Full Length Research Paper

ABSTRACT
Paspalum notatum Flüggé (bahiagrass) is the most important forage grass in the native grasslands of southern Brazil; the native germplasm is tetraploid and apomictic. Breeding to produce cultivars has been possible due to chromosome doubling of sexual diploid accessions, allowing hybridizations with apomictic genotypes, generating progenies with variability for agronomic traits and segregating for reproduction mode. The aim of this study was to determine the reproduction mode of selected hybrids derived from intraspecific crosses through cytoembryological analysis and to evaluate the level of apospory expres¬sivity. Eighty one tetraploid hybrids corresponding to 21 families of sexual and apomictic parents were generated and 28 most productive plants based on their superior agronomic performance in the field were evaluated. Sixteen hybrids were reproduced sexually and 12 of them were highly apomictic or facultative apomictic. Sexual hybrids may be used for further crosses in the breeding program while highly apomictic plants produce uniform progeny and become candidates of new cultivars to promote pasture diversification.
Key words: Aposporous apomixis, bahiagrass, intraspecific crosses, plant breeding.
INTRODUCTION
Genetic improvement of apomictic forage grasses has an important role in the economy of beef cattle production systems in the tropics and subtropics. Among warm-season apomictic forage grasses, the genus with the greatest economic and social value are Brachiaria, Panicum, Cenchrus, and Paspalum (Jank et al., 2014).
Paspalum notatum Flügge (bahiagrass) is an important forage species in the grasslands in South America and is used as forage due to its yield, quality, and persistence. Around the world, it is also used as turf and forage (Gates et al., 2004). It is an important provider to native pasture in southern Brazil and also can be used as pioneering species to reduce the negative effects of soil erosion and degradation (Barbosa et al., 2019).
Aposporous apomixis, asexual reproduction through seeds, is a common reproductive mode in polyploid (tetraploid) (2n = 4x = 40) biotypes of bahiagrass, while the natural sexually reproducing plants are diploid (2n = 2x = 20) (Miles, 2007). The reproductive barriers between tetraploids apomictic plants and sexual diploids are a clear obstacle for breeding programs (Zilli et al., 2018). Besides that, the apomixis affects the plant diversity among species and populations (Acuña et al., 2019). By doubling the chromosome number of sexual diploid accessions, sexual tetraploids were produced (Forbes and Burton, 1961; Quarin et al., 2001, 2003; Quesenberry et al., 2010; Weiler et al., 2015). The feasibility of crosses between sexual 4x, used as female parents, and apomictic 4x, used as pollen donors, have been reported widely in the literature (Acuña et al., 2009; 2011; Zilli et al., 2015; Weiler et al., 2018). So, it’s possible to produce populations segregating for apomixis and hybrids that display heterosis for agronomic traits of interest (Acuña et al., 2009, 2011; Ortiz et al., 2020), for example forage yield and cold tolerance as already reported in P. notatum (Acuña et al., 2011; Zilli et al., 2015; Weiler et al., 2018; Barbosa et al., 2019).
The selected genotypes, with superior agronomic characteristics fixed by apomictic reproduction at the tetraploid level in the breeding programs can be considered potential new cultivars (Acuña et al., 2009; Acuña et al., 2011; Jank et al., 2014; Zilli et al., 2015; Novo et al., 2020).
Inheritance and expressivity of apomixis in Paspalum is the consequence of a complex genetic and/or epigenetic control of the trait (Podio et al., 2014). Sexuality is a recessive character and apomixis is conditioned by a single dominant Mendelian factor which is the most widely accepted genetic model for the inheritance of apomixis in grasses (Acuña et al., 2011; Martínez et al., 2001; Ozias-Akins and Van Dijk, 2007; Stein et al., 2004). However, the full expression of the apomictic trait depends upon some pleiotropic effect and incomplete penetrance of dominant allele controlling the apospory development (Martínez et al., 2001). Recent data indicate that apomictic seed development in plants is linked to a dynamic pattern of transcriptional activity in ovule, probably regulated through epigenetic mechanisms (Kumar et al., 2017) as well as to environmental conditions (Acuña et al., 2009; Zilli et al., 2015). Therefore, it is common to occur segregation distortion against apospory (Martínez et al., 2001) and variable levels of apomixis expressivity can be expected among generations (Acuña et al., 2011; Zilli et al., 2015).
