Full Length Research Paper

ABSTRACT
The genetic structure and variability was described among 52 oil palm inter-population crosses after two selection cycles using simple sequence repeats markers (SRR). The 10 sets of SSR markers covering 62.5% of the oil palm genome showed a high level of polymorphism (average number of alleles per locus = 6.7; unbiased expected heterozygosity = 0.655; G”st = 0.346) across the oil palm samples. On the basis of allelic variability indices, three genetically diverse progenies (P1, P2 and P5) with estimates higher than the overall mean values were identified. Analysis of molecular variance revealed that partitioning of variance was higher (82%) among palms within each progeny than among the progenies (18%). High pairwise fixation index (FST > 0.150) among progenies was evident, particularly between progeny P4 and progenies P1, P2 and P3. Crossing of selected palms from highly differentiated progenies could generate offsprings with more genetic variation. The mean Nei's standard genetic distance across progenies was 0.364. The lowest genetic distance was observed between progeny P2 and P5 (0.090) and the highest was found between progeny P1 and P4 (0.653). Based on the results, there is ample variation among the inter-population progenies for maximum exploitation of heterosis and further gains in future breeding programme.
Key words: Elaeis guineensis Jacq., microsatellite markers, NIFOR, reciprocal recurrent selection.
INTRODUCTION
Indigenous to West Africa but with widest occurrence in the South-Eastern States of Nigeria, oil palm (Elaeis guineensis Jacq.) is cultivated for palm oil and palm kernel oil extracted from the mesocarp and kernel, respectively (Ataga and Van der Vossen, 2007). Palm oil is the most preferred natural oil in the diets of Nigerians both as crude red palm oil and as refined oil (olein). Modest but steady progress has been made in the genetic improvement of the crop in the past eight decades, culminating in an increased average yield from 2.5 to 5.0 metric tonnes of fresh fruit bunch ha-1year-1 and 0.5 to 1.0 tonnes oil ha-1year-1 between 1930s and 1950s to 20-25 metric tonnes of fresh fruit bunch ha-1 year-1 and 3.5 to 4.0 tonnes oil ha-1year-1 to date (Okwuagwu et al., 2005). Even with this progress, there is still a huge discriminating and competitive demand on the yield and quality of palm oil for edible and non-edible industries. With an estimated 3 million hectares of land under cultivation, Nigeria’s share of the global palm oil production is less than 2% with about 1.01 million metric tonnes of palm oil per annum (USDA, 2019). Although this level of production is below the domestic consumption of 1.34 million metric tonnes, the deficit in supply is complemented by an annual importation of about 325,000 metric tonnes. Consequently, continuous and systematic improvement for these traits of economic importance with a proportionate expansion of area under cultivation using improved genetic stock is highly desirable to meet industry demand and ensure a country-wide sufficiency of oils and fats.
The Nigerian Institute for Oil Palm Research (NIFOR) has continued to make advances in the improvement of bunch yield and oil quality of oil palm by the implementation of modified reciprocal recurrent selection (RRS) scheme. It was reported that the RRS scheme had increased oil yield by almost 18% per cycle compared to the base population (Okwuagwu et al., 2005; Durand-Gasselin et al., 2009). During each cycle of selection, the dura (D) and tenera/pisifera (T/P) base populations are kept separate and outstanding dura and tenera palms are identified through the performance of D × T and D × P progenies. These inter-population crosses of the base population are essential in the comparative trials of the breeding programme because it represents the evaluation units to identify the best crosses for prospective inter-population hybrid production. Additionally, it is one of the main sources of parents for the development of new populations exploitable in subsequent recombination cycles of the breeding programme (Falconer, 1989). However, frequent hybridization and selection among few parents in the RRS scheme tend to affect the effective population size, allele variability and genetic structure of the populations which risks long-term genetic progress in the scheme (Cao, 1995). For rapid and continued progress in oil palm breeding, the potential variability of the populations must be considered because genetic variability is required to achieve genetic gains.
