Full Length Research Paper

ABSTRACT
Studies have shown arbuscular mycorrhizal fungi (AMF) enhance phosphorus (P) uptake and drought tolerance in maize (Zea mays L.) grown in semiarid soils. However, little is known regarding the contribution of AMF to maize treated with different levels of phosphorus and grown in different soil moisture levels. This study was conducted to determine the effects of AMF (Glomus fasciculatum) inoculation on growth and P uptake of maize treated with different levels of soil P and soil moisture. Different P levels (0, 50, and 100 kg P ha-1) were applied on maize grown in soils with and without mycorrhizal fungi, and at different moisture levels producing -0.05, -0.4, -0.8, and -1.5 MPa of drought stress. Increasing P rates significantly (p < 0.05) reduced mycorrhizal colonization. Mycorrhizal colonization was higher under moderate than under lower soil moisture levels. Drought stress × soil P content × AMF inoculation interaction had significant (p < 0.05) effect on maize shoot and root dry weight and tissue P concentration. Overall, results of this study suggest that mycorrhizal inoculation enhances P uptake and maximizes maize biomass under low, moderate, and high soil moisture conditions without P applications. Except for the lowest soil moisture level (-1.5 MPa), mycorrhizal plants produced higher biomass, with greater tissue P content than nonmycorrhizal plants at all soil P and soil moisture levels. These results indicate that establishing efficacious AMF with maize could be an efficient alternative for growers than relying on P fertilizer application and its associated costs and environmental concerns.
Keywords: Arbuscular Mycorrhiza Fungi, Drought Tolerance, Phosphorus Levels, Maize Plants.
INTRODUCTION
Low levels of available phosphorus (P) and moisture limit crop productivity in many arid and semi-arid regions of the world (Mumtaz et al., 2014). Depleted water resources also negatively affect food production (Yang and Zehnder, 2002). Crops may suffer drought conditions in these areas, especially if dependent on rainfall. Irrigation brings additional concerns, such as salinity stress (Albacete et al., 2014). This situation is worsening due to climate change (Chaves and Oliveira, 2004). In many areas, drought is considered the most important abiotic factor restricting plant productivity (Bray, 2004). Any intervention improving drought tolerance will contribute to greater crop production in these areas (Al-Karaki et al., 2004), many of which are projected to experience more drought in coming years (Calvo-Polanco et al., 2016).
A number of studies have investigated mycorrhizal fungi interactions as one way to improve drought tolerance in plants (Xu et al., 2018; Li et al., 2019). Stahl et al. (1998) have reported mycorrhizal big sagebrush (Artemisia tridentata Nutt. ssp. wyomingensis) seedlings had greater survival than non-mycorrhizal seedlings in dry soil conditions. There is also increasing interest in drought tolerance using crops colonized with mycorrhizae (Calvo-Polanco et al., 2016). Al-karaki et al. (2004), determined mycorrhizal inoculation improves growth, yield and nutrient uptake in wheat plant and reduces the effect of drought stress under field condition in semiarid areas. Furthermore, the evidence is strong for the ability of AMF to alleviate drought stress in different crops such as wheat, barley, maize, soybean, strawberry, and onion ((Mena-Violante et al., 2006; Ruiz-Lozano et al., 2015; Yooyongwech et al., 2016; Moradtalab et al., 2019). Arbuscular mycorrhizal fungi (AMF) are known to increase nutrient and water uptake by host plants (Michelsen and Rosendahl, 1990; Manjunath and Habte, 1991; Allen, 2007; Birhane et al., 2012; Kayama and Yamanaka, 2014) but disparate species and geographical isolates of AMF differ in their ability to influence host plant uptake of materials (Rajan et al., 2000; Cavallazzi et al., 2007). The productivity of numerous crop plants grown in soils inhabited with AM fungi have improved under drought stress (Al-Karaki and Al-Raddad, 1997; Al-Karaki and Clark, 1998). AMF increases drought tolerance, in part, by enhancing P uptake (Wu and Zou, 2009b) in soils with low available P (Huang et al., 1985; Nelsen and Safir, 1982).
AMF tends to benefit plants greatest in soils with low P and decreases under increasing soil P (Schubert and Hayman, 1986; Koide and Li, 1990). Many other studies have demonstrated the role of mycorrhizal fungi in plant nutrition, particularly increased P uptake (Marschner and Dell, 1994; Smith et al., 2003, 2011; Liu et al., 2014; Mai et al., 2018; Ping et al., 2014). Koide (1991) found mycorrhizal fungal colonization may decrease at extremely high or low soil P concentrations. In addition, high soil P levels may contribute to low rates or suppressed mycorrhizal plant colonization (Koide, 1991). Additionally, the ability to influence drought tolerance varies with mycorrhizal fungus and host plant (Stahl and Smith, 1984). For example, inoculation of winter wheat plants by G. fasciculatum increased drought tolerance, while inoculation by G. mosseae did not (Allen and Boosalis, 1983). The objective of this study is to examine the influence of an isolate AMF (G. fasciculatum [Thaxter sensu Gred.] Gerdemann & Trappe) from Egypt on biomass production and tissue P concentration of maize grown under a range of soil P content and soil moisture levels.
MATERIALS AND METHODS
Hypotheses were tested in a greenhouse experiment conducted at Omar Al-Mukhtar University-Albida- Libya. The experimental design was completely randomized (CRD) and arranged in 2*3*4 factorial design with three replications, resulting in a total of 72 pots. Three experimental factors included two different levels of AMF inoculation (0, and 350 g), 3 P application rate (0 [P deficient], 50 [P practical], and 100 [P excess]), and 4 moisture regimes (-0.05, -0.4, -0.8, and -1.5 MPa). The arbuscular mycorrhizal fungus used in this study was an isolate of G. fasciculatum obtained from the Faculty of Agriculture at Alexandria University, Alexandria, Egypt. The fungus was isolated from an agricultural soil and included in this study to improve productivity of agricultural plants in libyan soil. There have been very few studies screening AMF symbionts for libyan agriculture.
Soil and seed preparation
Soil used in this study was collected from a depth of 0-20 cm from an agricultural field used for wheat production in Albida, Libya. The soil was air dried and sieved through a 2 mm sieve. Chemical and physical characteristics of the soil are shown in (Table 1). Soil samples were analyzed for pH and electrical conductivity on 1:1 soil: water paste (Hesse, 1971). Soil was extracted with NaHCO3 for determination of available P (Olsen and Sommers, 1982) and organic matter was measured by rapid oxidation (Walkley and Black, 1934). Soil texture analysis was performed by the hydrometer method (Gee and Bauder, 1986), nutrients were determined in soil extraction. Soil was subsequently autoclaved at 121°C for 1 h to determine AMF in preparation for treatments (Powell and Bagyaraj, 1984). Phosphorus was applied as triple super phosphate (46% P2O5). The ‘Gold’ variety of maize seeds was obtained from Alkufra Production Project-Libya. Seeds were surface sterilized with bleach (10% sodium hypochlorite) for 3 min and subsequently rinsed three times with deionized water.
