Full Length Research Paper

ABSTRACT
Sorghum is an important staple food crop in Kenya, but its yield is low due to many constraints which include diseases, drought, soil fertility, and pests, notably parasitic weed Striga hermonthica. Twenty elite sorghum genotypes and one resistant check were evaluated in three replications in a complete randomized design in Agar Gel (AG) experiment and randomized complete block design on selected Striga hotspot farms at Kadel and Nyahera in western Kenya. Significant differences (p<0.001) were realized both in the fields (plant height, dry shoot biomass yield, grain yield, field Striga count, and Striga damage ratings) and in the AG experiment with respect to maximum germination distance and numbers of induced germinated Striga seeds. Among the sorghum genotypes evaluated under field conditions, T53B, N57, N68, C26, IESV 92036-SH, T30B, and Uyoma 47 White genotypes had stable yields under Striga infestation in the two sites, whereas Nyadundo 1, Nyadundo 2, and Uyoma 8 were susceptible. In the AG experiment, E117B, T30B, Uyoma 8, Uyoma 42 STR, and T53B induced high numbers of germinated Striga seeds with maximum germination distances (MGD). In contrast, Uyoma 47 Brown, IESV 92038/2-SH, and IESV 92036-SH sorghum genotypes secreted low amounts of SLS suggesting resistance. This study has identified and selected four Striga tolerant (T53B, N68, N57, and T 30B) and resistant (C 26, Uyoma 47 Brown, IESV 92038/2-SH, and IESV 92036-SH) sorghum genotypes which can be utilized in further research programs or adopted to improve sorghum productivity in the highly Striga infested regions of western Kenya.
Key words: Striga, sorghum genotypes, resistant, susceptible, tolerant, Agar Gel experiment
INTRODUCTION
Sorghum (Sorghum bicolor (L.) Moench) is the fifth most important staple food crop that can meet the increased food needs of more than 500 million people in marginal regions that still experience periodic food deficits (Aragaw et al., 2021). Its drought tolerance and ability to withstand more of other biotic and abiotic stresses compared to maize has made it an important food security crop in most regions of sub-Saharan Africa (Hadebe et al., 2017).
In Kenya, it is grown mainly under unreliable rain-fed agriculture in Striga stricken agricultural regions of Eastern and western Kenya (Muui et al., 2019).![]() The area under subsistence cultivation of sorghum in Kenya has increased from 240,403 ha in 2018 to 301,705 ha in 2019 (FAOSTAT 2019), this owes to the resilience of the crop and frequent maize failures experienced especially in the drought-prone agricultural regions of Kenya (Okeyo et al., 2020). Despite this, the national yield average of sorghum has remained low at 0.8 tons per hectare (Okeyo et al., 2020) attributed to a number of abiotic and biotic constraints, especially drought and increased susceptibility of the cultivars to Striga weed. This follows despite sustained research intervention measures such as breeding for better yields and distribution of high-yielding and stress-tolerant sorghum varieties (Atera and Onyango, 2011).Striga hermonthica is the largest biological constraint to sorghum production in Sub-Saharan Africa (Yohannes et al., 2017). The weed is invasive and has rapidly invaded new geographical regions that were initially free from Striga (Teka, 2014). Studies have shown that Striga can reduce host grain yield to almost zero in high field infestation levels and as a result, many farmers have abandoned their fields that are no longer productive (Shaibu et al., 2021). In Sub Saharan Africa, yield losses due to Striga in sorghum and millet ranges between 20 to 95 and are estimated at approximately US $ 7 billion annually (Joel and Muchugi, 2018) In Ethiopia and Sudan, grain yield losses of between 65 to 100% have been reported from heavily infested fields (Bejiga, 2019). In Kenya, about 80,000 hectares cropped to Maize, Sorghum, and Millet are severely infested causing an estimated respective average grain yield losses of 1.15, 1.10, and 0.99 tons per hectare (Makete et al., 2017). The observed Striga-related high yield losses have occurred mainly due to the wide adoption of cultural methods of Striga control such as crop rotation of infested land with non-susceptible crops, prolonged fallowing, sanitation, and hand weeding of mature Striga plants particularly in subsistence farming. The practical limitations of these techniques are the more than 3 years required for rotation together with the tedious, and time-consuming nature of hand weeding of mature Striga plants that have rendered these Striga control methods impractical and less effective in controlling Striga (Lee and Thierfelder, 2017). Besides, the recommended chemical control methods such as the application of ethylene for suicidal Striga seed germination and use of nitrogen fertilizers go beyond the reach of small-scale farmers and are not feasible due to high costs (Abbas et al., 2018). Biological methods of control on the other hand are less available to farmers due to the required implementation technical skills (Bo et al., 2020). Therefore, developing Striga-resistant sorghum genotypes could provide a sustainable long-term solution to the Striga problem especially to the resource-poor farmers since its adoption requires no additional input (Badu-Apraku and Fakorede, 2017). Striga damages its host in several ways which include withdrawal of hosts (photosynthate, water, mineral nutrients, and amino acids), allelopathy, hormonal imbalance, and reduction of the host’s photosynthetic parameters. These effects lead to phenotypically reduced heights, leaf necrosis, and relatively small panicle sizes in the infested host (Beyene and Egigu, 2020). The extent of host agronomic damage is determined mainly by its resistance, tolerance, or susceptibility to Striga. The genotypes which support the growth of few Striga plants and suffer minimal agronomic damage are considered to be resistant while those that are tolerant can produce equally good yield even when under severe field Striga infestation (Mbuvi et al., 2017). The susceptible genotypes, on the other hand, incur high yield depression due to reduced panicle sizes and often fail to reach the reproductive stage either when under severe or low Striga infestation (Dafaallah and Babiker, 2019). These observations encourages breeders to develop more resistant/tolerant sorghum cultivars to minimize Striga-related high yield losses since field observations and surveys still indicate that some genotypes grown locally by farmers are tolerant to Striga and can produce good yield even when highly infested with Striga (Joel et al., 2018).