Genetic variability can be estimated through the application of different molecular techniques and to date a series of different genetic markers have been explored and developed (Powell et al., 1996). Molecular genetic markers, especially Simple Sequence Repeats (SSRs) provide an important method of assessing genetic variability and have been widely employed in a variety of plant species including oil palm (Powell et al., 1996; Bakoumé, 2016). The preference for SSR markers is associated with their high information content, co- dominance, abundance in the genome, reproducibility and PCR based detection (Powell et al., 1996).
This study is a subset of a larger programme designed to describe the genetic structure and variability among NIFOR oil palm breeding populations using molecular markers. The present paper aims at estimating the available genetic variability and population structure within and between five inter-population progenies of the NIFOR oil palm main breeding programme with SSR markers. Results obtained from this study would be exploited in the selection of new parent palms for the breeding programme as well as for the production of commercial planting material.
MATERIALS AND METHODS
Fifty-two individual palms from five divergent dura x tenera inter-population crosses in the comparative trials of NIFOR oil palm main breeding programme, referred to as P1 to P5 were evaluated. The dura (female parent) population is characterized by consistent high yield of heavy bunches and good bunch composition traits while the tenera (male parent) population has a high bunch number with excellent oil-to-mesocarp ratio. The genetic backgrounds of the parent palms were dura from Ecuador, Calabar, and Ufuma, with the tenera mainly from Umuabi open pollinated genetic collections (Okwuagwu, 1985). The experimental site is located at Benin City, Nigeria; 06°312 N latitude and 05°402 E longitude and at an altitude of 149.4 m.a.s.l. The annual rainfall range from 1595 to 1958 mm and a mean annual temperature of 31.8 to 32.0°C. The number of evaluated palms in each cross ranged from 8 to 13 individuals after hybrid validation of the progenies.
Total genomic DNA was extracted from lyophilized leaf tissues from an unopened spear leaf of individual palms following the protocol described by Doyle and Doyle (1990) with minor modifications. This protocol was originally designed for relatively large-scale DNA isolation but was scaled down to suit the present study. The amount of DNA needed for the SSR reaction is very small, only about 25 ng for a single reaction of a total volume of 10 μl. The PCR conditions using M13-tailed primers described by Ting et al. (2013) was used for the amplification of 10 SSR loci/primer pairs (mEgCIR0793, sMg00156, sEg00154, sMo00102, sMg00016, mEgCIR3519, mEgCIR0790, sEg00151, sMg00179, and sMg00087) developed for oil palm (Billotte et al., 2005; Singh et al., 2008). The detection of amplification products was carried out with an automated capillary DNA genetic analyzer (ABI 3739, Applied Biosystems, USA) at the Genomics unit of Advanced Biotechnology and Breeding Centre (ABBC), Malaysian Palm Oil Board (MPOB) Malaysia. Allele calls and sizing was performed using the software GeneMapper® version 4.1 (Applied Biosystems, USA).
Marker informativeness was evaluated based on the standardized genetic differentiation measure (G”st; Hedrick 2005) after correction for small population size as described by Peakall and Smouse (2006; 2012). In addition, estimates of population genetic variation such as average number of alleles (Ao), effective number of alleles (Ae), number of private alleles (PA) - number of alleles unique to a particular population, Shannon information index (I), observed heterozygosity (Ho), unbiased expected heterozygosity (uHe), and fixation index (FST) were computed for each progeny across the 10 SSR loci. The proportion of genetic variability components within and between the inter-population progenies was determined by analysis of molecular variance (AMOVA). Pairwise (Wright, 1978) and genetic distance values among progenies based on unbiased Nei's standard genetic distance (Nei 1978) were estimated to provide information of the relatedness among the progenies. All calculations were facilitated using GenAlEx version 6.5 software (Peakall and Smouse, 2006; 2012).
