Full Length Research Paper

ABSTRACT
The use of multiple traits for drought study affirms the complexity of drought tolerance in cowpea. Despite the availability of several traits for drought tolerance evaluation, the rapid screening technique used by many scientists for seedling drought in wooden boxes is the simplest method for screening large populations. The objective of this study was to select drought tolerant cowpea recombinant lines developed from a drought tolerant and susceptible parent using the wooden box screening technique. Two hundred Recombinant Inbred Lines (RILS) an F2:6 generation were used for the study. The parents were drought tolerant line crossed with susceptible line. Screening was done in wooden boxes and plants stressed for 4 weeks and in two sessions. Leaf wilting, relative water content, chlorophyll content during stress, and recovery from drought data were taken. Results from this study showed significant differences (p < 0.05) for relative chlorophyll content for the 4 weeks of water stress and relative water content taken on the second week of water stress for all 200 inbred lines, but no significant differences were observed for the parental checks. Relative water contents taken for RILS during water-stress ranged between 70-20% for drought tolerant and drought susceptible lines respectively. The parental lines used as checks both had relative water contents of 60%. Relative water content for the second experiment ranged between 74-22% for tolerant and susceptible RILS respectively. About 12 inbred lines consistently performed well for recovery, 13 RILS were susceptible. RILS that maintained a higher relative water and chlorophyll contents, with high proportion of survived seedlings were 11. Potential seedling drought tolerant RILS have been identified.
Key words: Cowpea, leaf wilting, chlorophyll, recombinant inbred lines, drought tolerance, relative water content.
INTRODUCTION
Cowpea [Vigna unguiculata (L.) Walp.] being an excellent source of protein, contains all the essential amino acids (Vasconcelos et al., 2010; Oliveira et al., 2016). It is also rich in carbohydrates, vitamins and minerals, besides having great fibre content and low-fat content, constituting an important food component in several countries including Ghana (Freire Filho et al., 2012; SARI, 2014). As such the crop is grown in all the Savanna ecological regions of Ghana where drought and heat stress compounded by poor soil fertility conditions limit the production of many other crops. Among the abiotic constraints to production of the crop, drought is one of the most important stresses because, the crop is typically grown in the Savanna and Sahel regions where rainfall amount and distribution are unreliable. The severity and occurrence of drought stress is expected to increase as a result of global environmental changes, causing major implications for food supply (Fan et al., 2015). Compounding this is an increasing world population that requires a rise in food production by more than 70% before 2050 (Godfray et al., 2010; Parfitt et al., 2010; Wallace, 2000).
Verbree et al. (2015) indicated that, the use of multiple traits for evaluation of drought tolerance affirms the complexity of drought tolerance in cowpeas compared to other crops. Also, Aliyu and Makinde (2016) and Swain et al. (2017) indicated that cowpea breeding is largely based on selection of parents, followed by hybridization, in order to form a base population and generation advancement with simultaneous selection for more than one trait (Batieno et al., 2016). Therefore, the most common breeding method consists of screening under controlled drought stress; the offspring derived from populations, followed by the assessment of selected genotypes at a location where drought occurs frequently, and testing the most promising genotypes for yield potential and yield stability in multiple sites representing the target ecology (Batieno et al., 2016; Ortiz et al., 2002). Analysis of genetic divergence seeks to identify parents for creating populations with genetic variability and consequent genetic gain in successive selection cycles (Santos et al., 2016). Though various cowpea-breeding materials such as F2, F3 and backcross populations have been used for drought tolerance studies in cowpea, the empirical approach mainly relies on the use of recombinant inbred lines (RIL) to enable the consistent evaluation of performance and understanding of genotype-by-environment interaction, as the intensity and frequency of naturally occurring drought stress are not predictable. The RIL population, developed through single seed descent of several selfed generations consists of individual lines carrying dispersed homozygous segments of a parental chromosome. The objective of this study was to select drought tolerant cowpea recombinant lines developed from a drought tolerant and susceptible parents using the wooden box screening technique.
