Full Length Research Paper

ABSTRACT
Three pollination control tents (PCTs) made from novel nonwoven synthetic fabrics with more open pore structure (DWB10, DWB23 and DWB24) were compared with the standard DWB01 fabric for pollen proofing in sugar beet (Beta vulgaris L.) at the research station of Lion Seed Ltd Essex, UK in 2019. PCTs of 63.5 x 63.5 cm footprint accommodated single potted plants. A completely randomised trial with five replications including an open pollinated control was conducted using cytoplasmic male sterile line 1TM37. Analysis was computed for (a) full data and (b) excluding three DWB23 defective tents. Differences among treatments were non-significant for all morphological traits except for number of secondary branches in (a) only. There was thus no micro-climatic difference among treatments for morphological traits of the 1TM37 CMS line. Among the seed related traits, 1000-seed weight and 10-day germination (%) were significant between treatments in (a) but only 1000-seed weight in (b). The mean 1000-seed weight was significantly higher for the open control than all other PCT treatment means which did not differ significantly from zero. Therefore, all four fabrics of PCTs were equally pollen proof in preventing pollen contamination. It is concluded for the first time that mini-tents of these novel nonwoven fabrics, engineered for both larger pores for air permeability and fibre architecture to prevent pollen transmission, adequately eliminated cross-pollination while maintaining ambient environmental conditions and are effective for sugar beet breeding. The PCT technology may be equally usefully deployed in other traditional, commercial and fibre crops for hybrid seed production.
Key words: Sugar beet, nonwoven synthetic fabric, pollination control tent, male sterility.
INTRODUCTION
Male sterility (MS) is the result of non-functional pollen in plants (Chen and Liu, 2014). Sugar beet hybrid seed production uses cytoplasmic-genetic male sterility (McGarth and Panella, 2018) by involving three parental lines: a cytoplasmic male-sterile (MS or A-line) family, an O-type maintainer family (also called B-line) and a pollinator with restorer gene (R-line). Commercial hybrid seed production is performed in open fields, where the pollinator and MS family (F1 of A-line and O-line) are grown next to each other. Pollination occurs in the following year once the parental lines have overwintered and vernalised for transition from vegetative to reproductive phase. Reproduction of the MS family (A-line) is achieved through pollination by an O-type maintainer that is equivalent in the nuclear genome; the seed so produced on the male sterile plants is intended to faithfully reproduce male sterile plants of the A-line (Brown et al., 2014). The male sterile families in sugar beet are phenotypically true-breeding and near-homozygous for practical purposes with all plants looking alike and any phenotypic observable variation among plants of the family is regarded as environmental or non-genetic although some of this variation may be residual genetic. We exploit this feature of male sterile family 1TM37 in the present investigation.
Sugar beet (Beta vulgaris L.) is pollinated by wind and possibly by insects (Bodnar, 2010). According to Hecker (1988) the mean diameter of pollen of diploid (2x) strains of sugar beet was 20.8 µm (19.3 to 22.5 µm) and that of auto-tetraploid (4x) to be 25.9 µm (23.4 to 27.4 µm). Clifton-Brown et al. (2018) reported average size of sugar beet pollen to be ~20-25 µm. The pollen may be carried up to 1200 m (Darmency et al., 2009). The Animal and Plant Health Agency (2016) in its Technical Standard Supplements laid out 1000 m isolation distance from any pollen source of genus Beta for producing basic or certified seed. When breeding sugar beet and other crops with small pollen, breeders typically rely on distances for seed production or open ended poly-tunnels angled away from the prevailing wind. Other methods such as pollination control bags, isolation chambers with controlled conditions and tents may be used for breeding operations. However, all these methods have their own limitations and advantages.