RESULTS
Genetic variation is fundamental for populations to be able to face the present environmental changes and to ensure long term response to selection for traits of economic interest. In this study, we have examined levels of allelic variability and population structure within and among five inter-population progenies of oil palm, using SSR markers. Table 1 gives the synopsis of the 10 SSR markers including the basic genetic diversity parameters estimated. The marker informativeness (G”st) value for all the scored markers in this study ranged from 0.081 to 0.635. Six markers namely, mEgCIR0793, sEg00154, sMo00102, sMg00016, sEg00151 and sMg00087 revealed high G”st values (>0.250) which provided sufficient discrimination to assess genetic variation within and between the progenies. The number of different alleles across the 10 SSR markers varied from 5 (sEg00154 and sMg00016) to 9 (mEgCIR3519 and sMg00087) with a mean value of 6.7 alleles per locus (Table 1).
The observed heterozygosity (Ho) values ranged from 0.372 (mEgCIR3519) to 0.808 (sMg00156) with an overall average of 0.585. The unbiased expected heterozygosity (uHe) showed slightly higher values than Ho with values ranging from 0.482 (mEgCIR3519) to 0.825 (sMg00087) and a mean value of 0.655. The high mean uHe value suggests a high expected heterosis effect upon which oil palm yield depends. The high allelic variation and genetic diversity across the 10 SSRs were relatively higher than those reported by Putri et al. (2017; Ao = 3.0; He = 0.543) for MTG oil palm commercial variety from Socfindo, Indonesia. However, the extent of SSR diversity in the current study (He = 0.655) was lower than that reported by Augustina et al. (2013) among 85 oil palm pisifera accessions from SA Indonesia (Ao = 8.2; He = 0.70). This variability could be explained by differences in the number, origin of genotypes and the number of SSR markers used.
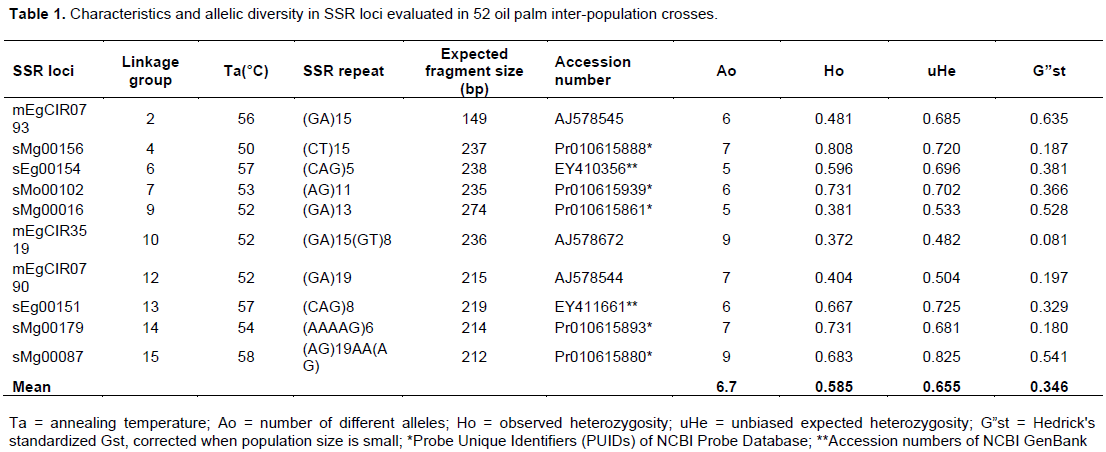
Results of the allelic patterns across the progenies revealed the presence of 19 different alleles varying from 3.2 (Progeny P4) to 4.4 (Progeny P2) with a mean value of 3.8 alleles per locus (Figure 1).