In order to use newly developed hybrids in crosses, the reproduction mode and apospory expressivity needs to be reliably assessed to define their usefulness for crop improvement of the species and the progenies to be studied and characterized (Acuña et al., 2007, 2019; Miles, 2007). For screening the reproduction mode, molecular markers give a fast classification of plants, which is very useful for breeding purposes.
However, markers do not provide information concerning the expression level of apospory, necessary for apomictic hybrid selection (Zilli et al., 2018). On the other hand, the cytoembriological method is accurate to identify reproduction mode and apospory expressivity in the hybrids. The objective of this study was to determine the reproduction mode of selected segregating hybrids of bahiagrass derived from intraspecific crosses through cytoembryological analysis and to evaluate the level of apospory expressivity among hybrids, in order to use them as parents for future breeding programs.
MATERIALS AND METHODS
Plant material
The experiment was carried out at the Department of Forage Plants and Agrometeorology of the Agronomics School, Federal University of Rio Grande do Sul (UFRGS), located in the city of Porto Alegre, Rio Grande do Sul State, Brazil (latitude 30° 1'16.13" S and longitude 51°13'23.99” W). Crosses with Paspalum notatum genotypes were performed in the greenhouse to produce hybrids. The female parents were three sexual tetraploids genotypes, C4-4X (Quarin et al., 2001), Q4188 and Q4205 (Quarin et al., 2003). As male parents, the apomictic tetraploid accessions called 30N, 36N, 70N, 48N, 83N, 95N and V4 were selected after evaluation for forage production by Fachinetto et al. (2012).
Hybridization
Crosses were performed in the greenhouse following the methodology described by Burton (1948). In the mother plants, before the anthesis, rooted culms bearing inflorescences were collected and placed in a jar with water. These culms were placed in an artificial fog chamber, with a high level of air humidity to avoid the anthers from dehiscence. In the morning of the next day, the flowers were emasculated, using sharp pointed tweezers to remove the anthers. Non-flowering spikelets were eliminated from the inflorescence. Fresh pollen from the apomictic parent was collected in paper envelopes and later dusted on emasculated inflorescences of the sexual plants. After pollination, the inflorescences were bagged and labelled. Seeds were collected at least 21 days after pollination. The progeny from each cross was referred as a family and a number was given to identify each hybrid. Nine families produced hybrids, which were selected to determine the reproduction mode.
Embryo sac observations
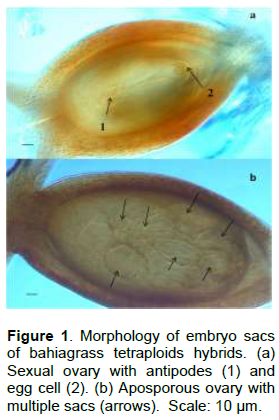
The reproduction mode of progenies was analyzed in inflorescences collected at anthesis (embryo sacs fully developed). The flowers of various inflorescences were dissected and fixed in FAA solution [40 ml 95% ethanol: 14 ml distilled water: 3 ml 40% formalin: 3 ml glacial acetic acid] for 24 h. Subsequently, the flowers were transferred to ethanol 70% and refrigerated. The pistils were dissected and the ovaries were dehydrated with alcohol and clarified with methyl salicylate according to the method established by Young et al. (1979), modified by Acuña et al. (2007). Slides with the ovaries were prepared and observed using a differential interference contrast microscope and some of them were photographed with a Zeiss camera. A minimum of 40 ovules per plant were examined. The reproduction mode was determined based on the observation of embryo sac morphology. The ovules were separated in sexual (presence of antipodal cells) or apomictic (without antipodal cells, multiple or single embryo sacs with the egg apparatus and the central cell) (Acuña et al., 2009; Kumar et al., 2017). The reproduction mode of the male parents was also checked using embryo sac observations.
