Full Length Research Paper

ABSTRACT
Twenty-eight hybrids of Zea mays L. produced in 2015-16 from 8×8 half diallel mating design were evaluated along with their parents in 2016-2017 under saline condition to find out genetic action and potency ratio of some agronomic and yield contributing traits. Genetic action and potency ratio were analyzed following Hayman’s diallel analyses and Smith’s formula, respectively. Genetic parameter like additive variance (D), dominance variance (H1), proportion of positive and negative genes in the parents (H2), relative frequency of dominant and recessive alleles in the parents (F), dominance effect over all loci in heterozygous phase (h2) suggested that traits like days to tasseling, days to silking, anthesis silking interval, ear height and number of grains/plant are governed by dominant alleles in these traits. Contrariwise, the data of plant height and 100-grain weight were shown to have higher frequency of recessive alleles. Proportion of genes with positive and negative [(4DH1)0.5 + F]/[(4DH1)0.5 – F] effects suggested the asymmetrical distribution of dominant and recessive alleles for all the traits except yield/plant. Under saline condition heritability in narrow sense (h2n) was found very low which indicates a possible strong influence of stress in the growing environment. These parameters along with Vr-Wr graphs and potency ratio indicated overdominance in desirable direction, and thus, heterosis breeding is important to improve those traits in maize under salinity.
Key words: Maize, salinity, genetic parameter, Vr-Wr, potence, inheritance.
INTRODUCTION
Maize (Zea mays) is the solely cultivated species of the genus ‘Zea’ and the tribe Maydeae. It is the oldest crop species domesticated as food crop in the world which is a C4 crop. Being a C4 crop, maize possesses most potentiality of ensuring food security in the coming days (Chohan, 2012). It can be cultivable in broad climatic conditions worldwide. Globally, maize occupies the third most important position as a crop. Maize is compatible with wide range of agro-climatic zones. The suitability of maize to diverse environments is unparalleled to any other crops (Hossain et al., 2016). Maize can be grown from below sea level to higher altitudes like >3000 m.
The rainfall range is around 250 mm to greater than 5000 mm per year. The life cycle of maize can vary from 3 to 10 months (Sheikh et al., 2017). According to FAO (2016), in the year 2016 total maize cultivation area was 188 million hectare (ha) while production was 1050.1 million ton and average yield of 5.64 ton ha-1.
Moreover, the multipurpose uses of maize coupled with its maximum grain yield make it popular globally. It is used as food for human, feed for poultry, livestock and fish. In Bangladesh, poultry and fish feed industry is increasing. In parallel with this, maize grain demand is also increasing in many folds (Ali et al., 2009). Therefore, high yielding maize hybrid developing program has been executed worldwide. To achieve yield improvement through genetic improvement approaches, gene action determination is essential for conveying necessary breeding strategies. By understanding the nature of gene action, governing a specific trait is essential for improving that particular character which will ultimately increase the yield any crop. Additionally, the choice of a fit breeding program depends on the understanding of the nature of gene action of yield and yield related traits. On one hand, hybrid development program prefers dominance gene action, whereas on the other hand, additive gene action can efficiently improve any character (Hossain et al., 2016). Diallel analysis system was extensively used in heredity related research to investigate the inheritance strength of any important trait in a set of genotypes (Yan and Hunt, 2002). Components of genetic control can assist breeders to select competent parents for a crossing program. Thus, gene action study facilitates decision making of a suitable breeding procedure for genetic improvement of various quantitative traits (Jinks and Hayman, 1963; Walters and Morton, 1978; Reza et al., 2004; Begum et al., 2018).
Information about inheritance pattern of any specific trait can be obtained from Hayman numerical approach (Hayman, 1954a, b) while Griffing (1956) provides a feature on genetic action of parental lines. Diallel crosses have been used for a long time in genetic research to determine the inheritance of a trait among a set of genotypes and to identify superior parents for hybrid or cultivar development (Aliu et al., 2009). These methods have been highly practiced in different crops like maize (Njeri et al., 2017; Owusu et al., 2017; Lay and Razdan, 2017; Brahmbhatt et al., 2018), rice (Huang et al., 2015; Kundan et al., 2013), Brassica (Tian et al., 2017) and cassava (Tumuhimbise et al., 2014). Hence, it is necessary to understand the nature and magnitude of gene action as well as combining ability of yield and its attributes.