Socfindo (Putri et al., 2018). It is possible that the lower number of alleles observed among the progenies is a consequence of selection pressure on the parents of the progenies. Several studies have shown a general tendency of allele loss or reduction in the number of alleles after several cycles of selection (Cochard et al., 2009). Private alleles (PA) were identified among the progenies evaluated (Figure 1). The progenies with the most private alleles were P1 (0.6), P2 (0.5), and P3 (0.5) respectively. Progenies with private alleles can be used as sources of parents in the breeding programme to broaden the genetic base of the breeding population as well as development of more adapted variety. Progeny P4 recorded the least Shannon’s index (I = 0.894) whereas P2 revealed the highest (I = 1.105). The mean observed (Ho) and unbiased expected (uHe) heterozygosities across the progenies were 0.578 and 0.590 respectively, implying that the progenies are highly heterozygous regardless of the two generations of selection by the parents. The uHe values ranged from 0.55 (P4) to 0.61 (P1 and P2). Meanwhile, Ho was lowest for P3 (0.52) while the highest was for P2 (0.64). The uHe values obtained in these progenies were high compared to those obtained from Ulu Remis Deli (He = 0.493; Bakoumé et al., 2007) and E. oleifera (He = 0.221; Maizura et al., 2017). However, the values were lower than those reported for oil palm germplasm from Owerri (He = 0.745; Bakoumé et al., 2007). Mean F-value (inbreeding coefficient) was low and negative (-0.027), implying excess of heterozygosity due to heterotic selection in the inter-population crosses. An excess of heterozygosity suggests the impact of selection, which favours heterozygous individuals. The negative F-values for most of the populations showed the prevalence of outcrossing, whereas the positive value in P4 indicated some level of inbreeding. Considering the high Ho and uHe values in most of the populations (P1, P2 and P5), the presence of private alleles unique to a specific population may explain the excess of heterozygosity. Figure 2 represents the components of genetic variance of the oil palm inter-population progenies estimated by AMOVA.
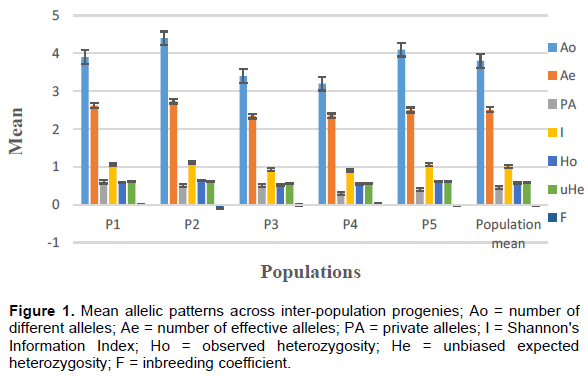
The analysis of molecular variance showed that 82% of the total genetic variations were due to differences within populations, while 18% were due to genetic variation among populations. The maximum percentage of variation (82%) present was among palms within populations, while the remaining 18% of the total variation was partitioned among populations. Permutation tests suggest that the total genetic differentiation among populations (PhiPT) was significant (0.179; p = 0.001) which indicates that differences among progenies are significant. This high level of intra-population diversity and low but significant genetic differentiation among oil palm progenies is in agreement with previous reports on oil palm (Bakoumé et al., 2007; Allou et al., 2008; Maizura et al., 2017), and consistent with outbreeding perennial species maintaining most of their variation within populations (Hamrick et al., 1992).
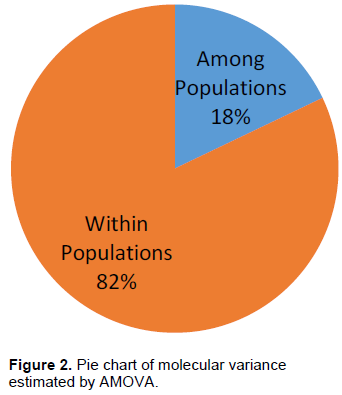
Genetic differentiation estimated by pairwise FST values among progenies were in the range of 0.029 to 0.179 (Table 2; upper diagonal). Progeny P4 was highly differentiated from P1, P2 and P3 with FST values above 0.150. According to Wright (1978), FST values between 0.15 and 0.25 indicate a high level of inter-population divergence. This high genetic relationship between individual palms of different progenies will enhance selection of parents for the establishment of crossing programmes to maximize the probability of finding transgressive hybrids in any of the following crosses; P4 x P1, P4 x P2 and P4 x P3. The unbiased Nei's standard genetic distance values ranged from 0.090 (P2 vs P5) to 0.653 (P1 vs P4) with an average distance of 0.364 (Table 2; lower diagonal). Considering the wide range of distance between the progenies and the assumption that progenies with the greatest distance is associated with inter-population heterotic potential, it may be reasonable to infer that crossing of the most distant progenies (P1 vs P4) with the other progenies will lead to the development of superior hybrids.