MATERIALS AND METHODS
Population development
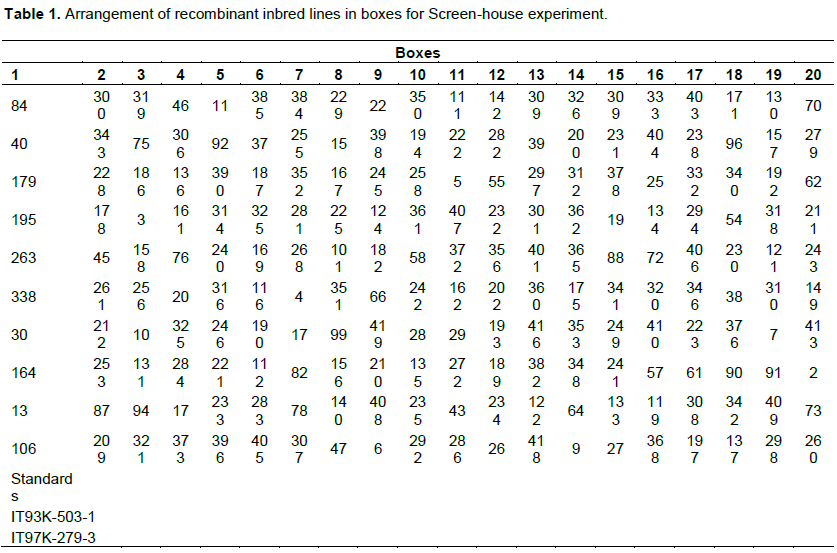
Four hundred and fifty Recombinant Inbred Lines (RILS) were developed through single seed decent and an F2:6 generation was obtained between 2010 and 2015. The parents for the developed population were IT-93k-503-1; a drought tolerant and a medium maturing, indeterminate line crossed with IT97k-279-3; an early maturing line with determinate character. These two lines were obtained from the International Institute of Tropical Agriculture (IITA) Kano, Nigeria.
Geographical location and experimental design for screen-house experiment
The study was carried out at Savanna Agricultural Research Institute (SARI), Nyankpala. Nyankpala is located in the Northern Guinea Savanna Zone with a mean annual rainfall of about 1000 mm. It is located on latitude 9°, 25″ N and longitude 0°, 58″ W with an altitude of 183 m above sea level. Wooden boxes of 130 cm length, 65 cm width, 15 cm depth, and 2.5 cm thick planks were arranged in a screen house as described by Singh et al. (1999a). The boxes were lined with perforated polyethylene sheets and filled with one: one mixture of top soil and sand which averaged 39% sand, 2% clay, and 59% silt by analysis of the composited soil used to fill the boxes. The composite soil had a pH value of 6 and an organic matter content of 6% with N, P, K of 0.05, 3, and 45% respectively. The soil had a bulk density of 1.33 g/cm3. Bulk density (g/cm3) was manually estimated as: Dry soil weight (g) / Soil volume (cm3). The boxes were filled to 12 cm depth leaving about 3 cm space on the top for watering. The polyethylene lining along the sides and bottom of the boxes ensured even distribution of water. A spirit level was used to ensure a flat soil surface on the boxes before and after watering. Equidistant holes were made in straight rows 10 cm apart with a hill to hill distance of 5 cm within the rows. Each box was watered thoroughly and allowed to drain for two days before planting. The moisture content was then taken before planting using the WET sensor with the HH2 moisture meter (Plate 3). Two seeds were sown in each hole and were thinned to one plant per hill one week after germination. Each box contained one row each of 10 recombinant inbred lines plus the two parental checks making it 12 lines in each box (Table 1.). Seedling drought screening was done in two sessions using augmented design because of large size of recombinant inbred lines to be tested (200), as well as limited space for experiment arrangement in the screening house. Screen house experiment one (session one) was done between June and July 2015, while screen house experiment two (session two) was done between October and November 2015, as a repeat to confirm seedling screening for drought tolerance. The boxes were watered daily using a small watering can until the appearance of the first trifoliate leaf, after which watering was stopped (Plate 1 and 2).

Climatic data for the period of the screen-house experiment
The mean average temperature within the screen house during the period of the experiment for the two sessions ranged between 26.4 and 30.7°C, similarly, the mean relative humidity ranged between 47 and 83%.
Drought treatment
Moisture stress was applied by watering the plants until the full expansion of the first trifoliate leaves (two weeks after planting), after which watering was withdrawn for four weeks, in order to take drought response measurements (Muchero et al., 2008). The plants were then re-watered twice a week for a period of two weeks, before taking recovery measurements. The SM300 soil moisture meters with the HH2 reader manufactured by the DELTA- T Devices Ltd, UK, was used together to monitor the soil moisture directly in the soil on a weekly basis during the water stress imposition until the end of the experiment (Plate 3).