AMF inoculum
The arbuscular mycorrhizal fungus (G. fasciculatum) inoculum was supplied by the Faculty of Agriculture at Alexandria University, Alexandria, Egypt. Initially, AMF inoculum was propagated and increased on sorghum plants in the collected soil for three months prior to the experiment according to procedures described by Walker (1999). Prepared inoculum was composed of soil, spores, extraradical hyphae and infected sorghum root fragments. For AMF positive treatments, soil was added to pots until they were two/third filled; a layer of G. fasciculatum inoculum 2 cm deep was placed on top of the soil and then covered with soil until the pot was full (~10 kg pasteurized soils on a dry weight basis). For non-mycorrhizal treatments, the same procedure was followed except nonmycorrhizal sorghum roots and soil were not added to pots at the time of planting. Approximately 7 seeds were planted by hand over the inoculum and covered with soil. Three weeks after emergence, seedlings were thinned to two per pot.
Irrigation of pots
Water was supplied daily, if needed, to ensure the soil moisture level was maintained at field capacity until seedling emergence, and before drought stress treatments to promote normal growth and allow for AMF infection. Drought stress treatments began three weeks after planting. Soil moisture content was 32, 20.5, 18.8, and 15.9% when soil was exposed to tension of -0.05 MPa, -0.4 MPa, -0.8 MPa, and -1.5 MPa, respectively. Water content was maintained at the target levels by using a Moisture Meter Type HH2 device daily and adding the suitable volume of water. All treatments were maintained under experimental soil moisture conditions in a greenhouse for 8 weeks before harvesting. Plants were grown at ambient temperature (25-35°C) in a greenhouse and day/night cycle of 14-10 h.
Parameter measurements
Plants were harvested eleven weeks after emergence. Shoots and roots were oven dried at (70°C for 48 h) and then weighed. The presence of AMF infection was determined visually by examining plant roots under a microscope using the methods in (Phillips and Hayman, 1970). Root samples were carefully washed, cut into 1 cm long segments, cleared in 10% KOH overnight, acidified in 10% HCl for 1 min, washed three times with water and stained with 0.066% Trypan blue lactophenol. Ten 1 cm long pieces of fine roots were randomly selected and arranged parallel to each other on a microscope slide. From each root sample, the mycorrhizal root colonization was determined microscopically at 100X. Three vision fields were (ca. 1 mm root segments) examined in each 1 cm root section; therefore, 30 field visions were examined for each slide and the infection by AMF was recorded when AMF hyphae, vesicles, or arbuscules were observed. The percentage of infection was calculated as a ratio of the infected to the total number observed. To determine plant tissue P content, plant tissue samples were dried and powdered. For P concentration, dried tissue of 200 mg was digested in a HCIO3-H2O2 (v/v 7:3) mixture for 20 min in a closed chamber. Water was added (25 ml) to digest samples. After extraction, the colorimetric method was used to determine P concentrations spectrophotometrically using the ascorbic acid method (Walsh and Beaton, 1973).
Statistical analysis
To determine interaction effects, three-way analysis of variance was performed in the R statistical language version 3.5.1 (R Core Team, 2018). AMF, phosphorus, and irrigation were fixed factors. For data on AMF root colonization rates, One-way ANOVA was conducted to determine influence of water potential and P rate. Post-hoc treatment comparisons were performed at alpha of 0.05 using Tukey comparison method.
RESULTS
Mycorrhizal colonization
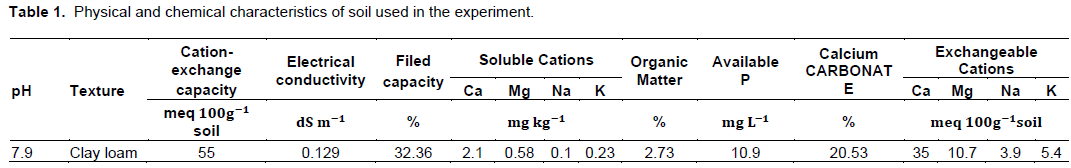
Uninoculated maize plants did not developed mycorrhizal root systems, but maize plants inoculated with 350 g of AMF inoculum formed arbuscular mycorrhizal root systems with between 39 and 75% of 1 cm root segments inhabited by G. fasciculatum. No evidence of AMF were found in the nonmycorrhizal treatments. Mycorrhiza formation by AMF was significantly impacted (p < 0.05) by phosphorus fertilization and soil water potential levels (Figures 1 and 2). Maize plants receiving no fertilizer P had the highest AMF root colonization (Figure 1). Plants grown in soil with P added at 50 and 100 kg ha-1 had lower mean AMF colonization rates than unfertilized plants (Figure 1). Also, soil water content significantly (p < 0.05) affected mycorrhiza formation with the highest rate of root AMF colonization developing at moderate soil water content (-0.4 MPa) (Figure 2). The lowest AMF colonization rate was 74%, observed in the most negative water tension treatment (-1.5 MPa). However, statistical analysis indicates no significant interaction between P and soil moisture tension in the influence on AMF colonization.