CONCLUSION
The results suggest that the inter-population progenies showed considerably high levels of allelic variation that will ensure continued progress in subsequent selection cycles of the breeding programme. Since high degree of genetic variation was observed among palms within the progenies, more palms should be sampled for a better insight on the genetic structure within and between the inter-population progenies. Within the framework of genetic differentiation and genetic distance of the inter-population hybridization programme, crossing of the genotype P4 with P1, P2 and P3 could offer a possible alternative for the maximum exploitation of heterosis and maintenance of genetic variation requisite for further gains in future breeding programme. The crossing between P1 and P3 may be acceptable for some gain in heterosis, although this gain will likely be lower than the crossing between P4 and P3.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGMENTS
The authors wish to express their gratitude to the members of the Genomics unit, Advanced Biotechnology and Breeding Centre (ABBC), Malaysian Palm Oil Board (MPOB) Malaysia, for their assistance with molecular analyses. They express their thanks to Ms Chizzy Colette Okoye for her assistance in collection, scoring and storage of the plant materials for DNA extraction. They also acknowledge the helpful comments made by Dr. Mehmood Hassan of ICRAF in the preliminary development of this manuscript.
REFERENCES
|
Augustina L, Rivallan R, Zulhermanna, Puspitaningrum Y, Sudarsono, Perrier X, Asmono D, Billotte N (2013). Allelic diversity of 22 Sampoerna Agro's oil palm Pisifera based on Microsatellite markers. In proceedings of the International Oil Palm Conference, Indonesia. |
|
|
Allou D, Adon B, Sangare A (2008). Molecular variability from two selection of BRT10 population in an inbreeding program of oil palm (Elaeis guineensis Jacq.) in Côte d'Ivoire. African Journal of Biotechnology 7(20):3550-3553. |
|
|
Ataga CD, Van der Vossen HAM (2007). Elaeis guineensis Jacq. In: "Plant Resources of Tropical Africa 14: Vegetable oils and fats. Submitted to PROTA, Wageningen, The Netherlands. |
|
|
Bakoumé C (2016). Genetic Diversity, Erosion, and Conservation in Oil Palm (Elaeis guineensis Jacq.). In: Ahuja MR, Mohan Jain S (eds) Genetic Diversity and Erosion in Plants: Case Histories. Springer International Publishing, Switzerland 2:1-34. |
|
|
Bakoumé C, Wickneswari R, Rajanaidu N, Kushairi A, Amblard P, Billotte N (2007). Allelic diversity of natural oil palm (Elaeis guineensis Jacq.) populations detected by microsatellite markers: implication for conservation. Plant Genetic Resource: Characterization and Utilization 5(2):104-107. |
|
|
Bakoumé C, Wickneswari R, Siju S, Rajanaidu N, Kushairi A, Billotte N (2015). Genetic diversity of the world's largest toil palm (Elaeis guineensis Jacq.) field gene bank accessions using microsatellite markers. Genetic Resource Crop Evolution 62:349- 360. |
|
|
Billotte N, Marseillac N, Risterucci AM, Adon B, Brottier P, Baurens FC, Sing R, Herrán A, Asmady, Billot C, Amblard Ph, Durand- Gasselin T, Courtois B, Asmono D, Cheah SC, Rhode W, Ritter E, Charrier A (2005). Microsatellite-based high density linkage map in oil palm (Elaeis guineensis Jacq.). Theoretical and Applied Genetics 110(4):754-765. |
|
|
Cao TV (1995). Organisation de la variabilité génétique chez le palmier à huile (Elaeis guineensis Jacq.): Conséquences pour l'amélioration des populations et la création variétale. Thèse de Doctorat, Institut National Agronomique, Paris-Grignon, France. |