Evaluation of the reproduction mode and apospory expressivity
The plants were classified as sexual, highly apomictic, and facultative apomictic based on the number of ovules with aposporous embryo sacs. Aborted or immature ovules were not counted (Marcón et al., 2019); only embryo sacs identified as sexual or apomictic were used for classification. Plants with almost all ovules (>90%) bearing aposporous embryo sacs were classified as highly apomictic; plants with less than 90% (1 - 89%) of aposporous embryo sacs on the same inflorescence were classified as facultative apomictic whereas plants without aposporous embryo sacs, bearing only single reduced embryo sacs were classified into sexual. Acuña et al. (2009) and Ortiz et al. (2020) classify the aposporic hybrids as ‘obligate’ or ‘highly apomictic’ when almost all ovules (>90%) show one or several aposporous embryo sacs (AESs).
RESULTS AND DISCUSSION
From the crosses, we obtained 81 tetraploid hybrids of sexual and apomictic parents, corresponding to 21 families (C4-4X x 30N; C4-4X x 36N; C4-4X x 48N; C4-4X x 70N; C4-4X x 83N; C4-4X x 95N; C4-4X x V4; Q4188 x 30N; Q4188 x 36N; Q4188 x 48N; Q4188 x 70N; Q4188 x 83N; Q4188 x 95N; Q4188 x V4; Q4205 x 30N; Q4205 x 36N; Q4205 x 48N; Q4205 x 70N; Q4205 x 83N; Q4205 x 95N and Q4205 x V4). The hybrid progenies were transplanted to the field, at the Agronomic Experimental Station of the Federal University of Rio Grande do Sul (UFRGS), Rio Grande do Sul State, Brazil (latitude 30°05´S and longitude 51°39´W) and evaluated to estimate their agronomic value. Five cuts were taken during two years. The variable total production of dry mass of leaves (LDM) showed the highest phenotypic correlation with the production of total dry mass (TDM). The 28 selected hybrids from nine families (Table 1) presented higher forage production than the ecotypes used as parents and also from the standards. Cumulative TDM varied from 191.0 (g. plant-1) (plant 225) until 414.1 g.plant-1 (plant 437), while the cultivar Pensacola produced 76.1 g.plant-1. The reproduction mode and apospory expressivity were then assessed in these selected hybrids.
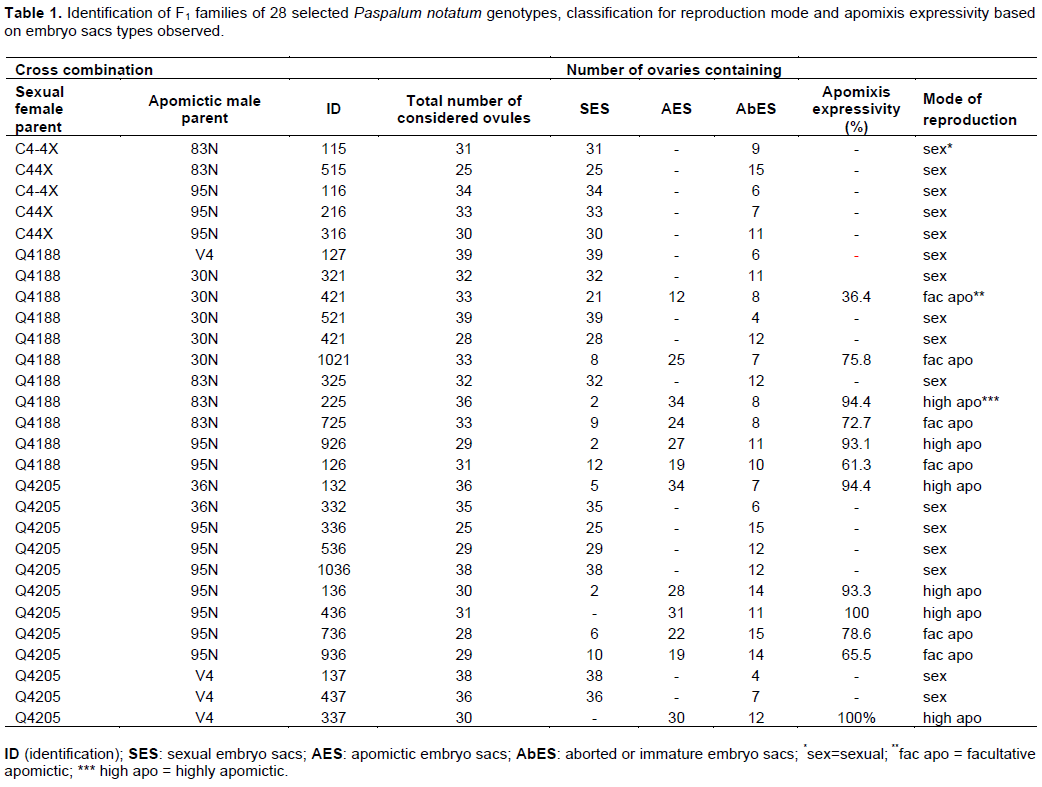
Classification of reproduction mode
As expected, the hybridization between induced sexual tetraploid and apomictic plants results in F1 plants that segregated for reproduction mode. Abnormal, atrophied and sterile ovaries was found in almost all plants (Table 1). Ovules bearing both sexual and apomictic embryo sacs were not observed. In the same family, some hybrids were sexual while others were apomictic, because a single dominant Mendelian factor controls apospory in bahiagrass, where sexuality behaves as recessive (aaaa).