Commonly, plant breeders create artificial isolations with pollination control bags (PCBs) made of various materials. Commonly used PCBs made of paper, cellulose or polyethylene are cheap but are easily damaged by birds, wind and bad weather. Further, transparent film PCBs create higher temperature in them especially in the hot season and may adversely affect the pollination outcome (Gitz et al., 2015; Scheffert et al., 2016, 2018, 2019). More recently, specially developed nonwoven synthetic materials have been used for PCBs for their greater strength against bird damage and inclement weather conditions of heavy rains and wind, greater air permeability, lower moisture absorption, re-usability and pollen proofing (PBS International, 2020a, b, c). Such bags have been shown to have advantage over the controls for greater seed harvest by Gitz et al. (2015), Schaffert et al. (2016, 2018, 2019) and Gaddameedi et al. (2017) in sorgum; Clifton-Brown et al. (2018) in sugar beet, wheat, Arabidopsis and Miscanthus; Hayes and Virk (2016) in Miscanthus; Vogel et al. (2014) and Adhikari et al. (2015) in grasses; and Bonneau et al. (2017) in oil palm. Anecdotal evidence suggests that certain nonwoven materials when used as pollination control in very hot climates can result in the plant getting too warm affecting plant health and pollen viability.
Therefore, finding more open nonwoven materials having greater air flow but retaining pollen proofing is important in increasing pollination performance. In this study, we uniquely and purposely included pollination control tents (PCTs) made from nonwoven synthetic fabrics designed to have greater pore sizes than the standard for air permeability, but a complex fibre arrangement to optimise pollen proofing ability in sugar beet, with the ultimate goal of using them in hybrid seed production and other breeding operations.
Comparing performance of stable cytoplasmic-genetic male sterile (MS) line in mini-isolation tents with an open control should provide tests for the following hypotheses:
1) The morphological plant traits of the MS line under mini-tents and in open control do not perform differently, H0. Significant variation indicates differential micro-environment within tents from the control and rejects H0.
2) The mean number of seeds set on MS plants in the tents should be zero; H0. In the event of seed set being significantly higher than zero there ought to be pollen contamination and null hypothesis stands rejected.
Usually, pollination bag materials have porosity smaller than the pollen size to avoid contamination (Hayes and Virk, 2016). However, in the present study, we use for the first time, specially designed single plant mini-tents made from nonwoven synthetic fabrics with greater strength and air permeability including pores larger than the sugar beet pollen to be controlled. As such this investigation lays a foundation for a new research area on pollination tents which has been sparingly investigated. It opens up new avenues for enhancing pollination performance in crop breeding. The major objectives were to: (i). Evaluate PCTs if they create within them a micro-climate different from the open control and (ii). Establish the pollen proofing ability of different fabrics with more open structures for use in sugar beet breeding.
MATERIALS AND METHODS
The germplasm used for the experiment was Lion Seeds family 1TM37. This is an established cytoplasmic-genetic male sterile line which has shown excellent performance in the past. There were five treatments of pollination control tents:
1. Open control with no cover. This treatment indicated the adequacy of pollen pressure in the field.
2. Currently used standard control at Lion Seeds as classic duraweb® or DWB01.
3. Three new nonwoven synthetic fabric tents: DWB10, DWB23 and DWB24.
The specific characteristics of new nonwoven fabrics specially obtained from PBS International, Scarborough, UK, in comparison to presently used DWB01 are given in Table 1. DWB01 is a standard nonwoven fabric used in sugar beet which has thin filtration layer. It is heat bonded making the fabric smooth and easy to working with. On the other hand, the new nonwoven fabrics are spun-bond and thicker with a more complex filtration layer (Table 1). This gives the fabrics greater strength. All new fabrics have similar mass per unit area but greater pore size that makes them more air permeable for better temperature and humidity control. Light transmission in the range of 350 to 800 nm was 4% higher for DWB23 and DWB24 than the standard (Table 1).