Weekly chlorophyll meter readings
Soil Plant Analytical Development (SPAD) chlorophyll meter reading was taken at a weekly interval from the first week of the experiment until the end of the experiment. The Minolta handheld portable SCMR meter (SPAD- 502 Minolta, Tokyo, Japan), was used as per Markwell et al. (1995)to acquire a rapid estimate of the leaf chlorophyll content in nmol/cm. The measurements were taken on the upper most collared leaf halfway from the leaf base to the tip and halfway from the midrib to the leaf margin. Four measurements were taken per plant and the results averaged resulting in a single value to represent each inbred line. In recording the Specific chlorophyll metre readings (SCMR), care was taken to ensure that the SPAD meter sensor fully covered the leaf lamina and the interference from veins and midribs were avoided.
Leaf wilting
Leaf wilting index were calculated from the first week of stress to the final week using Mai-Kodomi (MAIK) scale, by Mai-Kodomi et al. (1999); total number of leaves per plant; number of leaves showing wilting signs with the following wilting scale: 0 = no sign of wilting, 1 = 25% of wilting 2 = moderate wilting, 50%, 3 = yellow and brown leaves with 75% wilting, 4 = completely wilted.
Relative water content measurements (RWC)
Relative water content (RWC) was calculated on new fully expanded leaflets after the second and fourth weeks of stress, as outlined in Bogale et al. (2011). The leaves for RWC were detached from the plant between 10 am and 2 pm during bright days, in order to avoid the effects of weather conditions on water loss from the detached leaves. Immediately after cutting at the base of the lamina, the leaves were weighed to obtain the fresh weight (FW). After weighing, the leaves were soaked in deionized water for 48 hours at room temperature for rehydration: and then re-weighed for turgid weight (TW). The leaves were then dried in an oven at 70°C for 72 h before dry weight (DW) measurements were taken. The RWC was calculated as follows:
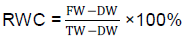 (Bogale et al., 2011).
(Bogale et al., 2011).
Visual vigour rating of seedlings under water stress
The following parameters were recorded after stressing the plants: wilting, using Mai-Kodomi (MAIK) (Mai-Kodomi et al. 1999) scales: total number of leaves per plant; number of leaves showing wilting signs per plant; and RWC. The Leaf Wilting Index (LWI) were calculated weekly, from the first week to the final week of stress, as the ratio between leaves showing wilting signs and the total number of leaves per plant. Both the IB and MAIK scales were scored on a weekly basis from the second week until the end of the stress period.
Recovery from drought
After re-watering, data were collected on: Survival count (SC): number of surviving plants per genotype.
Recovery rate (RR)
Recovery rate was computed as:
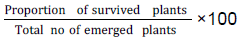 (Fatokun et al. 2012)
(Fatokun et al. 2012)
Plate 4 shows the reaction of inbred lines to water stress treatment up to the fourth week, followed by recovery after watering resumed (Plate 5 and 6).


Data analysis
Data was analysed using GenStat edition version 12, and SAS (version 9.4; SAS Institute, Cary NC). Phenotypic correlation and regression analysis were then performed using PROC CORR and PROG REG to determine the association among the physiological parameters. Significant means were separated using the least significant difference at 5% probability level (LSD 0.05).