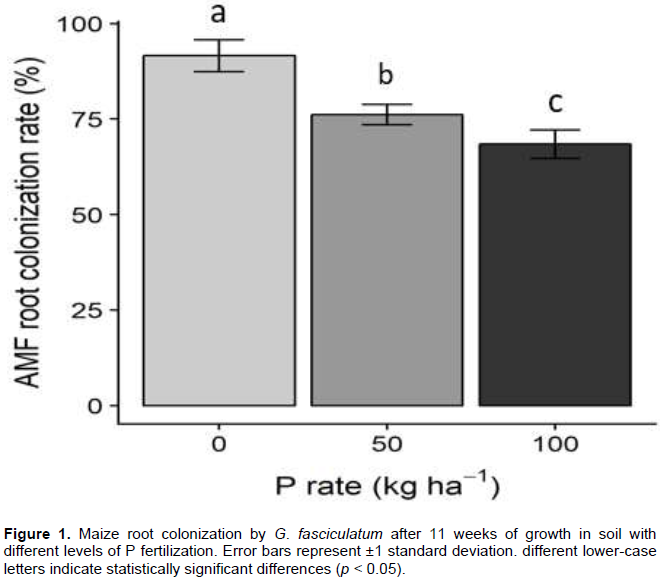
Plant growth
Shoot dry weight
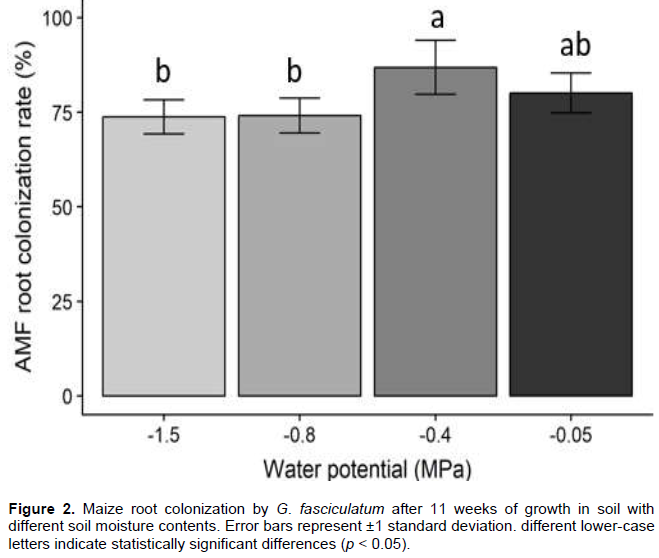
Mycorrhizal maize plants produced more aboveground biomass than non-mycorrhizal plants at all P levels except 50 kg P ha-1 in very dry soil (Figure 3). Statistical analyses indicated shoot dry biomass production by maize was significantly impacted by soil moisture content, P fertilization and AMF, with a significant (p < 0.05) interaction of these factors. Generally, the greatest difference in shoot dry biomass production for mycorrhizal vs. nonmycorrhizal plants occurred when maize plants were grown in soil amended with 50 kg P ha-1, except in very dry soil where the greatest differences were found at 0 kg P ha-1 added P.
Another significant difference was found between growth of mycorrhizal and nonmycorrhizal maize in soil with 0 kg P ha-1 added P in very dry soil (- 1.5 MPa), but no significant difference between growth of mycorrhizal and nonmycorrhizal maize was found in soil amended with 100 kg P ha-1. Adding P increased biomass production by nonmycorrhizal plants in the 3 least moist soils. Mycorrhizal plants showed an inconsistent response in terms of biomass production to increasing soil P.
Under wet soil conditions (- 0.05 MPa), shoot dry biomass production by mycorrhizal plants was consistently higher than by nonmycorrhizal plants across different P fertilizer treatments (Figure 3). A significant (p < 0.05) difference was found between mycorrhizal (greater) and nonmycorrhizal plants in shoot dry biomass production when soil was amended with 50 kg P ha-1. Meanwhile, under 0 and 100 kg P ha-1, there were no significant differences between mycorrhizal and nonmycorrhizal plants (Figure 3). Mycorrhizal plants grown in wet soil (-0.05 MPa) and without P produce greater shoot dry biomass (158.1 g plant-1) than mycorrhizal plants grown in soils treated with 50 (149 g plant-1) and 100 kg P ha-1 (91.3 g plant-1). For nonmycorrhizal plants, the highest shoot dry biomass (115.5 g plant -1) was recorded in soil with no added P.
At moderate water potential of - 0.4 MPa, mycorrhizal plants produced significantly (p < 0.05) higher shoot dry biomass than nonmycorrhizal plants under 0 and 50 kg P ha-1, but there was no significant difference under 100 kg P ha-1 (Figure. 3). Doubling the amount of fertilizer P resulted in less shoot dry biomass production by mycorrhizal plants, but it resulted in more shoot dry biomass production by nonmycorrhizal plants.
In soil maintained at -0.8 MPa of water potential, there was no significant difference in maize biomass production between mycorrhizal and nonmycorrhizal plants at the same P level. Plants grown in 100 kg P ha-1 treatment produced greater shoot dry weight compared to 0 and 50 kg P ha-1 for both mycorrhizal and nonmycorrhizal plants. At soil water tension -1.5 MPa and 0 kg P ha-1, there was a significant (p < 0.05) difference in dry shoot biomass between mycorrhizal and nonmycorrhizal plants, while under 50 and 100 kg P ha-1 there was no significant difference. The highest shoot dry biomass production was 154.27 g plant-1 by mycorrhizal maize at 0 kg P ha-1. Under 50 and 100 kg P ha-1, no significant difference between mycorrhizal and nonmycorrhizal plants under -1.5 MPa soil water content were found.
Root dry weight
Mycorrhizal plants consistently produced at least as much root biomass and often more than nonmycorrhizal plants at all levels of soil P and soil moisture except in the driest soil with highest P content (Figure 4). No conditions in this study produced non-mycorrhizal maize plants with significantly greater amounts of root biomass than mycorrhizal plants. Statistical analyses indicated root biomass production by maize was significantly impacted by soil moisture content, P fertilization rates and AMF, with significant (p < 0.05) interactions of these factors. The greatest differences in root dry biomass production for mycorrhizal vs nonmycorrhizal plants were observed when soil was amended with 0 and 50 kg P ha-1, except in -0.8 MPa soil water content, where the greatest differences were developed at 100 and 0 kg P ha-1 added P, respectively.
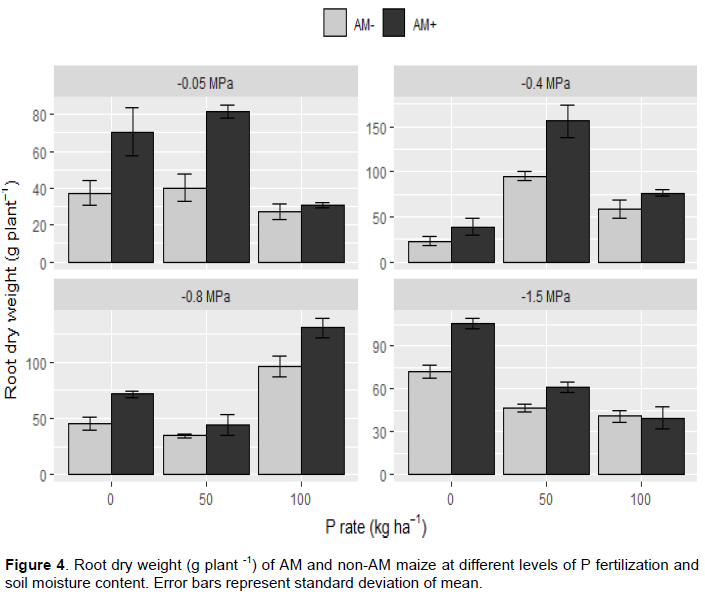
Under wet soil conditions (-0.05 MPa), root dry biomass production by mycorrhizal plants was significantly (p < 0.05) greater than nonmycorrhizal plants in 0 and 50 kg P ha-1 levels, but when soil was amended with 100 kg P ha-1, the difference was not significant. The greater difference in root dry biomass production by mycorrhizal vs nonmycorrhizal plants in wet soil was observed when plants were grown with 50 kg P ha-1 added P, whereas no significant difference between mycorrhizal and nonmycorrhizal plants was observed in soil with 100 kg P ha-1 added P. Root dry biomass production by mycorrhizal plants was 81.7 g plant-1, while nonmycorrhizal plants produced less than half as much root dry biomass (40.1 g plant-1) in soil fertilized with 50 kg P ha-1. Unexpectedly, the lowest root dry biomass production was in soil amended with 100 kg P ha-1 for both mycorrhizal plants and nonmycorrhizal plants, which was 30.66 and 27.17 g plant-1, respectively.