The male parents used in the crosses confirmed the apomictic reproduction as expected. All the parents presented almost all ovules (>90%) bearing aposporous embryo sacs and were classified as highly apomictic. Sixteen plants were classified as sexual because only a single reduced embryo sacs were observed in their ovules. Twelve plants had embryo sac morphology compatible with apomictic mode of reproduction and were classified as apomictic (Table 1 and Figure 1). The apospory expressivity was variable in the 12 apomict hybrids, with high and low levels observed. Six apomictic plants (named 421, 1021, 725, 126, 736 and 936) showed intermediate level of expressivity and were classified as facultative apomictic. The remaining six plants (named 225, 926, 132, 136, 436 and 337) were classified as highly apomictic because aposporous embryo sacs were observed in more than 90% of their ovules (Table 1), representing 21.43% of the total hybrids.
Acuña et al. (2009) reported the mode of reproduction and apospory expressivity of 173 hybrids from different families of bahiagrass. Of these, 19 plants were highly apomictic; 16 were classified as facultative apomictic and 138 plants were classified as sexual. Apomictic progenies represented 20% of the plants. The ratio between sexual and apomictic tetraploid hybrids of bahiagrass in the first-generation (FGTH) and in the second-generation (SGTH) was determined by Acuña et al. (2011). For the FGTH it was from the 4.3:1 (sexual:apomictic) and for the SGTH was of 4.6:1. Although similar, there was a strong distortion with an excess of sexual progeny, but the segregation patterns for apomixis remained constant through hybridization cycles. The expressivity was variable between generations and only 3% of the progenies were classified as highly apomictic, maybe due environmental differences between years of evaluation.
Zilli et al. (2015) also recorded variable expressivity for apospory in hybrids of bahiagrass. For 32% of the hybrids classified as aposporic, were observed highly levels of expressivity, between 81 and 100%. The variable expression of apomixis among the progenies remains uncertain. It is possible that a single genomic region is responsible for triggering the trait. However, others important elements as genetic or epigenetic factors may be affecting the expressivity (Acuña et al., 2009; Zilli et al., 2015; Ortiz et al., 2020). Genetic analyses in Paspalum showed that apomixis occur by various mechanisms, which share three common developmental components: apomeiosis, parthenogenesis and unbalanced endosperm formation (Kumar et al., 2017; Ortiz et al., 2020). The single locus is blocked in terms of recombination, hemizygosity and heavy cytosine methylation. Podio et al. (2014) evaluated the DNA methylation state of the apomixis-controlling region (ACR) in Paspalum simplex and Paspalum notatum. The study suggested that factors controlling repression of parthenogenesis might be inactivated in apomictic Paspalum by DNA methylation, while demethylation induces a significant depression of parthenogenesis. Environmental conditions/stress provide evidences to support the theory that apomixis is epigenetically regulated. So, it’s possible to reach desirable variability and adaptive advantages without altering DNA sequence (Rodrigues and Koltnow, 2005).