The area under subsistence cultivation of sorghum in Kenya has increased from 240,403 ha in 2018 to 301,705 ha in 2019 (FAOSTAT 2019), this owes to the resilience of the crop and frequent maize failures experienced especially in the drought-prone agricultural regions of Kenya (Okeyo et al., 2020). Despite this, the national yield average of sorghum has remained low at 0.8 tons per hectare (Okeyo et al., 2020) attributed to a number of abiotic and biotic constraints, especially drought and increased susceptibility of the cultivars to Striga weed. This follows despite sustained research intervention measures such as breeding for better yields and distribution of high-yielding and stress-tolerant sorghum varieties (Atera and Onyango, 2011).Striga hermonthica is the largest biological constraint to sorghum production in Sub-Saharan Africa (Yohannes et al., 2017). The weed is invasive and has rapidly invaded new geographical regions that were initially free from Striga (Teka, 2014). Studies have shown that Striga can reduce host grain yield to almost zero in high field infestation levels and as a result, many farmers have abandoned their fields that are no longer productive (Shaibu et al., 2021). In Sub Saharan Africa, yield losses due to Striga in sorghum and millet ranges between 20 to 95 and are estimated at approximately US $ 7 billion annually (Joel and Muchugi, 2018) In Ethiopia and Sudan, grain yield losses of between 65 to 100% have been reported from heavily infested fields (Bejiga, 2019). In Kenya, about 80,000 hectares cropped to Maize, Sorghum, and Millet are severely infested causing an estimated respective average grain yield losses of 1.15, 1.10, and 0.99 tons per hectare (Makete et al., 2017). The observed Striga-related high yield losses have occurred mainly due to the wide adoption of cultural methods of Striga control such as crop rotation of infested land with non-susceptible crops, prolonged fallowing, sanitation, and hand weeding of mature Striga plants particularly in subsistence farming. The practical limitations of these techniques are the more than 3 years required for rotation together with the tedious, and time-consuming nature of hand weeding of mature Striga plants that have rendered these Striga control methods impractical and less effective in controlling Striga (Lee and Thierfelder, 2017). Besides, the recommended chemical control methods such as the application of ethylene for suicidal Striga seed germination and use of nitrogen fertilizers go beyond the reach of small-scale farmers and are not feasible due to high costs (Abbas et al., 2018). Biological methods of control on the other hand are less available to farmers due to the required implementation technical skills (Bo et al., 2020). Therefore, developing Striga-resistant sorghum genotypes could provide a sustainable long-term solution to the Striga problem especially to the resource-poor farmers since its adoption requires no additional input (Badu-Apraku and Fakorede, 2017). Striga damages its host in several ways which include withdrawal of hosts (photosynthate, water, mineral nutrients, and amino acids), allelopathy, hormonal imbalance, and reduction of the host’s photosynthetic parameters. These effects lead to phenotypically reduced heights, leaf necrosis, and relatively small panicle sizes in the infested host (Beyene and Egigu, 2020). The extent of host agronomic damage is determined mainly by its resistance, tolerance, or susceptibility to Striga. The genotypes which support the growth of few Striga plants and suffer minimal agronomic damage are considered to be resistant while those that are tolerant can produce equally good yield even when under severe field Striga infestation (Mbuvi et al., 2017). The susceptible genotypes, on the other hand, incur high yield depression due to reduced panicle sizes and often fail to reach the reproductive stage either when under severe or low Striga infestation (Dafaallah and Babiker, 2019). These observations encourages breeders to develop more resistant/tolerant sorghum cultivars to minimize Striga-related high yield losses since field observations and surveys still indicate that some genotypes grown locally by farmers are tolerant to Striga and can produce good yield even when highly infested with Striga (Joel et al., 2018).
Strigolactones (SLs) exuded by sorghum roots and other Striga host plants is critical for both host’s root symbiotic colonization by arbuscular mycorrhizal (AM) fungi and induction of germination of conditioned soil Striga seeds (Waters et al., 2017; Mohemed et al., 2016). The genotypes, which secretes low amounts of SLs stimulates the germination of few conditioned soil Striga seeds and consequently support the growth of few Striga plants in field trials (Gobena et al., 2017). Conversely, all high producers of SLs lead to severe field Striga infestation often associated with susceptibility of the genotypes (Mohemed et al., 2018). Only a few studies have developed sorghum genotypes that are Striga resistant based on SLs secretion, thus the need to investigate a billion folds variations in the amount of SLs production among the cultivated and newly released lines/cultivars (Waters et al., 2017). Therefore, characterization of cultivars for SLs production together with field evaluation is a sure way of identifying resistant/tolerant genotypes, the genotypes that could be integrated to improve Striga management among small-scale sorghum farmers. The objectives of the study were, therefore, to assess performance of elite sorghum genotypes in yield and yield components under field Striga hermonthica infestation and to evaluate the genotypes’ variations in SLs production determined by maximum germination distances and numbers of induced germinated Striga seeds in agar gel assays under laboratory conditions.
MATERIALS AND METHODS
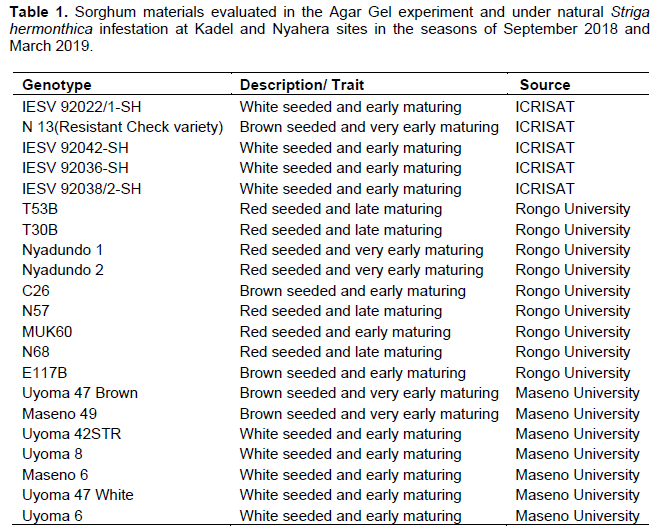
Twenty-one (21) elite sorghum materials (Table 1) from Maseno University, Rongo University, and International Centre for Research in Semi-Arid Tropics (ICRISAT) breeding programs were used in the study. Some of the materials have been released as varieties while others are elite materials yet to be released.
Experimental site description
Field trials for Striga resistance were respectively conducted in Striga hot spots at Nyahera and Kadel in Kisumu and Homabay counties. Nyahera lies at a longitude of 34°53.452′E, latitude of 0°35.977′N, and an altitude of 1490 m with an average annual rainfall of 1650 mm per annum. The soils are classified as ferralic with a pH of 5.9 (Mbogo et al., 2016). Kadel is found at a longitude of 34°24°E, latitude N-0°11’27” S, and an altitude of 1400 m above sea level with an average annual rainfall of 1200 mm (Ogenga et al., 2018). The soils are vertisol with a pH of 7.3 and made of fine loose textured clay soils. The two sites have a bimodal rainfall pattern where the first peak falls between March and June (Long rains season) and the second peak between September and December (short rains season).
Experimental design and field layout
Field trials for Striga resistance in the sorghum genotypes were laid out in a randomized complete block design (RCBD) replicated three times with two treatments, Striga plots, and Striga free plots. Evaluation of genotypes at Nyahera site was carried out under natural Striga infestation in the short rains of September 2018 and both under Striga free and Striga infestation in the long rains of March 2019. Nyahera is a site that had been used for Striga research by various organizations such as Nairobi University and International Maize and Wheat Improvement Center (Mbogo et al., 2016), the site, therefore, had already established Striga plots and Striga free plots. At Kadel, successful evaluation of sorghum genotypes was carried out only in the long rains of March 2019 under natural Striga infestation, given that the whole site was Striga infested. Land preparation was done using a disc plough and harrowed twice before planting. Planting in each plot was made in four-rows measuring 3 m in length with a row spacing of 0.75m and plant spacing of 0.2 m. Hand hoe weeding was carried out before Striga emergence and thereafter weeds other than Striga were regularly handpicked. In Striga free plots, hand hoe weeding was practiced throughout the trial as opposed to hand weeding.