At moderate soil water content (-0.4 MPa), mycorrhizal plants produced significantly (p < 0.05) more root dry biomass than nonmycorrhizal plants in soil amended with 50 and 100 kg P ha-1, while in soil with no added P the difference was not significant. The greatest root dry biomass production recorded in this study for both mycorrhizal and nonmycorrhizal maize plants was in soil with 50 kg P ha-1 maintained at ca. -0.4 MPa soil water potential. Doubling the amount of fertilizer P resulted in less root biomass production by both mycorrhizal and nonmycorrhizal plants.
In soil maintained at ca. -0.8 MPa water potential, there was no significant difference in root dry biomass production between mycorrhizal and nonmycorrhizal plants when soil was amended with 50 kg P ha-1. However, soil with 0 and 100 kg P ha-1 added P had a significant (p < 0.05) difference. The highest root dry biomass production at this soil moisture level (-0.8 MPa) was observed for both mycorrhizal (130 g plant-1) and nonmycorrhizal plants (96.1 g plant-1) in soil amended with 100 kg P ha-1, whereas the lowest root dry biomass production for both mycorrhizal and nonmycorrhizal plants was recorded under 50 kg P ha-1added P. When grown in very dry soil (-1.5 MPa), mycorrhizal plants produced significantly (p < 0.05) greater root dry biomass than nonmycorrhizal plants in soil with no added P. However, in soil with 50 and 100 kg P ha-1, no significant differences between mycorrhizal and nonmycorrhizal plants were found. In this dry soil (-1.5 MPa), doubling the amount of fertilizer P from 50 to 100 kg P ha-1 resulted in significantly less root dry biomass production by mycorrhizal plants.
Tissue P concentration
Mycorrhizal maize plants had higher tissue P concentration than non-mycorrhizal plants at all P levels except at 50 and 100 kg P ha-1 in very dry soil (-1.5 MPa). In most treatments, mycorrhizal plants had significantly greater P concentration than nonmycorrhizal plants (Figure 5). The only treatment in which nonmycorrhizal plants had significantly greater P concentration (p < 0.05) than mycorrhizal plants was the treatment with the driest soil (-0.05 MPa) and greatest added fertilizer P (100 kg P ha-1). Statistical analyses indicate P concentration in plant tissues was significantly affected by soil moisture content, P fertilization rate and AMF, with significant (p < 0.05) interactions between these three factors.
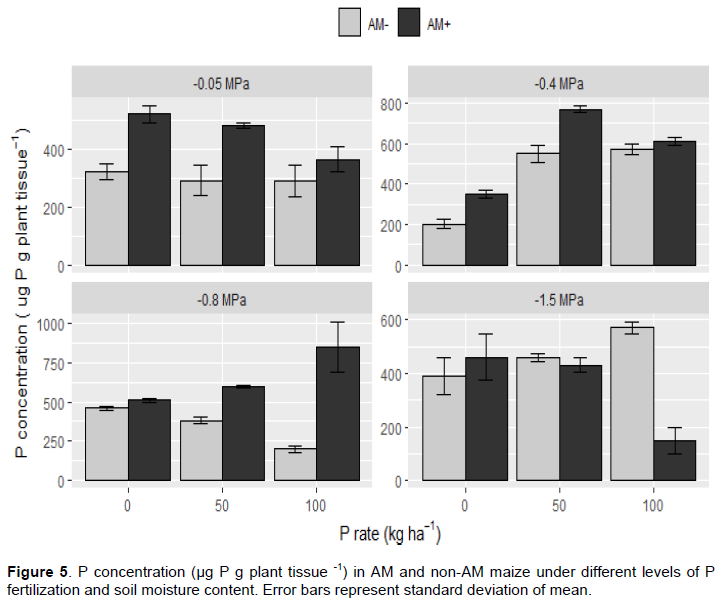
Under wet soil conditions (-0.05 MPa), P concentration in mycorrhizal plant tissues was significantly (p < 0.05) greater than nonmycorrhizal plants at 0 and 50 kg P ha-1 P fertilizer rates. However, at 100 kg P ha-1 and the same soil water potential, there was no significant difference between mycorrhizal and nonmycorrhizal plants. Tissue P concentration in mycorrhizal plants was lower when grown in soil fertilized with excess P than in soil to which no fertilizer P was added. Mycorrhizal plants grown in soil fertilized with recommended rates of P (50 kg P ha 1) had greater tissue P concentration than mycorrhizal plants grown in excess P fertilized soil, but less than mycorrhizal plants grown in soil with no added fertilizer P. Adding P fertilizer to wet soil did not result in greater tissue P concentration in either mycorrhizal or nonmycorrhizal plants. When mycorrhizal plants were grown with no added P in wet soil (-0.05 MPa), mean P concentration was 520 µg P g plant tissue-1, while under 50 and 100 kg P ha-1 P concentration were 480 and 363 µg P g plant tissue-1, respectively. For nonmycorrhizal plants, the highest tissue P concentration (320 µg P g plant tissue -1) was recorded in soil with no added P.
At the moderate water potential of -0.4 MPa, mycorrhizal plants had significantly (p < 0.05) higher tissue P concentration than nonmycorrhizal plants in soil fertilized with 0 and 50 kg P ha -1. Meanwhile, no significant difference in tissue P concentration was found between mycorrhizal and nonmycorrhizal plants in the treatment with 100 kg P ha-1. At this soil moisture level, doubling the amount of fertilizer P resulted in lower tissue P concentration in mycorrhizal plants and greater P concentration in nonmycorrhizal plants.
In soil maintained at -0.8 MPa water potential condition, no significant difference in P concentration between mycorrhizal and nonmycorrhizal plants was observed in soil with no added P, while in soil with 50 and 100 kg P ha-1 added P had significant (p < 0.05) differences. Highest mean P concentration was observed for mycorrhizal plants (847 µg P g plant tissue-1) when soil was amended with 100 kg P ha-1 and for nonmycorrhizal plants (460 µg P g plant tissue-1) when no P was added. Lowest mean P concentration was observed for mycorrhizal plants (510 µg P g plant tissue -1) when no P was added and for nonmycorrhizal plant (200 µg P g plant tissue-1) when soil was amended with 100 kg P ha-1.