Identification of highly apomictic, facultative apomictic and sexual hybrids has an important role in the use in the next phases in a breeding program. Sexual genotypes can be used in further crosses, while highly apomictic hybrids identified in this research can potentially produce uniform progenies and stability in the successive reproductive cycles (Acuña et al., 2019). However, variable levels of apospory expressivity can reduce the probability of fixing hybrids exhibiting heterosis for traits of interest by apomixis. Acuña et al. (2011) asserted that the low proportion of highly apomictic hybrids derived from artificial hybridization difficult the selection of superior genotypes highly apomictic. Even so, the apomictic hybrids selected in this research encouraged us to develop a breeding scheme that allows the register, protection and later the released of the apomictic hybrids as new cultivars.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGMENTS
The authors appreciate CAPES (Brazilian Federal Agency for Support and Evaluation of Graduate Education), CNPq (Brazilian Council for Scientific and Technological Development) and the South-Brazilian Association for the Promotion of Forage Research (SULPASTO) for financial support and grants.
REFERENCES
|
Acuña CA, Blount AR, Quesenberry KH, Hanna WW, Kenworthy KE (2007). Reproductive characterization of bahiagrass germplasm. Crop Science 47(4):1711-1717. |
|
|
Acuña CA, Blount AR, Quesenberry KH, Kenworthy KE, Hanna WW (2009). Bahiagrass tetraploid germplasm: reproductive and agronomic characterization of segregating progeny. Crop Science 49(2):581-588. |
|
|
Acuña CA, Blount AR, Quesenberry KH, Kenworthy KE, Hanna WW (2011). Tetraploid bahiagrass hybrids: breeding technique, genetic variability and proportion of heterotic hybrids. Euphytica 179(2):227-235. |
|
|
Acuña CAA, Martínez EJ, Zilli AL, Brugnoli EA, Espinoza F, Marcón F, Urbani MH, Quarin CL (2019). Reproductive systems in Paspalum: relevance for germplasm collection and conservation, breeding techniques, and adoption of released cultivars. Frontiers in Plant Science 10:1377. |
|
|
Barbosa MR, daMotta EAM, Machado JM, Krycki KC, Conterato IF, Weiler RL, Dall'Agnol M, Simioni C (2019). Herbage accumulation of bahiagrass hybrids in two different environments in southern Brazil. Pesquisa Agropecuária Gaúcha 25(1/2):58-69 |
|
|
Burton G (1948). Artificial fog chamber facilitates Paspalum emasculation. Journal American Society of Agronomy 40:281-282. |
|
|
Fachinetto JM, Schneider R, Hubber KGC, Dall'Agnol M (2012). Agronomic evaluation and persistence analysis in a collection of Paspalum notatum Flügge (Poaceae) accessions. Revista brasileira de Ciências Agrárias 7(1):189-195. |
|
|
Forbes I, Burton GW (1961). Induction of tetraploidy and a rapid field method of detecting induced tetraploidy in Pen¬sacola Bahiagrass. Crop Science 1:383-384. |
|
|
Gates RN, Quarin CL, Pedreira CGS (2004). Bahiagrass. In: Moser LE, Burson BL and Sollenberger, LE (Eds.), Warm-season (C4) grasses. Madison, WI: Agronomy Monography 45. pp. 651-680. |
|
|
Jank L, Barrios SC, Do Vale CB, Resende RMS and Alves GF (2014). The value of improved pastures to Brazilian beef production. Crop and Pasture Science 65(11):1132-1137. |
|
|
Kumar S, Saxena S, Gupta MC (2017). Marker-assisted screening of breeding populations of an apomictic grass Cenchrus ciliaris L. segregating for the mode of reproduction. Crop Breeding and Applied Biotechnology 17(1): 10-17. |