Data collection
In Striga infested plots, emerged Striga counts and host damage ratings were recorded per genotype in the net plots (two central rows) at intervals of 8, 10, and 12 weeks after crop emergence. Striga damage ratings were scored using a scale of 1-5 as described by Haussmann et al. (2000b) where 1=no damage, indicating normal plant growth and high level of tolerance, and 5 representing complete collapse or death of highly susceptible sorghum genotypes. At maturity, the data was collected on dry shoot biomass yield (Kg) and grain yield (t/ha) in both Striga plots and Striga free plots.
Genotypic variation in SLs secretion among the sorghum genotypes
The Agar Gel experiment was carried out in Maseno University botanical laboratory using Striga seeds obtained from ICRISAT and sorghum genotypes listed in Table 1. Maseno University is found along the equator at latitude of 00, latitude 34° 30’E, and an altitude of 1515 m above sea level.
Surface sterilization of sorghum seeds
Sorghum seeds were surface sterilized for 5 min in 10 ml of 1% sodium hypochlorite solution, rinsed three times with distilled water, and incubated in petri dishes on moist filter papers at 28°C for 24 h. Vigorously and uniformly germinating seeds were selected and used in the agar gel assay.
Surface sterilization, viability test, and conditioning of the Striga seeds
0.5 mg of dry Striga seeds were surface sterilized in a 10 mL of 1% sodium hypochlorite solution for 5 min in a 50-mL flask containing 3 drops of Tween 20. Floating seeds and debris were discarded and the remaining seeds rinsed thoroughly with 200 ml of sterile distilled water. The Striga seeds were spread on a glass fiber filter paper (Whatman GFA) in sterile petri dishes wetted with 5 ml of sterile distilled water. The petri dishes were wrapped with aluminum foil and incubated for 8 days at 30°C. To test for seed viability the procedure described by Gbèhounou et al. (1996) was used. Briefly, 1.5 ml of a 1% tetrazolium salt solution was added to the Striga seeds after 8 days of incubation and the viable pink stained Striga seeds were counted under a microscope. The viable seeds were then conditioned for 14 days at 30°C following the procedure described by Mohamed et al. (2010).
The Agar Gel Technique (AGT) Assay
The experiment was setup in a completely randomized design (CRD) replicated three times. Approximately 1500 conditioned Striga seeds (4 drops of settled seeds) were pipetted into sterile 9 cm diameter Petri dishes. A 0.7% water agar solution (1.05 g bacto agar in 150 ml H2O) that had been autoclaved for 15 min and cooled for at least one hour was poured before it solidified into petri dishes containing an even distribution of conditioned Striga seeds. The radicles of the pre-germinated sorghum seeds were submerged in the center of the solidifying agar near one edge of the plates with the root tips pointing across the plate. The dishes were covered and placed in an incubator at 28°C for five days. The germinated Striga seeds were observed through the bottom of the petri dishes using a dissecting microscope and positions of germinated Striga seeds marked and the distances in cm between the sorghum roots and the furthest observed germinated Striga seeds measured and recorded for each sorghum genotype. The procedure described by Mohamed et al. (2010) was used.
Data analysis
The variance analysis (ANOVA) and mean grain yield, dry shoot biomass yield, Striga count, Striga damage ratings, and numbers of induced and maximum germination distances in the AG experiment were analyzed using Statistical Analysis System (SAS) software package (release 6.1), tested for significance at 5% level and means separated using 5% least significant difference (L.S.D). Actual field Striga counts were transformed using the formula log (X + 1), where X is the original field Striga counts. Relative yield loss, which is the differences between genotypes’ realized grain yields in Striga free plots and Striga plots, was calculated using the formula developed by Rodenburg et al. (2006) shown below:
RYL= [(Yc – Ys) / Yc] x 100
Where Yc is the average yield of Striga-free plots of a specific sorghum genotype and Ys is the observed yield of the same sorghum genotype grown under Striga infestation.
RESULTS AND DISCUSSION
Variations in agronomic performance among sorghum genotypes under field Striga hermonthica infestation
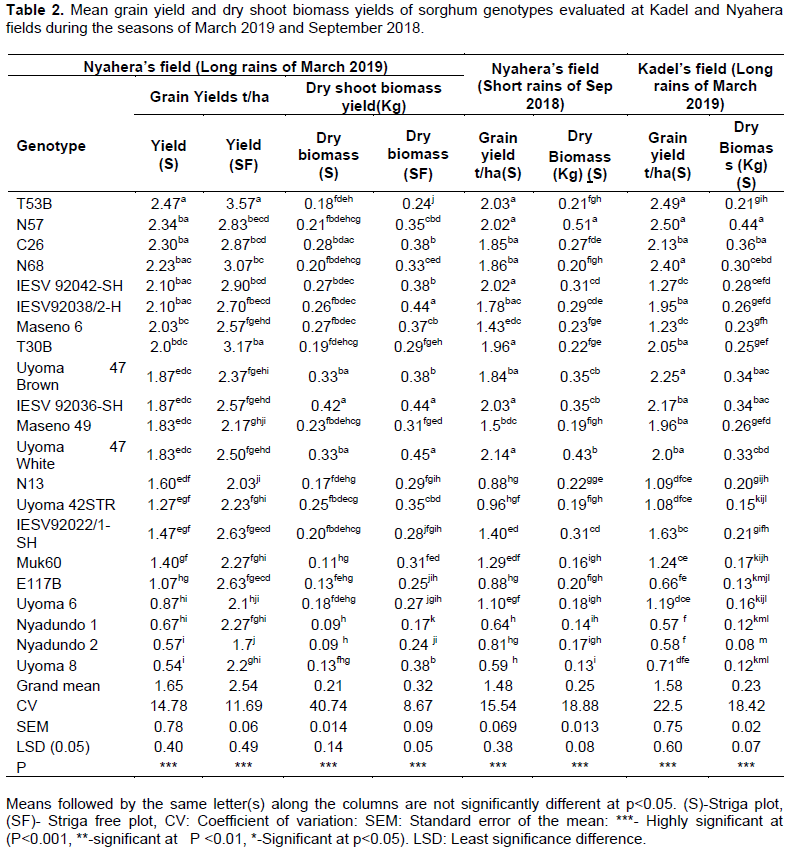