When grown in dry soil (-1.5 MPa), nonmycorrhizal plants had significantly (p < 0.05) greater tissue P concentration than mycorrhizal plants in soil amended with 100 kg P ha-1, but in soil with 0 and 50 kg P ha-1 added P, there were no significant differences between mycorrhizal and nonmycorrhizal plants. The greatest P concentration in this moisture level (-1.5 MPa) was observed (570 µg P g plant tissue -1) in nonmycorrhizal plants.
DISCUSSION
Data indicate AMF symbiosis (G. fasciculatum) was more effective at increasing growth and tissue P content of maize (Zea mays L.) under a range of soil moisture conditions in a slightly alkaline (pH 7.9) agricultural soil than fertilization with triple super phosphate (46% P2O5). Results indicate a positive effect of mycorrhizal fungi on host plant biomass production and tissue P content except under the highest P fertilization rates and driest soil moisture conditions. Greater effects of G. fasciculatum on maize biomass were recorded at moderate soil water content (-0.4 and -0.8 MPa) and moderate P fertilization (Figures 3 and 4). Highest growth of AM maize was recorded under moderate soil water content (-0.4 MPa) and P fertilization levels (50 kg P ha 1). Greatest growth of nonmycorrhizal maize occurred at – 0.8 MPa and 100 kg P ha-1. Bucher (2007) demonstrated that AMF fungi enhanced plant growth by increasing P uptake in low P soil. Mycorrhizal maize produced at least as much biomass, often significantly more, under all soil moisture conditions tested.
Auge et al. (2001) found mycorrhizal maize plants and non-mycorrhizal P-fertilized treatments grown under highest and lowest soil water content had similar biomass. High mycorrhizal plant growth may also be due to enhanced nitrogen, micronutrient and water uptake (Subramanian et al., 2006; Habibzadeh et al., 2013). Improvement of biomass production in mycorrhizal plants may be associated with enhanced photosynthesis related with improved nutrition in the plants (Begum et al., 2019). Previous studies have shown that maize inoculated with arbuscular mycorrhizal fungi Glomus fasciculatum and Intraradices sp. has enhanced water use efficiency under drought conditions (Zhao et al., 2015). Mycorrhizal plants in these treatments had higher tissue P content than nonmycorrhizal plants. Enhanced photosynthesis increases carbon (C) assimilation and allocation to roots of AMF plants (Koch and Johnson, 1984). This, in turn, promotes mycorrhizal root growth with positive feedback on P uptake (Smith et al., 2011). However, in this study, high P applications (100 Kg P ha-1) suppressed root biomass production in both dry and very wet soils compared with moderate soil water levels. This result could explain the harmful effect of excess P content in soil on the AMF by affecting the hyphal development and length which is considered as an important functional part of mycorrhizal symbiosis and main source of inoculum in soil (Rubio, 2003) and eliminate mycorrhizal effect on plant growth (Smith and Read, 2008; Campos et al., 2018).
AMF are associated with proliferation of root growth (Auge et al., 1994) in maize. This is especially important for maize as it is the only grass know to produce less root biomass than shoot biomass. The mycorrhizae enhancing plant root biomass was consistent across different soil moisture contents. Mai et al. (2018) also reported soil water content had more significant effects on root and hyphae growth than did P level effects. They further found out that under well-watered soil conditions, P fertilizer increased root length, while the root length was unaffected by P fertilizer under low soil water content (Mai et al., 2018) The fact that AMF increases host plant water uptake in dry soil and improves host plant water relations during drought is well documented (Stahl et al., 1998; Campos et al., 2018).
In the present study, the lowest AMF maize P content was recorded under the driest soil (-1.5 MPa) and the highest level of P fertilization (100 kg P ha-1). The greatest P tissue content of non-mycorrhizal maize occurred at – 1.5 MPa and 100 kg P ha-1, while the lowest P tissue content was recorded at – 0.8 MPa and 100 kg P ha-1. Mycorrhizal plants generally had higher tissue P concentration than nonmycorrhizal plants, except under high-water potential and practical and excess soil phosphorus levels. The result of our study agrees with findings from numerous studies (Subramanian et al., 1997; Asmah, 1995; Bucher, 2007; Nell et al., 2010) which have demonstrated plants colonized by AMF absorb more soil P than non-mycorrhizal plants, especially under phosphorus-limited condition. In soils with low P contents, AMF improve phosphorus supply to the infected roots of host plants (Bucher, 2007). AMF consider bio fertilizers which are very useful for plant growth and soil health (Sadhana, 2014). Therefore, AMF could be considered a good alternative to replace or reduce chemical fertilizer use, because mycorrhizal application can effectively reduce the quantitative use of inorganic fertilizer input especially of phosphorus (Ortas, 2012). Mycorrhizal fungi can improve nutrient uptake by increasing the exploration of the soil pore space and contact with soil solution (Bi et al., 2003).
In drought-stressed plants (-1.5 MPa), tissue P content was consistently lower in the mycorrhizal plants than in the non-mycorrhizal under highest P fertilization rate. The negative effect of increasing soil P concentration on mycorrhiza colonization was hypothesized and demonstrated by the observation of a declining amount of AMF formation with increasing soil P concentration (Balzergue et al., 2011; Lekberg and Koide, 2005). Previous studies have shown high P supply may inhibit the exudation of strigolactones, which are plant hormones that stimulate growth of AMF fungi (Balzergue et al., 2011, 2013). Moreover, high P application led to decreased AMF biomass per plant, including biomass in root and soil by changes in characteristics of root colonization (mainly reducing arbuscular development) (Smith and Read, 2008).
These results show G. fasciculatum colonized the root of maize plants under all soil water conditions tested (Figure 1). Mycorrhizal maize colonization was lowest in the driest soil (- 1.5 MPa), but still substantial (74% of 1cm root segments examined) (Oyetunji et al., 2007; Bagheri et al., 2012). Beltrano and Ronco (2008) found that AMF colonization was more negatively impacted by highest and lowest soil water content than moderate soil water potential. The results agree with this finding, water stress reduced AMF root colonization (Osonubi, 1994; Ruiz-Lozano and Azcon, 1995; Al-karaki et al., 2004). Increasing water stress reduces AMF root colorization, and our result suggests that moderate soil water content changes exert a great effect on AMF inoculation compared to findings by Smith and Read (2008). Poor colonization of roots under soil water stress may result in reducing spore germination (Daniels and Trappe, 1980; Zang et al., 2018) or hyphal growth and survival in soil (Walker, 1999).