Five replicate plants were used in each treatment to cover the possibilities of failures from damage by accident or a plant dying and to provide enough degrees of freedom for a valid statistical analysis. The design used was a completely randomised design with all plants of all five treatments being randomised together. Mini-tents with dimension of 63.5 x 63.5 cm, 120 cm height plus a 20 cm skirt at the base were designed for the experiment by PBS International (Figure 1). Each mini-tent accommodated a single potted plant (Figure 1). Tents were placed 50 cm apart in a single line of 22.1 m length that ensured equal pollen pressure over the whole experiment.
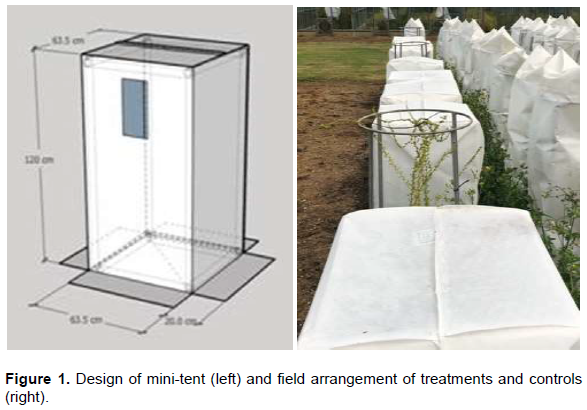
Single plants of male sterile line 1TM37 grown in pots were placed inside mini-PCT enclosures of various fabrics. Pollen pressure was generated by flowering sugar beet plants in adjacent poly-tunnels surrounding the experiment. To generate sufficient pollen pressure, the mini-tents were located in the down- wind direction from the pollen producing plants in the prevailing wind(s). Any seed observed on male sterile plants must result from the foreign pollen that might pass through the cover-fabric of the tent. The covers were adequately fixed to prevent pollen entry underneath the skirt and the frames adequately anchored to minimise the chance of wind damage. The frames were anchored to the ground on at least two sides and each side of the cover was anchored with one or more sandbags (Figures 1 and 2).
The plants were enclosed on 10th of June 2019 to ensure their isolation before flowering. The plants were harvested on 9th August 2019 and various data were collected.

Data collection
Length of primary stem was measured in centimetres and secondary branches counted. Cut weight of the whole plant was recorded in grams. Seeds were separated and weighed for each plant in grams and divided into those <2.8 mm ø (diameter) and >2.8 mm ø (Figure 3). The latter were taken as prospective seeds and weighed. These were divided into four replicates of 100 seeds each and weighed separately and then 1000-seed weight was derived. Each of these four replicates of 100 seeds per plant was sown for germination test. Final germination (%) over four replicates was computed after 10 days.

The amount of seed from an un-bagged plant can only be best assessed by considering the results from the controls because the amount varies from season to season. Sugar beet breeders usually expect at least 10 g with 75% germination – but much more is possible in good conditions. The germination certainly is an important aspect that needs to be taken into account. What is recorded as seed weight is really the mass of ‘seed sized material’ that was recovered by the sieving procedure. In many cases, aborted flowers will dry down into small seed shapes and are recovered but fail to germinate as they are not real, viable seed.
Implied number of seeds (IS)
Any seed-like material with >2.8 mm ø (diameter) was taken as probable seed and weighed together for each plant in grams (X).
Four samples of 100 seeds were taken and weighed, and weight for 1000 seeds was extrapolated from weight of four hundred seeds in grams (Y). The implied number of seeds (IS) was computed using X and Y:
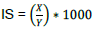
Implied total number of germinated seeds (ISG)
Number of seeds obtained from materials that looked like seed may be misleading. If it were a viable seed then it should germinate. Therefore, the number of implied seeds that could germinate by 10 days was computed to find out the actual number of seeds per plant as:

There were three cases of damage to pollination tent DWB23 on cage number 52, 62 and 70. Seams of 5 cm (at position 52) to 30 cm length (62 and 70 positions) were observed opened suspecting that they may allow pollen through. There was overgrowth of plant at position 62 that hit the top of cage forcing the seam open. In order to assess and exclude the effect of open seems on three plants of DWB23 treatment the statistical analysis was performed twice; once for the whole data set (a) and then by excluding (b) three defective cases of DWB23.