Besides knowing the genetics of the germplasm, it also becomes a crucial problem to breed for problematic areas of the world. Salinity is a growing threat for crop production and growth throughout the world. In arid and semi-arid regions, salinity level increases due to water shortage and elevated temperature. In Bangladesh, salinity affected area is increasing every year. In Bangladesh, during 1973, salinity affected 83.3 million hectares of land; this was increased to 102 million hectares by the year 2000. After that, salinity affected a recorded 105.6 million hectares during 2009 (SRDI, 2010). Over the last 35 years, salinity has increased around 26% in the coastal region of Bangladesh (Mahmuduzzaman et al., 2014). Therefore, saline tolerant maize variety development is a time demanding research in Bangladesh.
The present investigation of 8×8 diallel cross maize without reciprocal crosses was undertaken to supplement genetic parameters interpretations, pinpoint which parents contain the preponderance of dominance/ recessive genes with increasing/decreasing character attributes, and isolate superior inbred lines and better combining parents for utilizing their salinity affected area of Bangladesh in future breeding programs. Considering those, we analyzed the genetic action of maize in an 8×8 diallel progeny growing in saline soil to understand the nature of gene action in saline condition for developing suitable hybrids for saline area.
MATERIALS AND METHODS
Plant materials
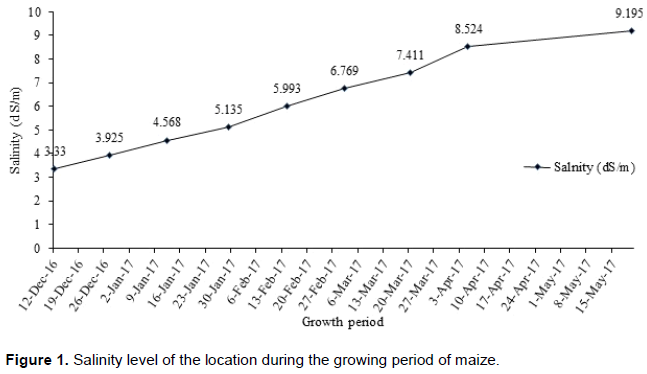
Eight maize inbred lines (CZ-28, CZ-29, BIL-65, CZ-36, CZ-12, CZ-26, CZ-24 and 9MG) collected from International Maize and Wheat Improvement Center (CIMMYT) and Plant Breeding Division, Bangladesh Agricultural Research Institute (BARI) were crossed in a diallel fashion excluding the reciprocals during the rabi season (winter) in 2015-2016 at BARI, Gazipur, Bangladesh. The resulting 28 F1’s and their 8 parents were evaluated in a randomized complete block design (RCBD) with three replications in saline area of Agricultural Research Station, Benarpota, Satkhira (22.43° N latitude and 89.06° E longitude), Bangladesh in the subsequent (winter) season of 2016-2017. The soil salinity level was also monitored in the growing location by a digital EC meter (Hanna 993310). It was found that salinity level increased throughout the plant growth period which ranged from 3.3 to 9.2 dS/m (Figure 1). This increasing tendency of salinity is due to depletion of soil moisture, because there was no rain in this period.
Experiment settings, crop management and data recording
Seeds of each entry were sown in two rows of 4 m plot. The spacing between rows was 60 cm and plant to plant distance was 25 cm. Fertilizers were applied at 250, 55, 110, 40, 5 and 1.5 kg ha-1 of N, P, K, S, Zn and B, respectively. One plant per hill was maintained after proper thinning. Agronomic and yield related traits of the plants were recorded on five randomly selected competitive plants. Agronomic trait includes days totasseling, days to silking, anthesis silking interval (days), plant height (cm) and ear height (cm). In each plots when 50% plants produced male flower, it was recorded as days to tasseling. Similarly, for days to silking, 50% silking of the plot was recorded. For the trait anthesis silking interval (ASI), days between 50% anthesis and 50% silking was considered. For plant height, ear height as well as yield related recorded traits like number of grains/plants, 100-grain weight (g) and grain yield (g) were measured from randomly selected 10 plants for each replication.