Novel findings of the study include the observation that at all levels of soil moisture and soil P content mycorrhizal plants produce more biomass than nonmycorrhizal plants, or in a few cases, at least as much. Also, at all levels of soil moisture and soil P content, mycorrhizal plants have higher P concentration than nonmycorrhizal plants except in the driest soil (-1.5 MPa) with highest P content (100 kg P ha-1). Another unusual observation was that mycorrhizal plants grown in wet soil (-0.05 MPa) and without P produce greater shoot dry biomass (158.1 g plant-1) in 8 weeks than mycorrhizal plants grown in soils treated with 50 (149 g plant-1) and 100 kg P ha-1 (91.3 g plant-1). In this study, at high soil moisture AMF always produce plants having higher P concentration than nonmycorrhizal plants, which indicated that G. fasciculatum is extremely efficient at P uptake from soil and transfer it to plants. Remarkably, at low soil moisture added P fertilizer did result in high tissue P concentration by nonmycorrhizal plants, indicating high levels of P uptake under these conditions without a mycorrhizal symbiont. We would have hypothesized, according the literature, that at high soil P content, nonmycorrhizal plants would produce more biomass than nonmycorrhizal plants and would have higher tissue P concentration than mycorrhizal plants (Grant et al., 2004). Our observations may be indicative of slow uptake of soil P by nonmycorrhizal plants caused by limiting amount of oxygen in very wet soil (Whitmore and Whalley, 2009). Further experimentation is required to determine the mechanisms involved.
CONCLUSION
The study demonstrates an efficacious mycorrhiza strain of G. fasciculatum can have greater positive impacts on Z. mays host plant growth and P uptake than fertilizing with phosphorus. AMF, being natural symbionts relationship, provide essential plant inorganic nutrients to host plants, thereby improving plant growth under unstressed and stressed conditions. These results indicate it may be more productive for farmers to optimize efficacious AMF formation on their corn crops than use P fertilizers. This data, as well as from other researchers, shows can inhibit AMF formation under certain conditions such as adding excess amount of P fertilizers. Additionally, activity of AMF are known to have positive impacts on soil health and quality, while use of P fertilizers is known to cause environmental problems under some circumstances.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGMENTS
This study was successful because of facilitation by Omar Almukhtar University, Soil and Water Department. We appreciate the help from Alkufar Production Project for analysis of plant soil samples and providing maize grain for this project and Mr. Addulhamed Bomatary, Ibrahim Bozaid, Fathi Altwati, Dr. Albert Adjesiwor for logistics and statistical analysis and Mr. Calvin and Mr. Michael Baidoo for editing.
REFERENCES
|
Albacete AA, Martínez-Andújar C, Pérez-Alfocea F (2014). Hormonal and metabolic regulation source-sink relations under salinity and drought: from plant survival to crop yield stability. Biotechnology Advances 32(1):12-30. |
|
|
Al-Karaki G, McMicheal B, Zak J (2004). Field response of wheat to arbuscular mycorrhizal fungi and drought stress. Mycorrhiza 14(4):263-269. |
|
|
Al-Karaki GN, Al-Raddad A (1997). Effects of arbuscular mycorrhizal fungi and drought stress on growth and nutrient uptake of two wheat genotypes differing in drought resistance. Mycorrhiza 7:83-88. |
|
|
Al-Karaki GN, Clark RB (1998). Growth, mineral acquisition, and water use by mycorrhizal wheat grown under water stress. Journal of Plant Nutrition 21(2):263-276 |
|
|
Allen MF (2007). Mycorrhizal Fungi: Highways for Water and Nutrients in Arid Soils. |
|
|
Allen MF, Boosalis MG (1982). Effects of two species of VA mycorrhizal fungi on drought tolerance of winter wheat. New Phytologist 93:67-76. |
|
|
Asmah AE (1995). Effect of phosphorus source and rate of application on VAM fungal infection and growth of maize (Zea mays L.). Mycorrhiza 5(3):223-228. |
|
|
Auge' RM, Duan X, Ebel RC, Stodola AJW (1994). Nonhydraulic signaling of soil drying in mycorrhizal maize. Planta 193:74-82. |
|
|
Auge' RM, Stodola AJW , Tims JE, Saxton AM (2001). Moisture retention properties of a mycorrhizal soil. Plant and Soil 230:87-97 |
|
|
Bagheri V, Shamshiri MH, . Shirani H, Roosta HR (2012). Nutrient Uptake and Distribution in Mycorrhizal Pistachio Seedlings under Drought Stress. Journal of Agricultural Science and Technology 14:1591-1604. |
|
|
Balzergue C, Chabauud M, Barker DG, Bécard G, Rochange SF (2013). High phosphate reduces host ability to develop arbuscular mycorrhizal symbiosis without affecting root calcium spiking responses to the fungus. Frontiers in Plant Science |
|
|
Balzergue C, Puech-Pagès V, Bécard G, Rochange SF (2011). The regulation of arbuscular mycorrhizal symbiosis by phosphate in pea involves early and systemic signaling events. Journal of Experimental Botany 62(3):1049-1060. |
|
|
Begum N, Qin C, Ahanger MA, Raza S, Khan MI, Ashraf M, Ahmed N, Zang L (2019). Role of Arbuscular Mycorrhizal Fungi in Plant Growth Regulation: Implications in Abiotic Stress Tolerance. Frontiers in Plant Science 10. |
|
|
Beltrano J, Ranco MG (2008). Improved tolerance of wheat plants (Triticum aestivum L.) to drought stress and rewatering by the arbuscular mycorrhizal fungus Glomus claroideum: effect on growth and cell membrane stability. Brazilian Journal of Plant Physiology 20(1):29-37. |
|
|
Bi LY, Li XL, Christie P (2003) . Influence of Early Stages of Arbuscular Mycorrhiza on Uptake of Zinc and Phosphorous by Red Clover from a Lowphosphorous Soil Amended with Zinc and Phosphorous. Chemosphere 50(6):831-837. |
|
|
Birhane E, Sterck F, Fetene M, Bongers F, Kuyper T (2012). Arbuscular mycorrhizal fungi enhance photosynthesis, water use efficiency, and growth of frankincense seedlings under pulsed water availability conditions. Oecologia 169(4):895-904. |