|
|
Cochard B, Adon B, Rekima S, Billotte N, Desmier R, Koutou A, Nouy B, Omore A, Razak A, Glazsmann J, Noyer J (2009). Geographic and genetic structure of African oil palm diversity suggests new approaches to breeding. Tree Genetics and Genomes 5(3):493-504. |
|
|
Doyle J, Doyle L (1990). Isolation of plant DNA from fresh tissue. Focus 12:13-15. |
|
|
Durand-Gasselin T, Cochard B, Amblard P, Nouy B (2009). Exploitation de l'hétérosis dans l'amélioration génétique du palmier à huile (Elaeis guineensis Jacq.). Le sélectionneur Français. 60:91-100. |
|
|
Falconer DJ (1989). Introduction to quantitative Genetics. 3rd Edition. Longman Scientific and Technical, Harlow, UK. |
|
|
Hamrick JL, Godt MJW, Sherman-Broyles SL (1992). Factors influencing levels of genetic diversity in woody plant species. New Forests 6:95-124. |
|
|
Hedrick PW (2005). A standardized genetic differentiation measure. Evolution 59(8):1633-1638. |
|
|
Maizura I, Chee-Keng T, Wickneswari R (2017). Genetic diversity of Elaeis oleifera (HBK) Cortes populations using cross species SSRs: implication's for germplasm utilization and conservation. BMC Genetics 18:37. |
|
|
Nei M (1978). Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 89:583-590. |
|
|
Okwuagwu CO (1985). The Genetic Basis of the NIFOR Oil Palm Breeding Programme. In Proceedings: International Workshop on Oil Palm Germplasm and Utilization. Palm Oil Research Institute Malaysia 10:228-237. |
|
|
Okwuagwu CO, Ataga CD, Okolo EC, Ikuenobe CE, Ugbah MM (2005). The production of NIFOR Elite Tenera hybrid planting material - The NIFOR EWS. Technical Report, Nigerian Institute for Oil Palm Research (NIFOR), Benin City, Nigeria. |
|
|
Peakall R, Smouse PE (2006). GENALEX 6: genetic analysis in Excel. Population genetic software for teaching and research. Molecular Ecology Notes 6:288-295. |
|
|
Peakall R, Smouse PE (2012). GenAlEx 6.5: genetic analysis in Excel. Population genetic software for teaching and research - an update. Bioinformatics 28:2537-2539. |
|
|
Putri LAP, Setiado H, Hardianti R (2017). DNA Profiles of MTG (Moderat Tahan Gano) Oil Palm Variety Based on SSR Markers Indonesian Oil Palm Conference Series: Materials Science and Engineering 180 0120144. |
|
|
Putri LAP, Basyuni M, Bayu ES, Arvita D, Arifiyanto D, Syahputra I (2018). Identification of molecular performance from oil palm clones based on SSR markers Indonesian Oil Palm Conference Series: Earth and Environmental Science 126 012149. |
|
|
Powell W, Morgante M, Andre C, Hanafey M, Vogel J, Tingey S, Rafalski A (1996). The comparison of RFLP, RAPD, AFLP and SSR (microsatellite) markers for germplasm analysis. Molecular Breeding 2:225-238. |
|
|
Singh R, Zaki NM, Ting NC, Rosli R, Tan SG, Low ETL, Ithnin M, Cheah SC (2008). Exploiting an oil palm EST the development of gene-derived and their exploitation for assessment of genetic diversity. Biologia 63:1-9. |
|
|
Ting NC, Jansen J, Nagappan J, Ishak Z, Chin CW, Tan SG, Cheah SC, Singh R (2013). Identification of QTLs associated with callogenesis and embryogenesis in oil palm using genetic linkage maps improved with SSR markers. PLoS ONE 8(1):e53076. |
|
|
The United States Department of Agriculture (USDA) (2019). Market Uncertainties of 2018/19 Haunt 2019/20 Prospects. |
|
|
Wright S (1978). Variability within and among natural populations. Vol. 4. The University of Chicago Press, Chicago. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