RESULTS
Variation in seedling stage drought tolerance based on box screening technique
Chlorophyll content
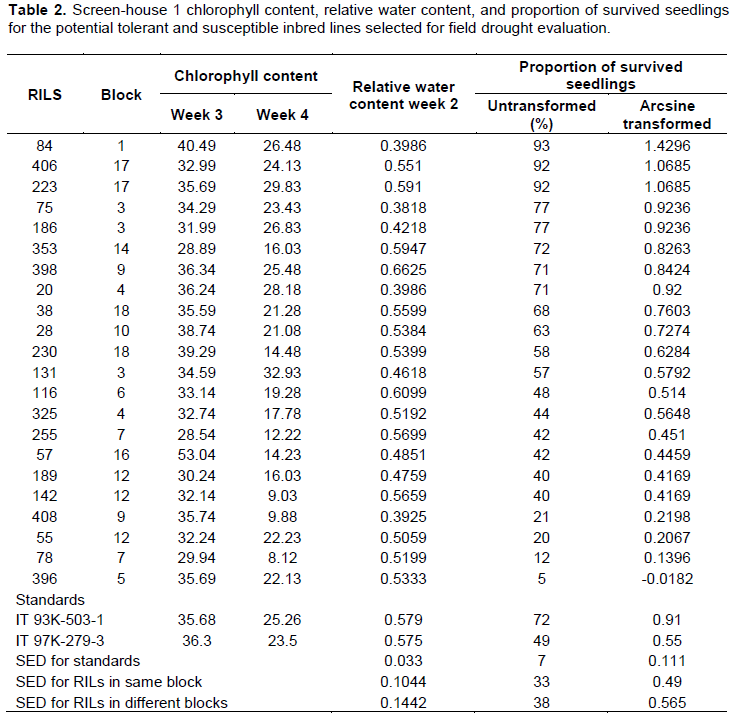
There were variations among the inbred lines screened for drought in the screen-house for both screen-house experiments one and two. The chlorophyll content in nmol/mg gradually decreased over the period of the stress imposition with chlorophyll content ranging between 40.49 and 28.89 with the parental checks of 35.68 and 36.30 for IT93K- 503-1 and IT97K-279-3 respectively for week one and 32.93 and 9.03 with the parental checks recording 25.26 and 23.50 for week four of stress for screen-house experiment one. Chlorophyll measurements for screen-house two showed similar pattern of variation in terms of reduction in chlorophyll as the stress imposition advanced. The chlorophyll for week one ranged between 46.72 and 38.52 with the parental checks recording values in the range of 39.56 and 35.43; week two values ranged between 42.11 and 30.97 with parental values of 38.56 and 33.45 for IT93K-503-1 and IT97K-279-3 respectively. Also, leaf chlorophyll content for week four ranged between 29.7 and 9.00 with the parental checks ranging between 18.25 and 25.85.
Relative water content for screen-house experiment one and experiment two
The relative water contents ranged between 20 and 70% for both drought susceptible and drought tolerant inbred lines across the population for screening. The parental lines used as checks, had relative water contents of 60% for Screen-house. Experiment one and a relative water content range of 74 and 22% for Screen-house experiment two with the parental checks of 40 and 35% for IT93K-503-1 and IT97K-279-3 respectively. These results are based on the summarized and selected potential tolerant and susceptible lines used for field screening for drought (Tables 2 and 3).
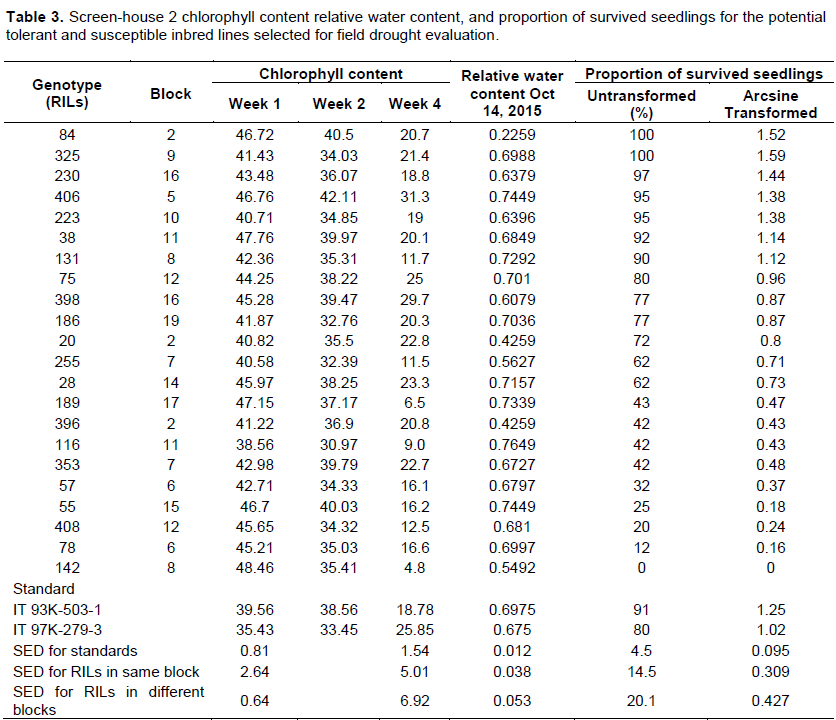
Proportion of survival for screen-house experiments one and two
The proportion of recovery and survival for screen-house experiment one ranged between 93 and 5% for the potential tolerant and susceptible inbred lines with the parental checks scores of about 72 and 49% for IT93K- 503-1 and IT97K-279-3 respectively. The proportion of survival for screen-house two were relatively higher compared to screen-house one. Survival for some inbred lines were 100%, whereas the lowest scored 54% with the parental checks 91 and 80% for IT93K-503-1 and IT97k-279-3 respectively. Results for the selected potential drought tolerant and susceptible inbred lines for screen-house screening at the seedling stage for experiment one and two are summarized and presented in Tables 2 and 3 respectively.