|
|
Bray EA (2004). Genes commonly regulated by water-deficit stress in Arabidopsis thaliana. Journal of Experimental Botany 55(407):2331-2341. |
|
|
Bucher M (2007). Functional biology of plant phosphate uptake at root and mycorrhizae interfaces. New Phytologist 173:11-26. |
|
|
Calvo-Polanco M, Sánchez-Castr I, Cantos M, Luis García J, Azcón R, Ruiz-Lozano JM, Beuzón C R, Aroca R (2016). Effects of different arbuscular mycorrhizal fungal backgrounds and soils on olive plants growth and water relation properties under well-watered and drought conditions. Plant, Cell and Environment 39:2498-2514. |
|
|
Campos P, Borie F, Cornejo P, López-Ráez JA, López-García A, Seguel A (2018). Phosphorus Acquisition Efficiency Related to Root Traits: Is Mycorrhizal Symbiosis a Key Factor to Wheat and Barley Cropping?. Frontiers in Plant Science 9:752. |
|
|
Cavallazzi JRP, Filho OK, Stürmer SL, Rygiewicz PT, Mendonça MM (2007). Screening and selecting arbuscular mycorrhizal fungi for inoculating micropropagated apple rootstocks in acid soils. Plant Cell, Tissue and Organ Culture 90:117-129. |
|
|
Chaves MM, Oliveira MM (2004). Mechanisms underlying plant resilience to water deficits: prospects for water-saving agriculture. Journal of Experimental Botany 55(407):2365-2384. |
|
|
Daniels BA, Trappe JM (1980). Factors affecting spore germination of the vesicular arbuscular mycorrhizal fungus, Glomus Epigaeus. Mycologia 72:457-471. |
|
|
Gee GW, Bauder JW (1986). Particle-size analysis. In: Klute, A. (Ed.), Methods of Soil Analysis, Part I, 2nd ed., Agron. Monographs, ASA and SSSA, Madison, WI 9:383-411. |
|
|
Grant C, Bittman S, Montreal1 M, Plenchette C, Morel C (2004). Soil and fertilizer phosphorus: Effects on plant P supply and mycorrhizal development. Canadian Journal of Plant Science 85(1). |
|
|
Habibzadeh Y, Pirzad A, Zardashti MR, Jalilian J, Eini O (2013). Effects of arbuscular mycorrhizal fungi on seed and protein yield under water-deficit tress in mung bean. Agronomy Journal and Enviromental Quality 105(1):79-84. |
|
|
Hesse PR (1971). Textbook of Soil Chemical Analysis. John Murray, London. |
|
|
Huang RS, Smith WK, Yost RS (1985). Influence of vesicular arbuscular mycorrhiza on growth, water relations and leaf orientation in Leucaena leucocphala (Lam.) de Wit. New Phytologist 99(2):229-243. |
|
|
Kayama M, Yamanaka T (2014). Growth characteristics of ectomycorrhizal seedlings of Quercus glauca, Quercus salicina, and Castanopsis cuspidata planted on acidic soil. Trees 28:569-583. |
|
|
Koch KE, Johnson CR (1984). Photosynthate partitioning in split root citrus seedlings with mycorrhizal and non-mycorrhizal root systems. Plant Physiology 75(1):26-30. |
|
|
Koide RT (1991). Nutrient supply, nutrient demand and plant response to mycorrhizal infection. New Phytologist 117(3):365-386. |
|
|
Koide RT, Li M (1990). On host regulation of the vesicular-arbuscular mycorrhizal symbiosis. New Phytologist 114(1):59-65. |
|
|
Lekberg Y, Koide RT (2005). Is plant performance limited by abundance of arbuscular mycorrhizal fungi? A meta-analysis of studies published between 1988 and 2003. New Phytologist 168(1):189-204. |
|
|
Li J, Meng B, Chai H, Yang X, Song W, Li S, Lu A, Zanf T, Sun W (2019). Arbuscular Mycorrhizal Fungi Alleviate Drought Stress in C3 (Leymus chinensis) and C4 (Hemarthria altissima) Grasses via Altering Antioxidant Enzyme Activities and Photosynthesis. Frontier in Plant Science 10:1-12. |
|
|
Liu W, Zheng CY, Fu ZF, Gai JP, Zhang JL, Christie P, Li XL (2014). Facilitation of seedling growth and nutrient uptake by indigenous arbuscular mycorrhizal fungi in intensive agroecosystems. Biology and Fertility of Soils 50:381-394. |
|
|
Mai W, Xue X, Feng G, Tian C (2018). Simultaneously maximizing root/ mycorrhizal growth and phosphorus uptake by cotton plants by optimizing water and phosphorus management. BMC Plant Biology 18:334. |
|
|
Manjunath A, Habte M (1991). Relationship between mycorrhizal dependency and rate variables associated with phosphorus uptake, utilization and growth. Communications Soil Science and Plant Analysis 22(13-14):1423-1437. |
|
|
Marschner H, Dell B (1994). Nutrient uptake in mycorrhizal symbiosis. Plant and Soil 159:89-102. |
|
|
Mena-Violante HG, Ocampo-Jimenez O, Dendooven L, Martinez-Soto G, Gonzalez-Castafeda J, Davies FT (2006). Arbuscular mycorrhizal fungi enhance fruit growth and quality of chile ancho Capsicum annuum L. cv San Luis plants exposed to drought. Mycorrhiza 16(4):261-267. |
|
|
Michelsen A, Rosendahl S (1990). The effect of VA mycorrhizal fungi, phosphorus and drought stress on the growth of Acacia nilotica and Leucaena leucocephala seedlings. Plant and Soil 124:7-13. |
|
|
Moradtalab N, Roghieh H, Nasser A, Tobias EH, Günter N (2019). Silicon and the association with an arbuscular-mycorrhizal fungus (Rhizophagus clarus) mitigate the adverse effects of drought stress on strawberry. Agronomy 9(1)41. |
|
|
Mumtaz MZ, Aslam M, Jamil M, Ahmad M (2014). Effect of different phosphorus levels on growth and yield of wheat under water stress conditions. Journal of Environment and Earth Science 4(19):23-30. |
|
|
Nell M, Wawrosch C, Steinkellner S, Vierheilig H, Kopp B, Lössl A (2010). Root colonization by symbiotic arbuscular mycorrhizal fungi increases sesquiterpenic acid concentrations in Valeriana officinalis L. Planta Medica 76:393-398. |
|
|
Nelsen CE, Safir GR (1982). Increased drought tolerance of mycorrhizal onion plants caused by improved P nutrition. Planta 154(5):407-413. |
|
|
Olsen SR, Sommers LE (1982). Phosphorus, P, In: A. L. et al.., eds. Methods of soil analysis. Chemical and biological methods. Part 2, 2 ed. Agron. Monogr. 9. ASA and SSSA, Madison, WI. 403-430. |
|
|