The elite sorghum genotypes differed significantly (p<0.001) in grain yield and dry shoot biomass yield under field conditions (Table 2). In the first season of Sep 2018 of field evaluation of genotypes under Striga infestation at Nyahera, Uyoma 47 White (2.14 t/ha) and T53B (2.03 t/ha) sorghum genotypes recorded the highest mean grain yields, while Uyoma 8 (0.59 t/ha), Nyadundo 1 (0.64 t/ha), and Nyadundo 2 (0.81 t/ha) genotypes recorded the lowest grain means. In the second season of March 2019 at Nyahera, T53B genotype recorded the highest mean grain yield of 2.47 t/ha while Uyoma 8 recorded the lowest yield mean of 0.54 t/ha under Striga infestation. In Striga free plots at Nyahera in March 2019 season, the highest and lowest yields of 3.57 and 1.7 t/ha were respectively recorded in T53B and Nyadundo 2 sorghum genotypes. In terms of dry biomass yields, IESV 92036-SH, Uyoma 47 White, and Uyoma 47 Brown sorghum genotypes consistently recorded high biomass means both in the Striga infested plots and in Striga free plots while Nyadundo 1 and Nyadundo 2 recorded low yield means. In March 2019 at field evaluation of genotypes for Striga resistance at Kadel, N57, T53 B, N68, and Uyoma 47 Brown sorghum genotypes exhibited high yields of between 2.25 and 2.50 t/ha while Nyadundo 1 (0.57), Nyadundo 2 (0.58 t/ha), E117B (0.66 t/ha) and Uyoma 8 (0.71 t/ha) yielded low (Table 2). Equally, N57 (0.44 kg) and Uyoma 47 Brown (0.34 Kg) in addition to C26 (0.36 Kg) and IESV 92036-SH (0.34 Kg) sorghum genotypes had high dry-shoot biomass yields, while Nyadundo 1 and Nyadundo 2 genotypes recorded low respective yields of 0.12 Kg and 0.08 Kg. In totality, T53B, N 57, N 68, and T 30B genotypes had stable and high grain yield means ranging between 1.9-2.5 t/ha (Table 2) under high Striga hermonthica infestation both at Kadel and Nyahera. C 26, IESV 92036-SH and Uyoma 47 white were also some of the genotypes with good yields above 1.85 t/ha under relatively low Striga infestation, while Uyoma 8, Nyadundo 1, and Nyadundo 2 had poor yields less than 1 t/ha in the two sites.
The differences in the agronomic performances of sorghum genotypes under Striga infestation indicated the existence of a large genetic variation among the genotypes. This was expected since the evaluated genotypes were of diverse origin. The observed variations in the grain and dry shoot biomass yields under Striga infestation imply differences in the genotypes’ genetic potential and probable differences in their Striga tolerance/resistance mechanism. Although T53B, N57, N68, and T30B sorghum genotypes experienced high Striga infestation levels, the genotypes still recorded high yields and had low Striga damage scores, and, therefore, were considered as tolerant materials. C26, IESV 92038/2-SH, IESV 92036-SH, and Uyoma 47 Brown had good grain yields but with comparatively low Striga counts and were classified as resistant. However, Nyadundo 1, Nyadundo 2, and Uyoma 8 sorghum genotypes were regarded as susceptible after they recorded very low grain yields under high Striga infestation.
These findings agree with those of Van et al. (2000) in sorghum and Menkir et al. (2012) on maize hybrids who together observed that both the tolerant and resistant genotypes displayed fewer disease symptoms, had low Striga damage ratings and produced relatively higher yields than their susceptible counterparts produce. Akaogu et al. (2013) also reported similar results after evaluating the performance of maize inbreds and hybrid under Striga-infested and Striga-free environments. The low grain and dry shoot biomass yields especially in the susceptible sorghum genotypes could have resulted due to reduced photosynthetic process together with the withdrawal of photosynthates in the host crop as a result of Striga infestation. According to Frost et al. (1997) who investigated the effect of Striga hermonthica on the rate of host’s photosynthesis, about 80 to 85% of growth loss occur in Striga infested susceptible host due to a reduction in the host’s photosynthetic capacity to less than half of that in healthy hosts.
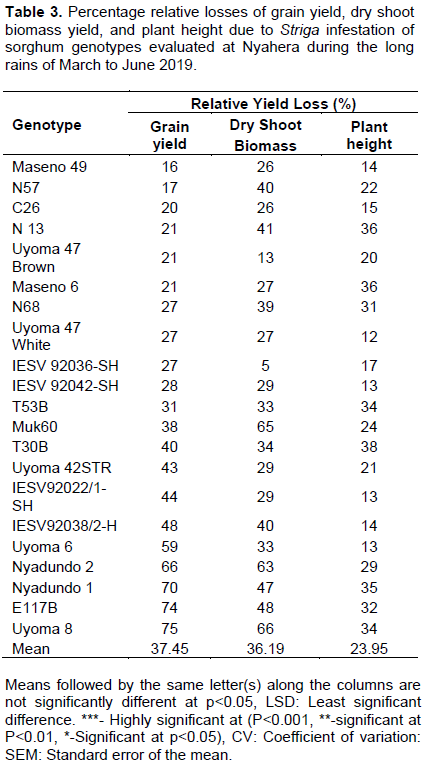
The relative grain yield losses ranged from 16 to 75% (Table 3). Uyoma 8 and E117B sorghum genotypes had the highest relative grain yield losses of 75 and 74% respectively while low scores were recorded in Maseno 49 (16%), N57 (17%), C26 (20%), Uyoma 47 Brown (21%), and N13 (21%) genotypes. Reductions in plant dry shoot biomass yields also occurred in the genotypes due to Striga infestation. The highest reductions were recorded in Muk 60 (65%), Nyadundo 2 (63%), and Uyoma 8 (66%) genotypes while the lowest reductions of 5 and 13% were respectively obtained in IESV 92036-SH and Uyoma 47 Brown sorghum genotypes (Table 3). For plant height, the highest and lowest reduction of 38 and 12% were respectively recorded in T30B and Uyoma 47 White sorghum genotypes. The high relative grain yield losses in the susceptible genotypes observed in this study conform to the findings of Mrema et al. (2017) who investigated production constraints of sorghum and Striga control practices and reported high yield losses in the susceptible sorghum genotypes infested with Striga. The findings of this study are also comparable to previous studies in maize by Badu-Apraku and Akinwale (2011) and sorghum by Abate et al. (2014) who reported significant reductions in plant height, ear height, and smaller panicle sizes in the genotypes evaluated in Striga plots than in Striga free plots.