Mean squares of measured traits for cowpea inbred lines evaluated in the screen -house for tolerance to drought
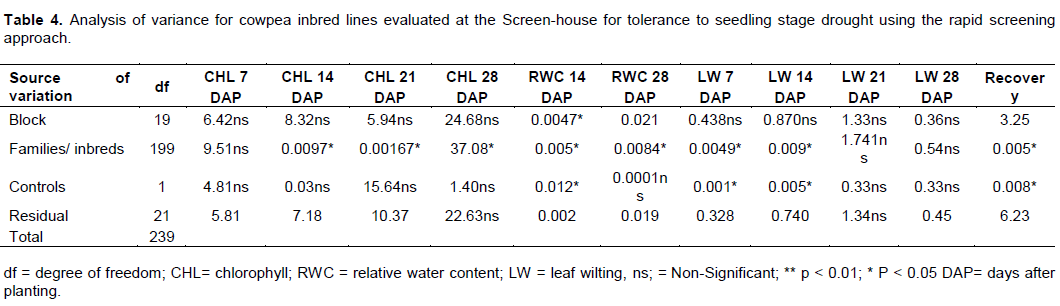
There were significant differences (p < 0.05) for chlorophyll contents for the 200 inbred lines used in the study. Chlorophyll contents taken during 7, 14, 21 and 28 days after water stress treatment significantly varied among the inbred lines, but no significant differences were observed for the parental checks used for the study (Table 4). Significant differences were observed for relative water content taken at 14 days of stress imposition, leaf wilting and recovery.
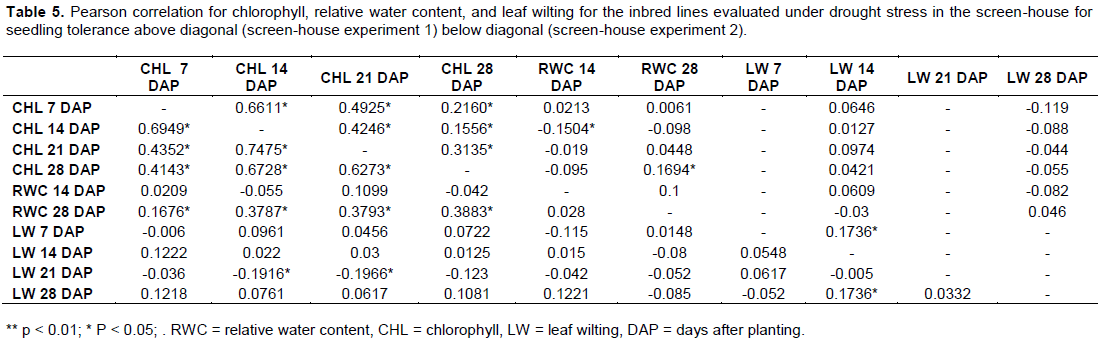
Pearson correlation coefficient between relative water content, chlorophyll and leaf wilting
Relative water contents at 14 days of water stress correlated negatively with chlorophyll at 7, 14, and 28 days of stress imposition. Relative water content also correlated negatively with leaf wilting at 7 days but positively correlated with leaf wilting at 14, 21 and 28 days of stress imposition. Relative water content at 28 days of stress however, correlated positively with chlorophyll at 7, 14, 21 and 28 days of stress and leaf wilting at 7 and 28 days but correlated negatively with leaf wilting at 14 and 21 days during the stress imposition (Table 5). Leaf wilting after 7 days of water stress correlated positively with chlorophyll for 7, 14, 21 and 28 days of water stress, however, leaf wilting 14, 21 and 28 days negatively correlated with chlorophyll at 7, 14, 21 and 28 days of stress imposition.