Statistical analysis
The statistical analysis of variance was performed as described in Sharma (1988). The genetic parameters of Hayman’s graphical and numerical approach (1954a, b) like additive variance (D), dominance variance (H1), proportion of positive and negative genes in the parents (H2), relative frequency of dominant and recessive alleles in the parents (F), dominance effect over all loci in heterozygous phase (h2). environmental variance (E), along with their allied parameters, mean degree of dominance (H1/D)0.5, proportion of genes with positive and negative [(4DH1)0.5 + F]/[(4DH1)0.5 – F] effects in the parents and heritability in narrow sense (h2n) was calculated as described in Sharma (1988). Vr-Wr graphs were plotted in MS Excel according to Singh and Chaudhary (1985).
Potency ratio was calculated according to Smith (1952) to determine the degree of dominance as follows: P = F1 – M.P./{0.5 (P2 -P1)}. Where, P: relative potency of gene set, F1: first generation mean, P1: the mean of lower parent, P2: the mean of higher parent, M.P.: mid-parents value = (P1 + P2)/2.
RESULTS AND DISCUSSION
Analysis of variance
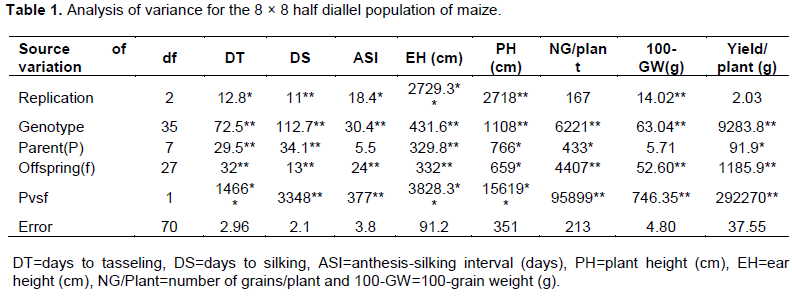
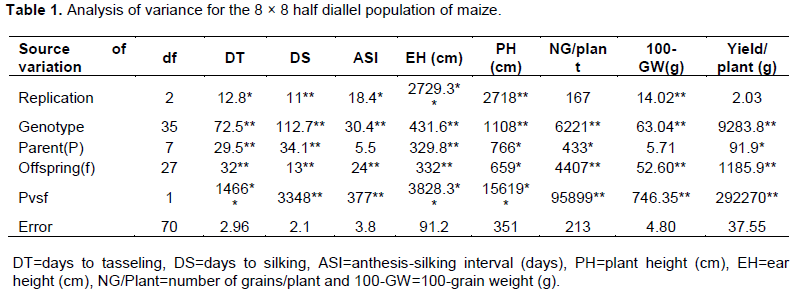
The analysis of variance showed that mean squares for the parent and diallel hybrid lines were significant at P ≤0.05 for the traits anthesis silking interval and days to tasseling while the values of other traits were highly significant (P≤0.01) (Table 1). Similarly, the mean sum of squares of genotypes were highly significant for all the recorded eight traits. In case of parents, all the traits, except anthesis silking interval and 100-grain weight, were significant. On the other hand, the values of offspring and parent vs offspring were highly significant. The significant variation among the values of the traits signified the existence of divergence and genetic variation among individuals (Chohan et al., 2012). Importantly, the highly significant mean sum of squares at 1 degree of freedom (df) indicated the possibility of heterosis for the traits.
Genetic parameter of different traits in maize under salinity
Gene action was clarified by genetical analysis of Hayman (1954a, b). The analysis of variance components indicated that both additive (D) and dominance variance (H1 and h2) are all significant for the 8 studied traits (Table 2). This result indicated the possibility of the traits are conditioned by both additive and dominance gene action. However, dominance components are more predominant than additive component. The dominance is also reflected by overall dominance effect over all loci in heterozygous phase (h2). Importantly, higher values of H2 than H1 with negative sign indicated the unbalanced distribution of dominant and recessive alleles with negative effects in the parents under salinity stress (Table 2). The size of mean degree of dominance (H1/D)0.5 was categorized as (H1/D)0.5 = 0, mean no dominance, (H1/D)0.5 = 1, mean complete dominance, (H1/D)0.5>1, mean over dominance and (H1/D)0.5<1 mean partial dominance. The study showed that under salinity condition, the values of (H1/D)0.5 were >1 for all the traits suggesting prevailing over dominance in all the loci for all the traits. The environmental component (E) was significant and in case of ASI, it was higher than D and H1 (Table 2) noticing higher environmental effect in expressing the trait. The proportion of dominant (p) and recessive (q) alleles is ascertained by the ration [(4DH1)0.5 + F]/[(4DH1)0.5 – F]. Its value explains that [(4DH1)0.5 + F]/[(4DH1)0.5 – F]@ 1.0 means nearly equal proportion of dominance and recessive alleles in parents, that is, symmetrical distribution; p=q=0.5. If its value is >1.0, it refers to an excess of dominant alleles and the minority of recessive alleles (p>q) while <1.0 means minority of dominant alleles and excess of recessive alleles (p<q). In the present study, this ration was >1.0 for days to tasseling, days to silking, anthesis silking interval, ear height and number of grain/plant (Table 2) indicating of excess of dominant alleles in these traits. Contrariwise, the data of plant height and 100-grain weight exhibited higher frequency of recessive alleles. However, yield/plant had almost symmetrical distribution of the alleles.