Variations in the sorghum genotypes’ and fields levels of Striga infestation
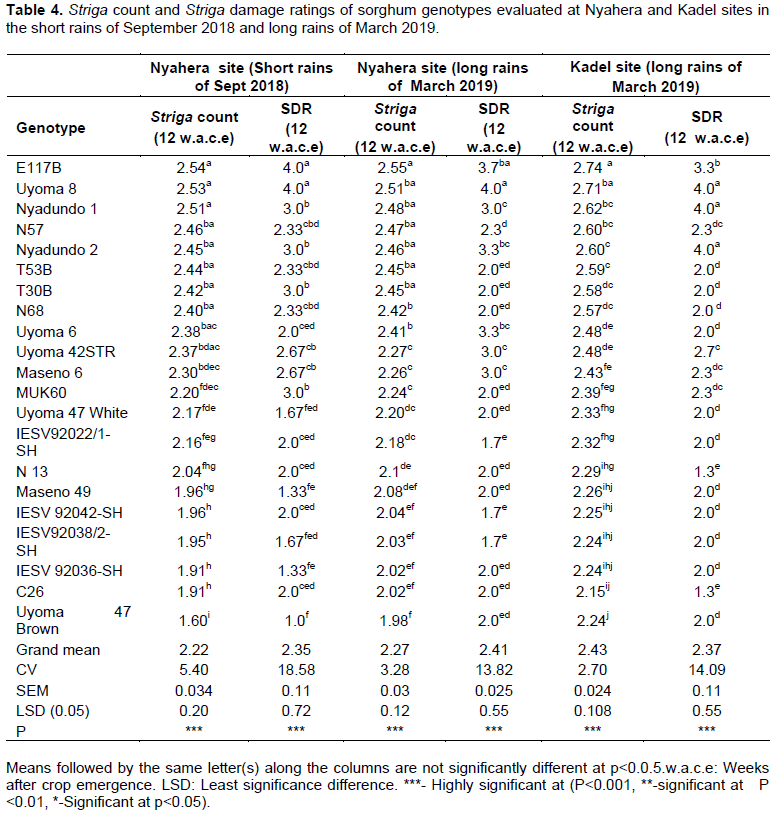
Significant genotypic variations (p<0.001) in the numbers of emerged Striga plants and Striga damage ratings were observed among the sorghum genotypes at 12 weeks after crop emergence (Table 4). At Nyahera site during the short rains of September 2018, C26, IESV 92036-SH, and Uyoma 47 Brown genotypes had the lowest mean emerged Striga counts and Striga damage ratings while E117B, Uyoma 8, and Nyadundo 1 genotypes had the highest mean emerged Striga counts and Striga damage ratings. In March 2019 in the same field, E117B, Uyoma 8, Nyadundo 1, N57, Nyadundo 2, T53B, T30B, N68, and Uyoma 6 supported the growth of many Striga plants that ranged between 2.41-2.55 per plot compared to Striga counts of 1.98 in Uyoma 47 Brown and 2.02 for both C26 and IESV 92036-SH. In terms of Striga damage ratings (SDR), the most affected sorghum genotypes under Striga infestation were Uyoma 8 and E 117 B with mean SDR scores of 4.0 and 3.7 respectively, while the least affected genotypes were IESV 92042-SH, IESV 92022/1-SH, and IESV 92038/2-SH, all with a low SDR score of 1.7. The Striga counts and Striga damage ratings followed a similar trend at field evaluation of genotypes for Striga resistance at Kadel in March 2019.
The variations in the genotypes’ field levels of Striga infestation could have been due to their differences in the amount of secreted strigolactone. The productions of root exudates and Striga seed germination have been found to correlate positively (Mohemed et al., 2016). The consistently reduced or high numbers of Striga plants supported by the evaluated sorghum genotypes under natural field Striga infestations in the two sites suggested similarity in response to Striga infestation in the two sites.
Kadel site had a higher and significantly different (p<0.001) mean Striga count of 2.43 compared to 2.27 realized at Nyahera site (Table 4). The observed difference in Striga infestation levels between the two sites could have resulted probably due to a higher pre- existing soil Striga seed bank at Kadel. This is because field Striga infestation levels have been found to strongly and positively correlate with soil Striga seed bank (Samejima et al. 2016). Moreover, previous studies have also indicated that host crops may increase the amount of secreted strigolactone in response to moisture stress (Bahadur et al., 2019; Vurro et al., 2019) resulting in high Striga infestation levels evident at Kadel field (Ogenga et al., 2018).
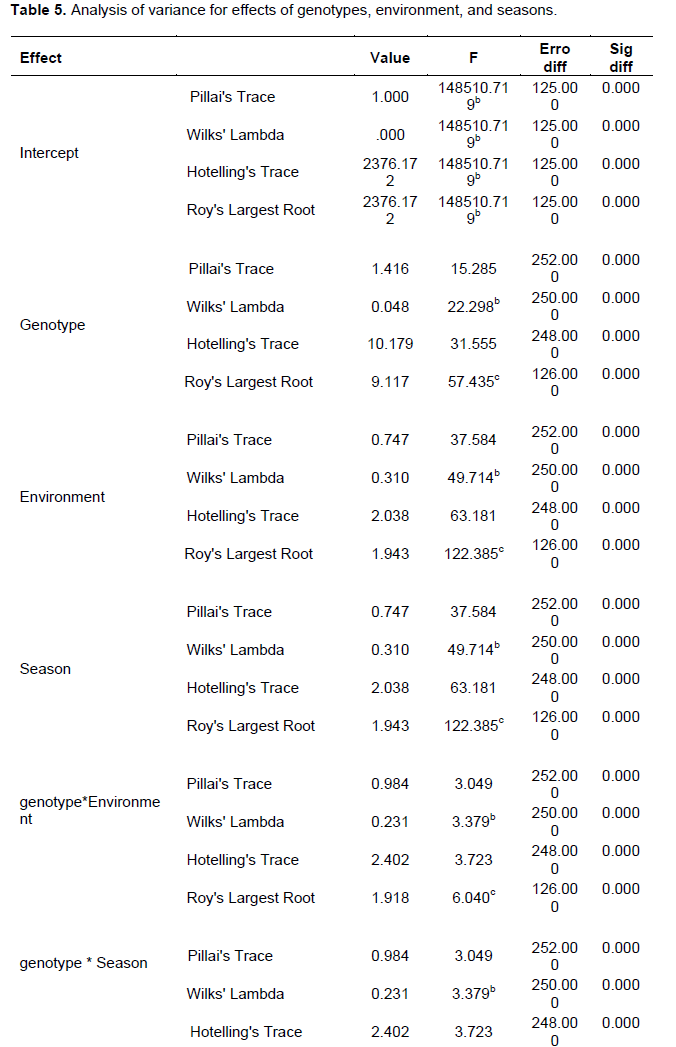
The significant total genotype × environment (G × E) variance (Table 5) that contains interaction effects between the genotypes and specific climatic and edaphic factors such as soil organic content, soil microorganisms, rainfall, and temperature that probably differed between Kadel and Nyahera could have also contributed to the observed differences in the sites’ field numbers of emerged Striga plants. This finding conforms with the findings of Haussmann et al. (2001) who investigated genotype × environment interaction patterns for Striga resistance in sorghum in Kenya and Mali and reported sites’ differences in levels of Striga infestations to have been caused by genotypes’ different response to climatic conditions. Interaction effects between (Genotype*Season), and (Season*Environment) were also found to be significant (Table 5) and therefore, combined site and seasonal analysis of Striga count, Striga damage ratings, and agronomic performance of the genotypes were not performed.