Statistical analysis of individual plants data and germination percent was performed following the analysis of variance technique described by Sokal and Rahlf (2011) using MINITAB17 statistical package. Standard errors of mean (SE) and lsd at 5% were computed as lsd = √2.SE.t value at 20 df (2.086). Fisher’s ‘Least significant difference (lsd)’ was used for pair-wise comparison of treatment means and significantly different means were labelled with different letters.
RESULTS AND DISCUSSION
In the context of our experiment seed on CMS 1TM37 under mini-tents can develop in two ways: (i) as a result of pollination by foreign pollen that passed through the tent fabrics and (ii) due to occasional failure of male sterility resulting in maternally produced selfed seeds. The occurrence of (ii) is very rare phenomenon and, in general, we do not expect seed set on CMS plants without pollination from extraneous pollen. The progeny of seeds from (i) will differ from the plants of mother family but that of (ii) will be phenotypically similar to the plants of the near-homozygous mother family within error limits. We anticipated confirming, if the seeds in tents resulted from outcrossing (i) or selfing (ii) through molecular marker studies by taking leaf samples of each of the trial plants before bagging for comparing with the progeny of seeds produced on CMS plants. However, this could not be accomplished and the simple criterion of the number of viable seed set under bagging was considered to be an indication of pollen contamination.
Analysis of data proceeded in two stages: (a) for the full data set and (b) for the data set excluding three defective cages for DWB23 and we shall refer to it in this way below.
Morphological plant traits
We do not expect significant variation for phenotypic traits among plants in the near- homozygous male sterile family 1TM37. The full data analysis (a) showed non-significant variation among treatments for cut plant weight and primary stem length but significant for number of secondary branches (Table 2). However, when analysis (b) was performed it turned out to be non-significant (Table 2). The examination of mean values for number of secondary branches shows that the significance among treatments in analysis (a) was due to the significantly lower performance of plants in the control than in PCT treatments, which were on par (Table 3). The number of secondary branches was on average 65% higher under cover than the control. It could be specific response of sugar beet to reduced light under cover. Wang and Feng (2004) reported typical leaf morphological responses to different light conditions in two species (Eupatorium adenophorum and Gynura sp.). At low light levels, plants enhanced light interception by means of increased biomass allocation to leaves and formation of large, thin leaves with high specific leaf area, leading to a high leaf area ratio. With a decrease in light intensity, plant of both species grew taller and produced more branches to intercept more light energy.


The mean number of secondary branches for DWB 23 in analysis (a) was 28.20 but for (b) it was 26.50. This reduction in number of branches reduced variation among treatments to a non-significant level in analysis (b). As seams of three mini-tents of DWB23 were open they encouraged plants to overgrow and push at the top. This perhaps is the physiological effect of shading. Plants growing in shade often tend to grow taller than they would grow outside under full sunlight. However, this is at the expense of energy and resources that could result in thinner main stem with fewer leaves or weaker roots and lower seed amount (Kniss and Schambow, 2016).
The mean values following analyses (a) and (b) showed (Table 3):
1. Mean number of secondary branches for the open control was significantly lower than the standard DWB01.
2. Mean number of secondary branches of the three new fabrics (DWB10, DWB23 and DWB24) were on par and non-significantly different from DWB01.
It can be concluded from the non-significant difference among mean values of PCT treatments for any morphological trait that the hypothesis of no micro-climatic difference among nonwoven synthetic fabrics and control is accepted. Thus, PCTs did not alter the plant environment to any significant effect on performance. However, Trammell et al. (2020) reported that the average temperature within tents was higher with lower average humidity than the open control but it produced a microclimate that gave 36% higher seed yield and disease free seeds. In the present study, because of the near-homozygous status of male sterile family all plants in it responded similarly to the changed environment under PCT covers.