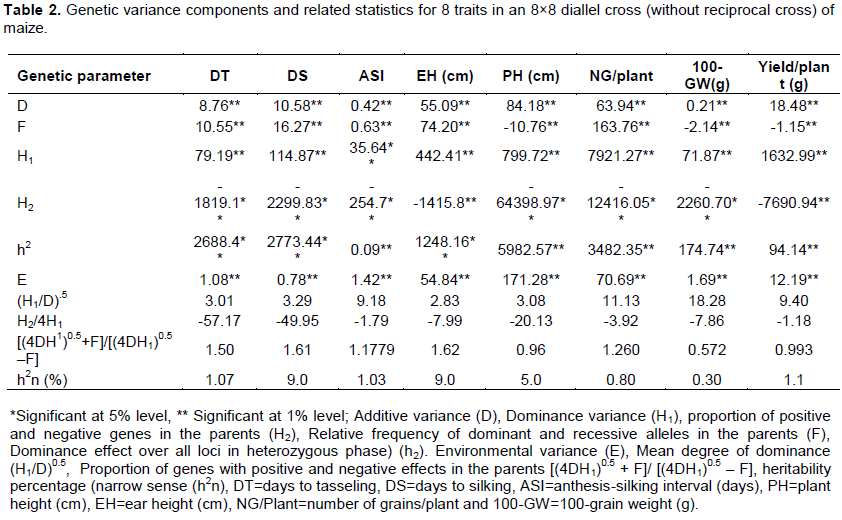
Another important genetic parameter, mean covariance of additive and dominant variance expressed by F is presented in Table 2. The value of F expressed that if F=0, it means balanced distribution (p=q=0.5); F>0 (+) means dominant alleles are more frequent than recessive alleles (p>q); F>0 (-) means recessives are more prevalent than dominant alleles (p<q). The value of F (>0) for days to tasseling, days to silking, anthesis silking interval, ear height and number of grain/plantalso confirmed excess of dominant alleles. The proportion of dominant genes with positive or negative effects in parents is determined by the ratio: H2/4H1 with the maximum theoretical value of 0.25, which stands up when p=q=0.5 in all loci. A deviation from 0.25 would be irregular when p¹q. The values of H2/4H1 deviated for all the traits, thus, dominance genes having increasing and decreasing effects on all the traits are irregularly distributed in the parents. These findings confirmed our previous study (Begum et al., 2018). However, the lower heritability estimate (h2n) predicted that the traits were very much influenced by environmental effect. Present results are in good agreement with Hussain et al. (2014) and Irshad-ul-Haq et al. (2010) where they found over dominance for pollen shedding and anthesis -silking interval under drought condition.