Induced and maximum distances of germinated Striga seeds in agar gel experiment
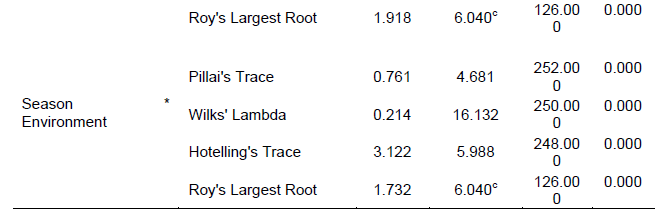
The genotypes showed significant differences (p<001) both in the numbers of induced germinated and distances of the furthest germinated Striga seeds (Table 6).
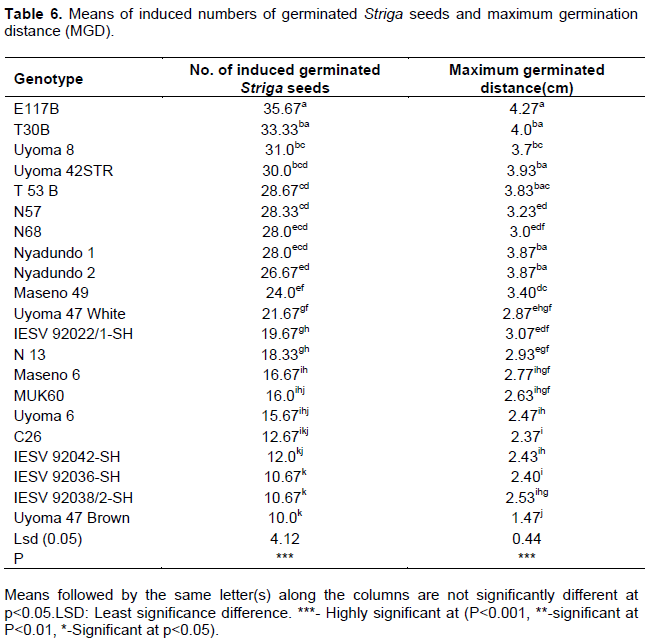
Cultivars E117B, T30B, Uyoma 8, and Uyoma 42STR induced more than 30 mean counts of in vitro germinated Striga seeds. The same genotypes also had the highest mean maximum germination distances of between 3.7-4.3 cm. Uyoma 47 Brown (10), IESV 92038/2-SH (10.7), and IESV 92036-SH (10.7) recorded the lowest mean numbers of induced germinated Striga seeds and distances of the furthest germinated Striga seeds of 1.5,2.5 and 2.4 cm respectively. The induced few and low mean maximum distances of germinated Striga seeds in Uyoma 47 Brown, IESV 92038/2-SH, and IESV 92036-SH indicate secretion of low amounts of Striga germination stimulant associated with potential resistance to Striga in these genotypes. These findings compare well with that of Mohamed et al. (2010) who carried out an in vitro technique for studying specific Striga resistance mechanisms in sorghum and reported maximum germination distances of 0.058, 0.3, 1.8, and 2.1 cm respectively in the resistant sorghum genotypes SRN 39, Framida, Dobbs and Serena and 2.9cm and 2.8cm for the susceptible varieties Shanqui-Red and IS 4225.
The differences exhibited in the maximum germination distances and numbers of induced germinated Striga seeds is associated with genotypic differences for secretion of strigolactones in the evaluated sorghum genotypes. Higher maximum germination distances together with higher numbers of induced germinated Striga seeds suggest higher amounts of secreted strigolactones in sorghum genotypes, an indication of potential susceptibility of the genotypes to Striga weed. These findings compare well with those of Yohannes et al. (2016) who evaluated sorghum genotypes for secretion of low Striga germination stimulant and Reda et al. (1994) who screened maize genotypes for low Striga asiatica stimulant production. Yohannes et al. (2016) reported reduced secretion of germination stimulant and subsequent Striga resistance in the evaluated landraces compared to the commercial checks while Reda et al. (1994) reported a high correlation between maximum germination distance and numbers of induced germinated Striga seeds. Both Yohannes et al. (2016) and Reda et al. (1994) concluded that high Striga germination percentages suggested high germination stimulant production and susceptibility of genotypes and vice versa. The observed low numbers of induced germinated Striga seeds in Uyoma 47 Brown, IESV 92038/2-SH, and IESV 92036-SH could have been due to root germination-inhibitory compounds that may have interfered with the germination response of the conditioned Striga seeds (Mohamed et al., 2010). However, such a hypothesis needs further investigation.
The study also revealed high numbers of induced germinated Striga seeds around the root periphery (Plate 1a) which indicated increased concentrations of secreted Striga germination stimulant closer to the root zone. This finding conforms with that of Karaya et al. (2012) who studied variations in Striga germination stimulants produced in maize and noted that the closer the Striga seeds to the source of stimulant the higher the numbers of seeds stimulated to germinate and vice versa. Similarly, Hess et al. (1991) reported host’s germination stimulant to be exuded and concentrated mainly in a distance closer to the radius of the root from the root apex. Studies by Yoneyama et al. (2010) also suggested that only Striga seeds within the root rhizosphere are capable of germinating upon the perception of root strigolactones.
Different developmental stages of the germinated Striga seeds were also observed in some of the sorghum genotypes. T53B revealed a developing radicle (Plate 1b) while the presence of an intrusive cell at the tip of the radicle was observed in E117B (Plate 1c) and even the establishment of a haustorium connection with Nyadundo 1 sorghum root (Plate 1d). The observations are comparable with those of Mohamed et al. (2010) and Omanya et al. (2004) who observed similar growth responses. The observed growth response could have been due to the secretion of a secondary metabolite in the genotypes’ host roots which was required for the development of haustorium. The establishment of haustorium connection with Nyadundo 1 sorghum root suggested that this variety failed to initiate a post-attachment hypersensitivity defense reaction mechanism to Striga hence it's susceptible to the weed.


The association between (maximum germination distances and numbers of induced germinated Striga seeds) and level of field Striga-host infestation levels among the genotypes are presented in Table 7.
