andreu GL, Fonseca-Fonseca LA, Souza DO de Assis AM, Ramirez-Sanchez J, Del Valle RMS, Nuñez-Figueredo Y (2017). Mitochondrial involvement in memory impairment induced by scopolamine in rats. Neurological Research 39(7):649-659. |
|
|
Wu Y, Sun J, George J, Ye H, Cui Z, Li Z, Liu Q, Zhang Y, Ge D, Liu Y (2016). Study of neuroprotective function of g inkgo biloba extract (eg b761) derived?flavonoid monomers using a three?dimensional stem cell?derived neural model. Biotechnology Progress 32(3):735-744. |
|
|
Yanev PG, Dimitrova DS, Getova-Spassova DP (2015). Effects of rivastigmine and memantine alone and in combination on learning and memory in rats with scopolamine-induced amnesia. Open Medicine 1(open-issue). |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