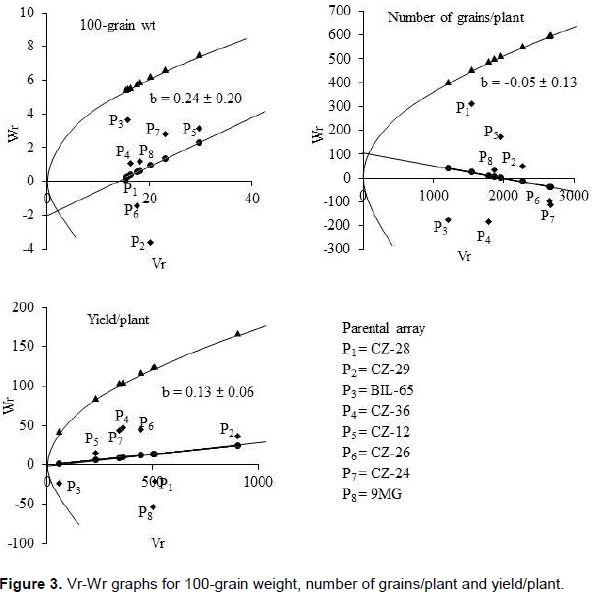
Vr-Wr graph
Hayman’s graphical approach to diallel analysis is based on monogenic additive model (Figures 2 and 3). This approach helps to understand the genetic action of the parental lines involved in a diallel mating system which is reported in many crops (EL-Hosary, 2014; Begum et al., 2018; Iftekharuddaula et al., 2008; Boye-Goni and Marcarian, 1985; Rohman et al., 2006). Vr-Wr graphs are the two dimensional depiction made based on the parental variance (Vr) and parent offspring co-variance (Wr). The average level of degree of dominance could be understood by the nature (+/-) and magnitude of ‘a’ (the Y intercept). The regression line passed above the point of origin suggesting partial dominance for controlling the trait. The regression line intersected Wr axis above the point of origin suggesting partial dominance for controlling the trait. The regression line intersected below the point of origin suggesting over dominance for controlling the trait. The distribution of array points indicated parental order of dominance which is provided by (Wr+Vr) values. For any trait, the parents containing dominant alleles will fall closer to the point of origin and showed lower value of (Wr+Vr) while, parent with maximum frequency of recessive alleles will fall far from the origin.
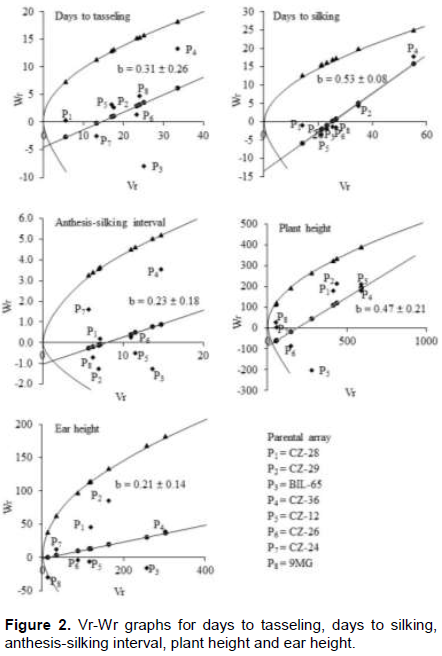
The Wr/Vr graphs of days to tasseling, days to silking, anthesis-silking interval, ear height and plant height are presented in Figure 2. In most of the cases, the regression line incepted below the origin suggesting presence of overdominance controlling the trains which was also depicted by (H1/D)0.5. Overdominance in different traits in maize was also reported in water stress (Hussain et al., 2014). CZ-36 exhibited maximum frequency of recessive alleles for days to tasseling, silking, anthesis-silking interval, and ear height being farthest from the origin while CZ-28 had maximum frequency of dominant alleles for days to tasseling and silking (Figure 2). On the other hand, both BIL-65 and CZ-36 seemed to possess more recessive alleles for plant height and ear height. The parents 9MG, CZ-24, CZ-28 and CZ-29 had more frequency of dominant alleles for anthesis-silking interval while 9MG, CZ-24, CZ-12 and CZ-26 for plant height, 9MG, CZ-24 and CZ-26 for ear height (Figure 2). However, this type of study under salinity is very limited. Hussain et al. (2014) and Irshad-ul-Haq et al. (2010) reported similar results under drought condition.
For yield and related traits, Vr-Wr graphs are presented in Figure 3. The regression lines of 100-grain weights and yield/plant passed below the origin suggesting overdominance action for those traits. On the other hand, the regression line of number of grains/plant passed above the origin and partial dominance effect was present for this trait. The distribution of parents showed most of them possess more dominant alleles for 100-grain weight while CZ-12 had more recessive alleles. For number of grains/plant, maximum parents had higher recessive allele frequency. On the other hand, parent CZ-29 exhibited maximum alleles frequency for yield/plant due to being farthest from origin while BIL-65 had maximum dominant alleles. Importantly, other parents showed more dominant alleles for this trait. Therefore, the distribution of dominant and recessive alleles is not symmetrical which was confirmed in genetic component analysis in Table 2. These findings are in good agreement with those of Saleem et al. (2002), Watto et al. (2002), Betran et al. (2003), Prakash and Ganguli (2004), Ali et al. (2007), Irshad-ul-Haq (2010), Khodarahmpour (2011) and Hussain et al. (2014) for grain number and grain yield per plant under drought stress.