DISCUSSION
Breeding for drought tolerance for cowpea improvement using various techniques, has been exploited by many research scientists all over the world, especially in cowpea producing countries. The use of wooden boxes has been found to be the most appropriate, fast and rapid screening approach for seedling drought tolerance for shoot related traits such as the relative water content, (Aref et al., 2013; Bogale et al., 2011; Pirzad et al., 2011; Pungulani et al, 2013); leaf wilting, chlorophyll contents, (Steidle Neto et al. 2017), and the estimation of proportion of survived seedlings after recovery (Olubunmi, 2015; Muchero et al., 2008; Tomar and Kumar, 2004). This has been the most successful approach for evaluating large populations and subsequent selection for field drought assessment of genotypes (Singh et al., 1999b). In this study, the use of wooden box technique to rapidly screen 200 inbred lines over a four-week period and a repeat for confirmation was helpful to discriminate among inbred lines for tolerance to seedling drought. This corroborates with similar study by Soltys-Khan et al. (2016) who reported that a fast screening tool would be helpful in selecting valuable genotypes with defined growth strategies that translates to drought tolerance and are therefore suitable for breeding experiments since the phenotype is controlled by genes derived from both parents.
In the current study, water stress significantly reduced the chlorophyll contents for both the parental checks and the inbred lines used. This result corroborates with similar drought related studies report that, drought related traits such as leaf area index, leaf area duration and chlorophyll contents decreased as the water stress duration increased (Khan et al., 2015; Pirzad et al., 2011; Deblonde and Ledent, 2000). Also, Yuan et al. (2016), who in their study on the effects of different levels of water stress on leaf photosynthetic characteristics and antioxidant enzyme activities of greenhouse tomato reported that water stress decreased stomatal conductance net photosynthetic rate, photosynthetic rate at light saturation, and chlorophyll content in all development stages of tomato resulting in yield reduction.
Water deficit affects the photosynthetic ability of plants by changing the content and components of chlorophyll, reducing the net CO2 uptake by leaves, thereby decreasing activities of enzymes in the Calvin cycle (Cornic and Massacci, 1996; Gong et al., 2005; Lawlor and Tezara, 2009). In this current study, as water stress imposition progressed, there were reductions in the chlorophyll contents for all the inbred lines.
Similar study by Tuberosa (2012), on a three-week drought treatment on Katahdin-derived potato cultivars resulted in a decrease in the leaf water content of the cultivars in relative to the control. There was significant variation among the inbred lines for relative water content (RWC) in this study. This observation corroborates with Zegaoui et al. (2017) who reported that two cowpea land races originating from the arid area, maintained a higher RWC over the duration of the drought stress and transpired less than the landrace from the temperate area. Studies by Bogale et al. (2011) and Pirzad et al. (2011) on wheat genotypes and Matricaria chamomilla respectively for drought tolerance reported that changes in the relative water content of leaves are considered as a sensitive indicator of drought stress and more useful indicator of plant water balance (Bogale et al., 2011; Clavel et al., 2005). Therefore, the inbred lines with higher relative water content may have a high potential for survival under field drought conditions and subsequently give good yields. Also, relative chlorophyll values for the screen-house experiment one and two in this study gradually reduced as the water stress advanced. This corroborates studies by Bogale et al. (2011) that water deficit has tremendous effects on chlorophyll fluorescence and leaf gas exchange parameters. Thus, photosynthesis rates decreased with decreases in stomatal conductance. Therefore, the relative water content, chlorophyll contents, leaf wilting, survival, and recovery from drought are very good indices for Screen-house selection for seedling drought tolerance of large populations of inbred lines for drought evaluation in the field (Pungulani et al., 2013; Muchero et al., 2008; Singh et al., 1999c).
CONCLUSION
There were variations with respect to seedling drought tolerance for the 200 inbred lines using the parents as checks in this experiment. The inbred lines that performed well for recovery were 84, 406, 325, 223, 75, 186, 131, 20, 38, 230, 398 and 353, the susceptible ones were, 142, 78, 55, 57, 408, 255, 396, 116, 189, 255, and 28. The inbred lines that maintained a higher relative water content during stress imposition avoided drought and recovered better, were inbred lines 406, 325, 84, 230, 38 and 75, which recorded higher percentages of relative water contents as well as chlorophyll content.