Seed related traits
The sieving process for the seeds resulted in different type of seed sizes (Figure 3). It can be seen in Figure 3 that from one of the unprotected controls the grains were larger and better formed with the same counting board for size reference compared with those from DWB24. While about 13 g seed from control achieved 70% germination the same amount from DWB24 in four 100 seed replicates returned only 1.5% germination (Table 3). The mass that looked quite convincing as seed was mostly non-seed inert plant material that was not viable in DWB24.
Of special interest are seed-related traits since seed on plants in the open control resulted from cross pollination and that on plants in tents from pollen contamination or by parthenogenesis without contribution of pollen that could pass through the cover (Zhuzhzhalova et al., 2016). Of the six seed related traits the full analysis (a) showed significant variation between treatments for 1000-seed weight and 10-day germination (%) only (Table 2). However, in the reduced analysis (b) the significant variation was retained for 1000-seed weight only as the 10-day germination (%) became non-significant (Table 2). Therefore, we shall discuss the 1000-seed weight further.
Looking at mean values in Table 3 we find that 1000-seed weight for the control was significantly and 140% higher than mean of all PCT treatments together in (a). The high mean seed weight in control must arise from viable seeds resulting from cross pollination (Table 3). The comparison among treatment means showed that:
i. Open pollinated control’s mean seed weight was significantly higher than the standard DWB01.
ii. New fabrics (DWB10, DWB23 and DWB24) all had mean seed weight on a par between themselves and with the standard, DWB01.
Therefore, the significant variation between treatments in both analyses of variance largely arose from the deviant seed weight of the control. However, the smaller mean seed weights from all four PCT treatments were significantly higher than zero value when compared with the SE and LSD. This means our null hypothesis of no differences among treatments does not hold at this stage. The important question, however, arises: did the total seed mass representing seed weight contain viable seeds? This could be verified through germination test.
The implication of heavier and real viable seeds in open control is further reflected in its out-rightly higher 10-day germination (27%) and implied seed number at 10-day germination (340) in both analyses. On the other hand, germination for the PCT treatments in analysis (a) ranged from 0.10 to 3% with implied average seed number at 10-day germination ranging from 0.9 to 40.3. The highest value of 40.3 implied seeds belonged to DWB23 which might be the result of contamination in the defective tents. The analysis (b) excluding the defective tents showed germination range of 0.10 to 1.6% and implied seed number at 10-day germination ranging from 0.9 to 26. When we tested the mean values of germination (%) and implied seed number at 10-day germination in both analyses (a) and (b) there was no mean value that was significantly higher than zero (Table 3). All germination and implied seed number means were as good as zero.
The between plant variances within treatments were highly and positively correlated with treatment mean values for 10-day germination (r = 0.99; P<0.01) and implied seed number at 10-day germination (r = 0.99; P<0.01) but non-significantly correlated for number of secondary branches (r = -0.42; P>0.05) and 1000-seed weight (r = 0.66; P>0.05). The within variances of treatments did not differ significantly on a Bartlett’s Chi-square test (T) for number of secondary branches (T= 0.03 at 4 df; NS) and for 1000-seed weight (T= 0.54 at 4 df; NS). Apparently, there was no differential response of plants of MS line for these traits within tents and outside in the open.
Statistically there was zero mean seed set in all PCTs. It proved our hypothesis of no contamination by foreign pollen in all the pollination control tent fabrics and all fabrics of pollination tents were pollen proof. It may be recognised that maximum pore size of the PCT materials was greater than the average pollen size of ~20-25 µm in sugar beet but the structural arrangement of the fabric resulted in no contamination from outside pollen. Clifton-Brown et al. (2018) also observed no pollen contamination on plants covered with nonwoven fabric pollination control bags from externally placed red hypocotyl sugar beet variety. This is attributed to physical complexity of nonwoven spun-bound fabrics that have torturous path through the fibrous mesh ensuring that the entry of external pollen is restricted. Wang and Gong (2006) reported that the pore structure, pore size distribution, air permeability, and fabric area density of the 3D thermally bonded nonwoven filter samples consisted of multiple filtration layers of interconnected pores and tortuous pore paths through the fabric thickness. This torturous but purposefully effective filtration of pollen through larger pore size may not assure impermeable conditions yet it provides a trade-off in pollination performance (Figure 4). With the tested fabrics of PCTs we clearly find an acceptable filtering level of co-optimisation of pollen exclusion in the present experiment. However, unlike Clifton-Brown et al. (2018) who tested the nonwoven fabrics in glasshouse we have established their pollen proofing ability in field conditions although some of the previous studies suggested that maximum pore size be kept under the pollen size of the crop (Vogel et al., 2014).