Potence ratio
The potence ratio (PR) of 28 F1 crosses are shown in Table 3. The positive values ratio specified the degree of dominance ranging from partial to over-dominance. Again, negative values indicate the degrees of recessiveness ranging from partial to under recessiveness (Solieman et al., 2013).
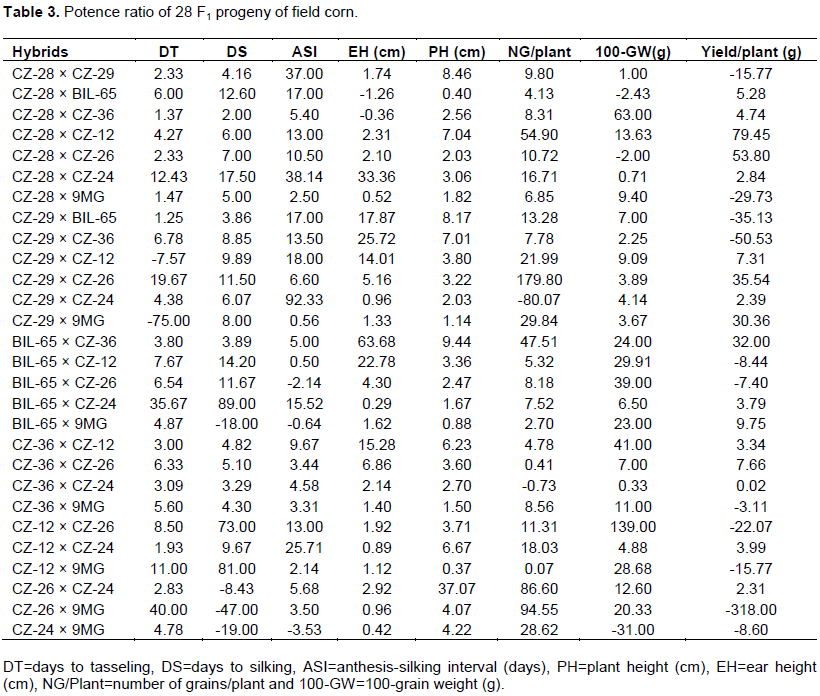
For the trait DT, PR value ranged from -25 (CZ-29 × 9 MG) to 40 (CZ-26 × 9 MG). Among them no crosses showed complete dominance (-1.0) and all the 28 crosses exhibited over-dominance (>±1). For the trait DS, the range of PR was -47.0 (CZ-26 × 9 MG) to 89 (BIL-65 × CZ-24) where all the crosses showed over-dominance (>±1). The PR of the trait ASI was distributed from -3.53 to 92.33. Apart from two crosses, all other crosses were over dominant. In case of the trait EH, potence ratio spectrum is -1.26 (CZ-28 × BIL-65) to 63.68 (BIL-65 × CZ-36). Out of 28 crosses, seven (CZ-28 × CZ-36, CZ-28 × 9 MG, CZ-29 × CZ-24, BIL-65 × CZ-24, CZ-12 × CZ-24, CZ-26 × 9 MG and CZ-24 × 9 MG) showed partial dominance whereas the rest of the crosses showed over dominance (PR >±1). PR for PH ranged from 0.37 (CZ-12 × 9 MG) to -37.07 (CZ-26 × CZ-24). Three crosses showed partial dominance while PR of the remaining crosses indicated over dominance. PR of the trait number of grain/plant ranges from -8.07 (CZ12 × CZ-24) to 94.55 (CZ-26 × 9 MG). The crosses CZ-36 × CZ-24, CZ-36 × CZ-26 and CZ-12 × 9 MG showed partial dominance. Other 25 crosses showed over dominance (PR >±1). Considering the PR of 100 grain weight (100-GW), the range was -31.00 (CZ-24 × 9 MG) to 40.00 (CZ-36 × CZ-12). While one cross (CZ-28 × CZ-29) showed complete dominance (PR=1.00), two other crosses (CZ-36 × CZ-26) and (CZ-28 × CZ-24) showed partial dominance (PR<1.00); and all the other crosses showed over dominance (PR >±1). In case of the last studied trait, yield/plant PR ranged from -47.00 (CZ-26 × 9 MG) to 79.45 (CZ-28 × CZ-12). The cross CZ-36 × CZ-24 showed absence of dominance for this trait. The rest of the crosses exhibited over dominant (PR >±1). Previously, Begum et al. (2018) reported similar results for different traits of maize under normal growing condition.