The inbred lines whose proportion of survival ranged between a score of 100- 60% consistently for screen-house 1 and 2 experiments were selected as the potential tolerant lines. Whereas the ones that had poor survival and recovery after re-watering; whose recovery ranged between 0-40% were identified as potential susceptible inbred lines. The relative water content and chlorophyll for the potential seedling tolerant lines were in the range of 40-70% and 32-53%. The screen-house experiment was repeated to confirm the potential seedling tolerant and susceptible inbred lines that were subsequently selected for the field screening under managed drought conditions. Most of the potential susceptible inbred lines also had lower chlorophyll contents but the relative water content was most often within average (39 and 50%). This could be due to the environmental conditions prevailing at the time of the second experiment thus leading to the high proportions for survived seedlings across all the treatments for all the blocks. The use of physiological traits in scoring for seedling tolerance in this study has facilitated the classification of the inbred lines into drought tolerant and susceptible inbred lines. The genetic variability found for this morphological trait among these inbred lines in the screen-house study, suggest that opportunity exists for selecting superior genotypes under water limited conditions in the field.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Aliyu OM, Makinde BO (2016). Phenotypic analysis of seed yield and yield components in cowpea (Vigna unguiculata L., Walp). Plant Breeding and Biotechnology 4:252-261. |
|
|
Aref I, El Atta H, El Obeid M (2013). Effect of water stress on relative water and chlorophyll contents of Juniperus procera Hochst. ex Endlicher in Saudi Arabia. Life Science Journal 10:681-685. |
|
|
Batieno BJ, Danquah E, Tignegre J-B (2016). Application of marker-assisted backcrossing to improve cowpea (Vignaunguiculata L. Walp) for drought tolerance. Journal of Plant Breeding and Crop Science 8:273-286. |
|
|
Bogale A, Tesfaye K, Geleto T (2011). Morphological and physiological attributes associated to drought tolerance of Ethiopian durum wheat genotypes under water deficit condition. Journal of Biodiversity and Environmental Sciences 1:22-36. |
|
|
Clavel D, Drame NK, Roy-Macauley H (2005). Analysis of early responses to drought associated with field drought adaptation in four Sahelian groundnut (Arachis hypogaea L.) cultivars. Environmental and experimental Botany 54:219-230. |
|
|
Cornic G, Massacci A (1996). Leaf photosynthesis under drought stress. Photosynthesis and the Environment 5:347-366. |
|
|
Deblonde P, Ledent J-F (2000). Effects of moderate drought conditions on crop growth parameters and earliness of six potato cultivars under field conditions. Agronomie 20:595-608. |
|
|
Fan Y, Shabala S, Ma Y (2015) Using QTL mapping to investigate the relationships between abiotic stress tolerance (drought and salinity) and agronomic and physiological traits. BMC genomics 16:43. |
|
|
Fatokun CA, Boukar O, Muranaka S (2012) Evaluation of cowpea (Vigna unguiculata (L.) Walp.) germplasm lines for tolerance to drought. Plant Genetic Resources 10:171-176. |
|
|
Freire FFR, Ribeiro VQ, Rocha M de M (2012). Production, breeding and potential of cowpea crop in Brazil. Embrapa Meio-Norte-Documentos. |
|
|
Godfray HCJ, Beddington JR, Crute IR (2010). Food security: the challenge of feeding 9 billion people. Science 327:812-818. |
|
|
Gong H, Zhu X, Chen K (2005). Silicon alleviates oxidative damage of wheat plants in pots under drought. Plant Science 169:313-321. |
|
|
Khan MIR, Nazir F, Asgher M (2015). Selenium and sulfur influence ethylene formation and alleviate cadmium-induced oxidative stress by improving proline and glutathione production in wheat. Journal of Plant Physiology 173:9-18. |
|
|
Lawlor DW, Tezara W (2009). Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells: a critical evaluation of mechanisms and integration of processes. Annals of Botany 103:561-579. |
|
|
Mai-Kodomi Y, Singh BB, Myers O (1999). Two mechanisms of drought tolerance in cowpea. Indian Journal of Genetics and Plant Breeding 59:309-316. |