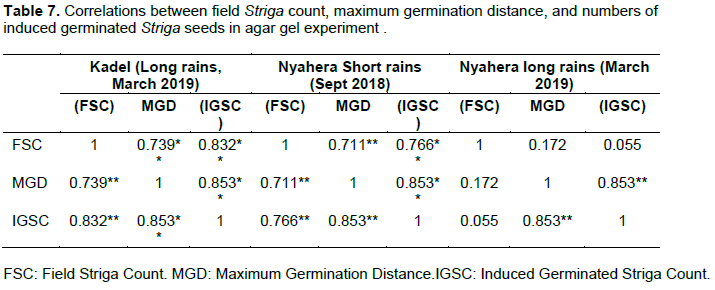
The data showed field Striga counts in the Kadel and Nyahera sites except for Nyahera’s long rains of March 2019 to be highly significant (P < 0.001) and positively correlate with both maximum germination distance and numbers of induced germinated Striga seeds. This, therefore, emphasizes the role of strigolactones in influencing field-host Striga infestation levels. These findings compare well with those of Xie et al. (2008) and Hess et al. (1992) who established a close association between strigolactone levels analyzed under laboratory conditions and field levels of Striga infection.
CONCLUSION
This study obtained significant variations among the genotypes in response to Striga infestations both under field and laboratory conditions. Striga weed significantly reduced sorghum performance in terms of grain yield, shoot dry mass, and plant height with susceptible genotypes suffering severe losses and recording higher Striga damage ratings. This study has identified four Striga tolerant (T53B, N57, N68, and T30 B) and Striga resistant (C26, IESV 92038/2-SH, IESV 92036-SH, and Uyoma 47 Brown,) sorghum genotypes based on both field and laboratory Striga screening techniques. The identified genotypes can be used directly by farmers to improve their yields in Striga-prone areas or for further breeding. The study recommends further research to identify possible Striga resistance mechanisms that the different varieties exhibited.
CONFLICT OF INTEREST
The authors have not declared any conflict of interest.
ACKNOWLEDGMENTS
The authors are grateful to Mcknight Foundation for the partial scholarship. The support from School of Agriculture, Natural resources and Environmental Studies, Rongo University, Kenya and School of Agriculture and Food Security, Maseno University, Kenya is appreciated.
REFERENCES
|
Abate M, Mekbib F, Hussien T, Bayu W, Reda F (2014). Assessment of genetic diversity in sorghum ('Sorghum bicolor'(L.) Moench) for reactions to' Striga hermonthica'(Del.) Benth. Australian Journal of Crop Science 8(8):1248-1256. |
|
|
Abbas T, Zahir ZA, Naveed M, Kremer RJ (2018). Limitations of existing weed control practices necessitate development of alternative techniques based on biological approaches. In Advances in Agronomy: Academic Press pp. 239-280. |
|
|
Akaogu IC, Badu AB, Adetimirin VO, Vroh BI, Oyekunle M, Akinwale RO (2013). Genetic diversity assessment of extra-early maturing yellow maize inbreds and hybrid performance in Striga-infested and Striga-free environments. The Journal of Agricultural Science 151(4):519-537. |
|
|
Aragaw G, Chala A, Terefe H (2021). Spatial distribution and association of factors influencing sorghum anthracnose (Colletotrichum sublineolum) epidemics in Eastern Ethiopia. International Journal of Pest Management 67(1):20-31. |
|
|
Atera EA, Itoh K, Onyango JC (2011). Evaluation of ecologies and severity of Striga weed on rice in sub-Saharan Africa. Agriculture and biology journal of North America 2(5):752-760. |
|
|
Badu AB, Akinwale RO (2011). Cultivar evaluation and trait analysis of tropical early maturing maize under Striga-infested and Striga-free environments. Field Crops Research 121(1):186-194. |
|
|
Badu-Apraku B, Fakorede M A B (2017). Breeding for Striga Resistance. In Advances in Genetic Enhancement of Early and Extra-Early Maize for Sub-Saharan Africa. Springer, Cham. pp. 245-286. |
|
|
Bahadur A, Batool A, Nasir F, Jiang S, Mingsen Q, Zhang Q, Feng H (2019). Mechanistic insights into arbuscular mycorrhizal fungi-mediated drought stress tolerance in plants. International journal of molecular sciences 20(17):4199. |
|
|
Bejiga T (2019). Effect of Striga Infestation on Sorghum Agronomic Traits and Its Breeding Strategy for Resistance in Ethiopia: A Review. Journal of Natural Sciences Research 9(15):17-24. |
|
|
Beyene T, Egigu MC (2020). Striga hermonthica (Del.) Benth has Dual Negative Effect on its Host Sorghum bicolor (L.) Moench. Turkish Journal of Agriculture-Food Science and Technology 8(1):165-170. |
|
|
Bo AB, Khaitov B, Umurzokov M, Cho KM, Park KW, Choi JS (2020). Biological Control Using Plant Pathogens in Weed Management. Weed & Turfgrass Science 9(1):11-19. |
|
|
Dafaallah AB, Babiker AE (2019). Assessment of the Damage Caused by Striga hermonthica (Del.) Benth. on the Performance of Cereals Hosts in Gadarif State, Eastern Sudan. University of Khartoum Journal of Agricultural Sciences 24(2):291-313. |
|
|
Food and Agriculture Organization of the United Nations Database of agricultural production (FAOSTAT) (2019). Statistical Databases Available at: http://www.fao.org/faostat/en/#data/QC |
|
|
Frost DL, Gurney AL, Press MC, Scholes JD (1997). Striga hermonthica reduces photosynthesis in sorghum: the importance of stomatal limitations and a potential role for ABA. Plant, Cell and Environment 20(4):483-492. |
|
|
Gbèhounou G, Pieterse AH, Verkleij JAC (1996). The decrease in seed germination of Striga hermonthica in Benin in the course of the rainy season is due to a dying-off process. Experientia 52(3):264-267. |
|
|
Gobena D, Shimels M, Rich PJ, Ruyter-Spira C, Bouwmeester H, Kanuganti S, Ejeta G (2017). Mutation in sorghum LOW GERMINATION STIMULANT 1 alters strigolactones and causes Striga resistance. Proceedings of the National Academy of Sciences 114(17):4471-4476. |
|
|
Hadebe ST, Modi AT, Mabhaudhi T (2017). Drought tolerance and water use of cereal crops: A focus on sorghum as a food security crop in sub?Saharan Africa. Journal of Agronomy and Crop Science 203(3):177-191. |
|
|
Haussmann BIG, Hess DE, Reddy BVS, Mukuru SZ, Kayentao M, Welz HG, Geiger HH (2001). Pattern analysis of genotype× environment interaction for Striga resistance and grain yield in African sorghum trials. Euphytica 122(2):297-308. |
|
|
Hess DE, Ejeta G, Butler LG (1991). Research into Germination of Striga Seeds by Sorghum Root Exudates. Proceedings of the 5th International Symposium of Parasitic Weeds, Nairobi, 24-30 June 1991. Available at: http://oar.icrisat.org/id/eprint/4656. |