CONCLUSION
Genetic parameters, Vr-Wr and potence ratio suggested that the traits are governed by over dominance gene action. Study of the hybrids suggests that all the 28 cross combinations showed over dominance for the traits. However, the lower values of h2n indicated that the traits were highly influenced by the growing environment. Considering all of this, the data will be important in exploring heterosis breeding to improve those traits in maize under salinity. However, this type of result under salinity stress is not available. Therefore, further research in this circumstance can provide important information for developing saline tolerant varieties of crops.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Ali G, Rather AC, Ishfaq A, Dar SA, Wani SA, Khan MN (2007). Gene action for grain yield and its attributes in maize (Zea mays L.). International Journal of Agricultural Science 3(2):278-281. |
|
|
Ali MY, Waddington SR, Hodson DP, Timsina J, Dixon J (2009). Maize-rice cropping systems in Bangladesh: status and research needs. Journal of Agricultural Science and Technology 3(6):35-53. |
|
|
Aliu S, Fetahu S, Kaciu S, Salillari A (2009). Combining ability study for kernel yield per ear of maize (Zea mays L.) hybrid. 44th Croatian and 4th International Symposium on Agriculture pp. 476-480. |
|
|
Begum S, Alam SS, Omy SH, Amiruzzaman A, Rohman MM (2018). Inheritance and combing ability in maize using a 7×7 diallel cross. Journal of Plant Breeding and Crop Science 10(9):239-248. |
|
|
Betran FJ, Ribaut JM, Beck D, Leon DG (2003). Genetic diversity, specific combining ability and heterosis in tropical maize under stress and non-stress environments. Crop Science 43:797-806. |
|
|
Boye-Goni SR, Marcarian V (1985). Diallel analysis of aluminum tolerance in selected lines of grain sorghum. Crop Science 25:749-752. |
|
|
Brahmbhatt BN, Kuchhadiya GV, Gosai MA, Joshi NR, Kanjariya KG (2018). Study of combining ability through diallel crosses in maize (Zea mays L.) for grain yield and protein content. Journal of Pharmacognosy Photochemistry 7(2):1754-1755. |
|
|
Chohan MSM, Saleem M, Ahsan M, Asghar M (2012). Genetic analysis of water stress tolerance and various morpho-physiological traits in Zea mays L. using graphical approach. Pakistan Journal of Nutrition 11(5):489-500. |
|
|
EL-Hosary AAA (2014). Comparison between some methods of diallel cross analysis in maize Egypt. Journal of Plant Breeding 18(4):715-736. |
|
|
Food Agriculture Organization (FAO) (2016). Agriculture Organization of the United Nations Statistics Division. |
|
|
Griffing B (1956). Concept of general and specific combining ability in relation to diallel crossing systems. Australian Journal of Biological Science 9:463-493. |
|
|
Hayman BI (1954a). The analysis of variance of diallel tables. Biometrics 10:235-244. |
|
|
Hayman BI (1954b). The theory and analysis of diallel crosses. Genetics 39:789-809. |
|
|
Hossain F, Muthusamy V, Bhat JS, Jha SK, Zunjare R, Das A, Sarika K, Kumar R (2016). Maize. M. Singh, S. Kumar (eds.), Broadening the Genetic Base of Grain Cereals, |
|
|
Huang M, Chen L, Chen Z (2015). Diallel analysis of combining ability and heterosis for yield and yield components in rice by using positive loci. Euphytica 205:37. |
|
|
Hussain M, Shah KN, Ghafoor A, Kiani TT, Mahmood T (2014). Genetic analysis for grain yield and various morphological traits in maize (Zea mays L.) under normal and water stress environments. Journal of Animal and Plant Science 24(4):1230-1240. |
|
|
Iftekharuddaula KM, Newaz MA, Salam MA, Akter K (2008). Genetic analysis for panicle characters in diallel cross of rice. Bangladesh Journal of Agricultural Research 33(3):631-638. |
|
|
Irshad-ul-Haq MS, Ajmal S, Munir M, Gulfaraz M (2010). Gene action studies of different quantitative traits in maize. Pakistan Journal of Botany 42(2):1021-1030. |