|
|
Marcón F, Martínez EJ, Rodríguez GR, Zilli AL, Brugnoli EA, Acuña CA (2019) Genetic distance and the relationship with heterosis and reproductive behavior in tetraploid bahiagrass hybrids. Molecular Breeding 39(6):1-13. |
|
|
Martínez EJ, Urbani MH, Quarin CL, Ortiz JPA (2001). Inheritance of apospory in bahiagrass, Paspalum notatum. Hereditas 135(1):19-25. |
|
|
Miles JW (2007). Apomixis for cultivar development in tropi¬cal forage grasses. Crop Science 47:S-238. |
|
|
Novo PE, Acuña CA, Urbani NH, Galdeano F, Espinoza F, Quarin CL (2020). Genetic transfer from several apomictic tetraploid Paspalum species to an elite group of sexual plants. Crop Science 60(4):1997-2007. |
|
|
Ortiz J, Pupilli F, Acuña CA, Leblanc O, Pessino SC (2020). How to Become an Apomixis Model: The Multifaceted Case of Paspalum. Genes 11(9): 974. |
|
|
Ozias-Akins P, Van Dijk PJ (2007). Mendelian genetics of apomixis in plants. Annual Review Genetics 41:509-537. |
|
|
Podio M, Cáceres ME, Samoluk SS, Seijo JG, Pessino SC, Ortiz JPA, Pupilli F (2014). A methylation status analysis of the apomixis-specific region in Paspalum spp. suggests an epigenetic control of parthenogenesis. Journal of Experimental Botany 65(22):6411-6424. |
|
|
Quarin CL, Espinoza F, Martínez EJ, Pessino SC, Bovo OA (2001). A rise of ploidy level induces the expression of apomixis in Paspalum notatum. Sexual Plant Reproduction 13(5):243-249. |
|
|
Quarin CL, Urbani MH, Blount AR, Martínez EJ, Hack CM, Burton GW, Quesenberry KH (2003). Regis¬tration of Q4188 and Q4205, sexual tetraploid germplasm lines of Bahiagrass. Crop Science 43(2):745-746. |
|
|
Quesenberry KH, Dampier JM, Lee YY, Smith RL, Acuña CA (2010). Doubling the chromosome number of bahia¬grass via tissue culture. Euphytica 175(1):43-50. |
|
|
Rodrigues JCM, Koltnow MAG (2005). Epigenetic aspects of sexual and asexual seed development. Acta Biologica Cracoviensia Series Botanica 47:37-49. |
|
|
Stein J, Quarin CL, Martínez EJ, Pessino SC, Ortiz JPA (2004). Tetraploid races of Paspalum notatum show polysomic inheritance and preferential chromosome pairing around the apospory-controlling locus. Theorical Applied Genetics 109(1):186-191. |
|
|
Weiler LR, Dall'Agnol M, Simioni C, Krycki KC, Pereira EA, Machado JM, Motta EAM (2018). Intraespecific tetraploid hybrids of Paspalum notatum: agronomic evaluation of segregation progeny. Scientia Agricola 75(1):36-42. |
|
|
Weiler RL, Krycki KC, Guerra D, Simioni C, Dall'Agnol M (2015). Chromosome doubling in Paspalum notatum var. saure (cultivar Pensacola). Crop Breeding and Applied Biotechnology 15(2):106-111. |
|
|
Young BA, Sherwood RT, Bashaw EC (1979). Cleared-pistil and thicksectioning techniques for detecting aposporus apomixis in grasses. Canadian Journal of Botany 57(15):1668-1672. |
|
|
Zilli AL, Acuña CA, Shulz RR, Brugnoli EA, Guidalevich V, Quarin C L, Martínez EJ (2018). Widening the gene pool of sexual tetraploid bahiagrass: generation and reproductive characterization of a sexual synthetic tetraploid population. Crop Science 58(2):762-772. |
|
|
Zilli AL, Brugnoli EA, Marcón F, Billa MB, Rios EF, Martínez EJ, Acuña CA (2015). Heterosis and expressiv¬ity of apospory in tetraploid Bahiagrass hybrids. Crop Science 55(3):1189-1201. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