Ortas I (2012). The effect of mycorrhizal fungal inoculation on plant yield, nutrient uptake and inoculation effectiveness under long-term field conditions. Field Crops Research 125:35-48. |
|
|
Osonubi O (1994). Comparative effects of vesicular-arbuscular mycorrhizal inoculation and phosphorus fertilization on growth and phosphorus uptake of maize (Zea mays L.) and sorghum (Sorghum bicolor L.) plants under drought-stressed conditions. Biology and Fertility of Soils 18:55-59. |
|
|
Oyetunji OJ, Ekanayake IJ, Osonubi O (2007). Chlorophyll Fluorescence Analysis for Assessing Water Deficit and Arbuscular Mycorrhizal Fungi (AMF) Inoculation in Cassava (Manihot esculenta Crantz). Advances in Biological Research 1(3-4):108-117. |
|
|
Phillips JM, Hayman DS (1970). Improved procedures for clearing and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Transactions of the British Mycological Society 55(1):158-161. |
|
|
Ping XU, Lin-Zhou L, Xiao-Ying D, Jing XU, Pei-Kun J, Ren-Fang S (2014). Response of Soil Phosphorus Required for Maximum Growth of Asparagus o?cinalis L. to Inoculation of Arbuscular Mycorrhizal Fungi. Pedosphere 24(6):776-782. |
|
|
Powell CL, Bagyaraj DJ (1984). VA Mycorrhiza. In. Menge JA (Eds), Inoculum production Florida, BR:CRC Press, Inc. pp.187-199. |
|
|
R Core Team R: (2018). A language and environment for statistical computing. R Foundation for Statistical Consulting. Vienna. |
|
|
Rajan SK, Reddy BJD, Bagyaraj DJ (2000). Screening of arbuscular mycorrhizal fungi for their symbiotic efficiency with Tectona grandis. Forest Ecology and Management 126(2):91-95. |
|
|
Rubio R, Borie F, Schaalchli C, Castillo C, Azcon R (2003). Occurrence and effect of arbuscular mycorrhizal propagules in wheat as affected by the source and amount of phosphorus fertilizer and fungal inoculation. Applied Soil Ecology 23(3):245-255. |
|
|
Ruiz-Lozano JM, Azcon R (1995). Hyphal contribution to water uptake in mycorrhizal plants as affected by the fungal species and water status. Physiologia Plantarum 95(3):472-478. |
|
|
Ruiz-Lozano JM, Aroca R, Zamarreño ÁM, Molina S, Andreo-Jiménez B, Porcel R (2015). Arbuscular mycorrhizal symbiosis induces strigolactone biosynthesis under drought and improves drought tolerance in lettuce and tomato. Plant, Cell and Environment 39(2):441-452. |
|
|
Sadhana B (2014). Arbuscular mycorrhizal fungi (AMF) as a biofertilizers-a review. Intnational Journal of Current Microbiology and Applied Science 3(4):384-400. |
|
|
Schubert A, Hayman DS (1986). Plant growth responses to Vesicular-arbuscular mycorrhiza XVI. Effectiveness of different endophytes at different levels of soil phosphate. New Phytologist 130:79-90. |
|
|
Smith SE, Read DJ (2008). Mycorrhizal Symbiosis, (Ed3), Academic Press, New York. London, Burlinghton, San Diego. |
|
|
Smith SE, Jakobsen I, Grønlund M, Smith FA (2011). Roles of arbuscular mycorrhizas in plant phosphorus (P) nutrition: interactions between pathways of P uptake in arbuscular mycorrhizal (AM) roots have important implications for understanding and manipulating plant P acquisition. Plant Physiology 156:1050-1057. |
|
|
Smith SE, Smith FA, Jakobsen I (2003). Mycorrhizal fungi can dominate phosphate supply to plants irrespective of growth responses. Plant Physiology 133:16-20. |
|
|
Stahl PD, Frost SM, Williams S E, Schuman G E (1998). Arbuscular Mycorrhizae and Water Stress Tolerance of Wyoming Big Sagebrush Seedlings. Soil Science Society of America 62(5):1309-1313. |
|
|
Stahl PD, Smith WK (1984). Effects of different geographic isolates of Glomus on the water relations of Agropyron Smithii. Mycologia 76(2):261-267. |
|
|
Subramanian KS, Charest C, Dwyer LM, Hamilton RI (1997). Effects of mycorrhizas on leaf water potential, sugar and P contents during and after recovery of maize. Canadian Journal Botany 75(9):1582-1591. |
|
|
Subramanian KS, Santhanakrishnan P, Balasubramanian P (2006). Responses of field grown tomato plants to arbuscular mycorrhizal fungal colonization under varying intensities of drought stress. Scientia horticulturae, 107(3):245-253. |
|
|
Walker C (1999). Methods for culturing and isolating arbuscular mycorrhizal fungi. Mycorrhiza News 11(2). |
|
|
Walkley AJ, Black CA (1934). An extimation of the Degtjareff method for determining soil organic matter and a proposed modification of the chromic acid titration method. Soil Science 37(1):29-38. |
|
|
Walsh LM, Beaton JD (1973). Soil testing and plant analysis (Eds.). Soil Science Society of America, Madison,WI P 512. |
|
|
Whitmore AP, Whalley WR (2009). Physical effects of soil drying on roots and crop growth. Journal of Experimental Botany 6(10):2845-2857. |
|
|
Wu QS, Zou YN (2009b). Mycorrhizal influence on nutrient uptake of citrus exposed to drought stress. Philippine Agricultural Scientist 92:33-38. |
|
|
Xu L, Li T, Wu Z, Feng H, Yu M, Zang X, Chen B (2018). Arbuscular mycorrhiza enhances drought olerance of tomato plants byregulating the 14-3-3 genes in the ABA signaling pathway. Applied Soil Ecology 125:213-221. |
|
|
Yang H, Zehnder AJB (2002). Water Scarcity and Food Import: A Case Study for Southern Mediterranean Countries. World Development 30(8):1413-1430. |
|
|
Yooyongwech S, Samphumphuang T, Tisarum R, Theerawitaya C, Chaum S (2016). Arbuscular mycorrhizal fungi (AMF) improved water deficit tolerance in two different sweet potato genotypes involves osmotic adjustments via soluble sugar and free proline. Scientia Horticulturae 198:107-117. |
|
|
Zhang F, Zou YN, Wu QS (2018). Quantitative estimation of water uptake by mycorrhizal extraradical hyphae in citrus under drought stress. Scientia Horticulturae 229:132-136. |
|
|
Zhao R, Guo W, Bi N, Guo J, Wang L, Zhao J, Zhang J (2015). Arbuscular mycorrhizal fungi affect the growth, nutrient uptake and water status of maize (Zea mays L.) grown in two types of coal mine spoils under drought stress. Applied Soil Ecology 88:41-49. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