|
|
Markwell J, Osterman JC, Mitchell JL (1995). Calibration of the Minolta SPAD-502 leaf chlorophyll meter. Photosynthesis research 46:467-472. |
|
|
Muchero W, Ehlers JD, Roberts PA (2008) Seedling Stage Drought-Induced Phenotypes and Drought-Responsive Genes in Diverse Cowpea Genotypes. Crop Science 48:541. |
|
|
Oliveira RM de M, Ribeiro VQ, Lopes ÂC de A (2016). Diallel analysis in cowpea aiming at selection for extra-earliness. Crop Breeding and Applied Biotechnology 16:167-173. |
|
|
Olubunmi D (2015). Genetic Analysis of Drought Tolerance in Cowpea [Vigna Unguiculata (L.) Walp]. (Doctoral dissertation, University Of Ghana). |
|
|
Ortiz R, Ekanayake I, Mahalakshmi V (2002). Development of drought resistant and water stress tolerant crops through traditional breeding. Proceedings of the 8th JlRCAS International Symposia, Tsukuba, 27, 28 November, 2001 |
|
|
Parfitt J, Barthel M, Macnaughton S (2010). Food waste within food supply chains: quantification and potential for change to 2050. Philosophical Transactions of the Royal Society of London B: Biological Sciences 365:3065-3081. |
|
|
Pirzad A, Shakiba MR, Zehtab-Salmasi S (2011). Effect of water stress on leaf relative water content, chlorophyll, proline and soluble carbohydrates in Matricaria chamomilla L. Journal of Medicinal Plants Research 5:2483-2488. |
|
|
Pungulani LL, Millner JP, Williams WM, Banda M (2013). Improvement of leaf wilting scoring system in cowpea ('Vigna unguiculata'(L) Walp.): From qualitative scale to quantitative index. Australian Journal of Crop Science 7:1262. |
|
|
Santos J, Leitão-Correia F, Sousa MJ, Leão C (2016). Dietary Restriction and Nutrient Balance in Aging. Oxidative medicine and cellular longevity. Article ID 4010357. doi: 10.1155/2016/4010357 |
|
|
Singh B, Mai Kodomi Y, Terao T (1999a). Relative drought tolerance of major rainfed crops of the arid tropics. Indian Journal of Genetics 54:437-444. |
|
|
Singh B, Mai-Kodomi Y, Terao T (1999b). A simple screening method for drought tolerance in cowpea. The Indian Journal of Genetics and Plant Breeding 59:211-220. |
|
|
Singh B, Mai-Kodomi Y, Terao T (1999c). A simple screening method for drought tolerance in cowpea. The Indian Journal of Genetics and Plant Breeding 59:211-220. |
|
|
Soltys-Kalina D, Plich J, Strzelczyk-Żyta D, Śliwka J, Marczewski W (2016). The effect of drought stress on the leaf relative water content and tuber yield of a half-sib family of 'Katahdin'-derived potato cultivars. Breeding Science 66(2):328-331. |
|
|
Steidle NAJ, Lopes DC, Borges JJCF (2017). Assessment of Photosynthetic Pigment and Water Contents in Intact Sunflower Plants from Spectral Indices. Agriculture 7:8. |
|
|
Swain S, Munsi PS, Das S, Lenka PC (2017). Selection of the parental combinations through multivariate analysis in dessert banana genotypes for hybridization programme. International Journal of Agriculture, Environment and Biotechnology 10:73. |
|
|
Tomar SMS, Kumar GT (2004). Seedling survivability as a selection criterion for drought tolerance in wheat. Plant Breeding 123:392-394. |
|
|
Tuberosa R (2012). Phenotyping for drought tolerance of crops in the genomics era. Frontiers in physiology 3:347. |
|
|
Vasconcelos IM, Maia FMM, Farias DF (2010). Protein fractions, amino acid composition and antinutritional constituents of high-yielding cowpea cultivars. Journal of Food Composition and Analysis 23:54-60. |
|
|
Verbree DA, Singh BB, Payne WA (2015). Genetics and Heritability of Shoot Drought Tolerance in Cowpea Seedlings. Crop Science 55:146-153. |
|
|
Wallace J (2000). Increasing agricultural water use efficiency to meet future food production. Agriculture, ecosystems and environment 82:105-119. |
|
|
Yuan XK, Yang ZQ, Li YX (2016). Effects of different levels of water stress on leaf photosynthetic characteristics and antioxidant enzyme activities of greenhouse tomato. Photosynthetica 54:28-39. |
|
|
Zegaoui Z, Planchais S, Cabassa C (2017). Variation in relative water content, proline accumulation and stress gene expression in two cowpea landraces under drought. Journal of Plant Physiology 218:26-34. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