|
|
Jinks JL, Hayman BL (1963). The analysis of diallel crosses. Maize Genetic Cooperative Newsletter 27:48-54. |
|
|
Khodarahmpour Z (2011). Gene action studies of different traits in maize (Zea mays L.) under heat stress and normal conditions. Journal of American Science 7(5):449-454. |
|
|
Kundan K, Shailesh M, Binod K (2013). Combining ability analysis in diallel crosses of wheat (Triticum aestivum L). The Bioscan 8:1557-1560. |
|
|
Lay P, Razdan AK (2017). Genetic Analysis of Grain Yield and its Components of Maize (Zea mays L.) Inbred Lines. International Journal of Current Microbiology and Applied Science 6(7):1366-1372. |
|
|
Mahmuduzzaman M, Ahmed ZU, Nuruzzaman AKM, Ahmed FRS (2014). Causes of salinity intrusion in coastal belt of Bangladesh. International Journal of Plant Research 4(4):8-13. |
|
|
Njeri SG, Makumbi D, Warburton ML, Diallo A, Jumbo MB, Chemining'wa G (2017). Genetic analysis of tropical quality protein maize (Zea mays L.) germplasm. Euphytica 213:261. |
|
|
Owusu GA, Nyadanu D, Obeng-Antwi K, Amoah RA, Danso FC, Amissah S (2017). Estimating gene action, combining ability and heterosis for grain yield and agronomic traits in extra-early maturing yellow maize single-crosses under three agro-ecologies of Ghana. Euphytica 213:287. |
|
|
Prakash S, Ganguli DK (2004). Combining ability for various yield component characters in maize (Zea mays L.). Journal of Research, Birsa Agricultural University 16(1): 55-60. |
|
|
Reza A, Yazdisamadi B, Zali A, Tallei A, Zeinali H, Rezael A (2004). Estimate of heterosis and combining ability in maize (Zea mays L.) using diallel crossing method. In: Genetic Variation for plant breeding. EUCARPIA, BOKU, Vienna, Austria. pp. 395-397. |
|
|
Rohman MM, Sultana R, Podder R, Islam ATMT, Islam MK, Islam MS (2006). Nature of gene action in barley (Hordeum vulgare L.). Asian Journal of Plant Science 5(2):170-173. |
|
|
Saleem M, Shahzad K, Javid M, Ahmed A (2002). Genetic analysis for various quantitative traits in maize (Zea mays L.) inbred lines. International Journal of Agriculture and Biology 4(3):379-382. |
|
|
Sharma JR (1988). Statistical and Biometrical Techniques in Plant Breeding. New Age International (P) Limited, Delhi, pp 178-203. |
|
|
Sheikh FA, Dar ZA, Sofi PA, Lone AA (2017). Recent Advances in Breeding for Abiotic Stress (Drought) Tolerance in Maize. International Journal of Current Microbiology and Applied Science 6(4):2226-2243. |
|
|
Singh RK, Chaudhary BD (1985). Biometrical methods in quantitative genetics analysis. Kalyani Publishers, New Delhi pp. 102-127. |
|
|
Smith HH (1952). Fixing Transgressive Vigour in Nicotia narustica. In: Gowen JW, editor. Heterosis. Ames (IA), Iowa State University Press. pp. 161-174. |
|
|
Solieman THI, El-Gabry MAH, Abido AI (2013). Heterosis, potence ratio and correlation of some important characters in tomato (Solanum lycopersicon L.). Scientia Horticulturae 150:25-30. |
|
|
Sprague GF, Tatum LA (1942). General vs. specific combining ability in single crosses of corn. Journal of American Society of Agronomy 34:923-932. |
|
|
SRDI (2010). Saline Soils of Bangladesh. Soil Resources Development Institute, Ministry of Agriculture, Dhaka, Bangladesh. |
|
|
Tian HY, Channa SA, Hu SW (2017). Relationships between genetic distance, combining ability and heterosis in rapeseed (Brassica napus L.). Euphytica 213:1. |
|
|
Tumuhimbise R, Melis R, Shanahan P (2014). Diallel analysis of early storage root yield and disease resistance traits in cassava (Manihot esculenta Crantz). Field Crops Research 167:86-93. |
|
|
Walters DS, Morton JR (1978). On the analysis of variance of a half diallel table. Biometrics 34:91-94. |
|
|
Yan W, Hunt LA (2002). Biplot analysis of diallel data. Crop Science 42:21-30. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