|
|
Hess DE, Ejeta G, Butler LG (1992).Selecting sorghum genotypes expressing a quantitative biosynthetic trait that confers resistance to Striga. Phytochemistry 31(2):493-497. |
|
|
Joel KA, Muchugi A (2018). Genetic diversity and virulence of Striga hermonthica from Kenya and Uganda on selected sorghum varieties. Nusantara Bioscience 10(2):111-120. |
|
|
Joel KA, Runo S, Muchugi A (2018). Genetic diversity and virulence of Striga hermonthica from Kenya and Uganda on selected sorghum varieties. Nusantara Bioscience 10(2):111-120. |
|
|
Karaya H, Njoroge K, Mugo S, Ariga ES, Kanampiu F, Nderitu JH (2012) Determination of Levels of Striga Germination Stimulants for Maize Gene Bank Accessions and Elite Inbred Lines. International Journal of Plant Production 6(2):209-224. |
|
|
Lee N and Thierfelder C (2017). Weed control under conservation agriculture in dryland smallholder farming systems of southern Africa. A review.Agronomy for Sustainable Development 37(5):1-25. |
|
|
Makete NN, Gohole LS, Opile WR, oduori CA (2017). Effect of Intercropping Finger Millet [Eleusine Coracana (L.) Gaertn.] With Desmodium Intortum on Striga hermonthica Emergence across Planting Seasons. African Journal of Education, Science and Technology 4(1):1-13. |
|
|
Mbogo PO, Dida MM, Owuor B (2016).Effect of Striga hermonthica (Del.) Benth on Yield and Yield Components of Maize (Zea mays L.) Hybrids in Western Kenya. Journal of Agricultural Science 8(8):112-125. |
|
|
Mbuvi DA, Masiga CW, Kuria EK, Masanga J, Wamalwa M, Mohamed, Runo SM (2017). Novel sources of witchweed (Striga) resistance from wild sorghum accessions. Frontiers in plant science 8(116):1-15. |
|
|
Menkir A, Makumbi D, Franco J (2012). Assessment of reaction patterns of hybrids to Striga hermonthica (Del.) Benth. under artificial infestation in Kenya and Nigeria. Crop Science 52(6):2528-2537. |
|
|
Mohamed AH, Housley TL, Ejeta G (2010). An in vitro technique for studying specific Striga resistance mechanisms in sorghum. African Journal of Agricultural Research 5(14):1868-1875. |
|
|
Mohemed N, Charnikhova T, Bakker EJ, van AA, Babiker AG, Bouwmeester HJ (2016). Evaluation of field resistance to Striga hermonthica (Del.) Benth. in Sorghum bicolor (L.) Moench. The relationship with strigolactones. Pest management science 72(11):2082-2090. |
|
|
Mohemed N, Charnikhova T, Fradin EF, Rienstra J, Babiker AG, Bouwmeester HJ (2018). Genetic variation in Sorghum bicolor strigolactones and their role in resistance against Striga hermonthica. Journal of experimental botany 69(9):2415-2430. |
|
|
Mrema E, Shimelis H, Laing M, Bucheyeki T (2017). Farmers' perceptions of sorghum production constraints and Striga control practices in semi-arid areas of Tanzania. International Journal of Pest Management 63(2):146-156. |
|
|
Muui CW, Muasya RM, Nguluu SN, Kambura A, Kathuli P, Mweu B, Odhiambo DO (2019). Sorghum landraces production practices in Nyanza, Coast and Eastern regions, Kenya. Journal of Economics and Sustainable Development 10(10):134-143. |
|
|
Ogenga JO, Mugalavai EM, Nyandiko NO (2018). Impact of Rainfall Variability on Food production under Rainfed Agriculture in Homa Bay County, Kenya. International Journal of Research and Scientific Publications 8(8):857-879. |
|
|
Okeyo SO, Ndirangu SN, Isaboke HN, Njeru LK (2020). Determinants of sorghum productivity among small-scale farmers in Siaya County, Kenya. African Journal of Agricultural Research 16(5):722-731. |
|
|
Omanya GO, Haussmann BIG, Hess DE, Reddy BVS, Kayentao M, Welz HG, Geiger HH (2004). Utility of indirect and direct selection traits for improving Striga resistance in two sorghum recombinant inbred populations. Field crops research 89(2):237-252 |
|
|
Reda F, Butler LG, Ejeta G, Ransom K (1994). Screening of maize genotypes for low Striga Asiatica stimulant production using the'agar gel technique'. African Crop Science Journal 2(2):173-177. |
|
|
Rodenburg J, Bastiaans L, Kropff MJ (2006). Characterization of host tolerance to Striga hermonthica. Euphytica 147(3):353-365. |
|
|
Samejima H, Babiker AG, Takikawa H, Sasaki M, Sugimoto Y (2016). Practicality of the suicidal germination approach for controlling Striga hermonthica. Pest management science 72(11):2035-2042. |
|
|
Shaibu AS, Badu-Apraku B, Ayo-Vaughan MA (2021). Enhancing drought tolerance and Striga hermonthica resistance in maize using newly derived inbred lines from the wild maize relative, Zea diploperennis. Agronomy 11(1):177. |
|
|
Teka HB (2014). Advance research on Striga control: A review. African journal of plant science 8(11):492-506. |
|
|
Van AA, Bastiaans L, Kropff MJ (2000). A comparative study on Striga hermonthica interaction with a sensitive and tolerant sorghum cultivar. Weed Research 40(6):479-493. |
|
|
Vurro M, Boari A, Thiombiano B, Bouwmeester H (2019). Strigolactones and parasitic plants. In Strigolactones, Biology and Applications. Springer: Cham. pp. 89-120. |
|
|
Waters MT, Gutjahr C, Bennett T, Nelson DC (2017).Strigolactone signaling and evolution. Annual Review of Plant Biology 1(68):291-322. |
|
|
Xie X, Yoneyama K, Kusumoto D, Yamada Y, Takeuchi Y, Sugimoto Y, Yoneyama K (2008). Sorgomol, germination stimulant for root parasitic plants, produced by Sorghum bicolor. Tetrahedron Letters 49(13):2066-2068. |
|
|
Yohannes T, Ngugi K, Ariga E, Abraha T, Yao N, Asami P, Monday A (2016). Genotypic variation for low Striga germination stimulation in sorghum "sorghum bicolor (L) moench" landraces from eritrea. American Journal of Plant Sciences 2016 (7):2470-2482. |
|
|
Yohannes T, Ngugi K, Ariga E, Ahonsi M, Yao N, Abraha T (2017). A Diagnostic Appraisal of Sorghum Farming in Striga Endemic Areas of Eritrea. Journal of Agricultural Science 9(5):133-141. |
|
|
Yoneyama K, Awad AA, Xie X, Yoneyama K, Takeuchi Y (2010). Strigolactones as germination stimulants for root parasitic plants. Plant and Cell Physiology 51(7):1095-1103. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