The nonwoven fabric pollination mini-tent technology is relatively new for sugar beet. This study, for the first time, tested novel nonwoven fabrics in the form of pollination control tents in sugar beet breeding. None of the fabrics caused significant deviation in their micro-climate that could adversely affect biological performance of plants grown under their covers. However, reduced light under tents increased number of secondary branches (Wang and Feng, 2004). Further, all new fabrics with larger pores and hence more air-permeability were pollen proof. It therefore creates a possibility of optimisation of pore size and pollen filtering in different crops in future developments of fabrics. However, future studies in sugar beet should confirm the pollen contamination by studying the progeny of seed set on male sterile family to establish its maternal or cross-contamination origin. Further studies will confirm it by using a dominant morphological marker such as red hypocotyl pollen parent or molecular markers or simply studying the quantitative variation for different traits of the progeny of seeds in comparison with the mother parent.
The present study though carried out in one season at one location indicates that the tent technology can be usefully deployed for maintaining genetic integrity in various breeding operations such as attempting single crosses, generation advance of progenies, seed increase of selected progenies for multi-locational testing and providing multiple isolations simultaneously over limited space in a season. However, before full confidence is placed on the technology more research involving multi-environmental trials will be desirable. Further, exact categorisation of seed set on male sterile lines, however small in number, whether of maternal or contamination origin will establish pollen proofing ability of nonwoven fabrics beyond doubt. Therefore, experiments will continue to resolve these issues in the future. The simple sequence repeats (SSR) will be used to distinguish maternal and outcross seeds based on the amplified alleles of the progeny and the seed parent (Adhikari et al., 2015).
Recent advances in tent technology allow covering of larger areas with bigger tents and with modular extendable provisions (Trammell et al., 2020). Flexibility in size from mini tents to a modular design allows units of 1.5 x 3 m cubes (of 1.5 or 2 m tall) to be joined together to make larger structures as intended, for example 1.5 x 6 m, 3 x 3 m or bigger (PBS International, 2020c).
Flexibility in sizing the tent-covered area will go a long way in adjusting the protected area to breeders’ requirement in any season for any crop. Further developments must be directed towards stronger but lighter frames that are robust in bad weather and easy to transport. More research on new nonwoven synthetic fabric covers needs to be conducted for universal extension of technology. The present study has opened up new avenues of research on PCT technology for different crop plants and situations with possibilities of use in hybrid seed production in traditional, commercial and fibre crops. An in-depth economic analysis of technology needs to be conducted for its wider use in seed production and breeding.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGMENTS
The authors thankfully appreciate Ian Hansell and Naomi Reece-Ford from Lion Seeds Ltd, Essex CM9 6SN, UK for helping in the conduct of trial, and Rob Coy and Stuart Carter from PBS International, Scarborough YO11 3UP, UK for standardising and providing the single plant mini- pollination tents.
REFERENCES
|
Adhikari L, Anderson MP, Klatt A, Wu Y (2015). Testing the efficacy of a polyester bagging method for selfing switchgrass. BioEnergy Research 8:380-387. |
|
|
Animal and Plant Health Agency (2016). Technical Standard Supplements. Dec 2016, pp 1-136. |
|
|
Bodnar A (2010). Sugar beet biology. Biology fortified Inc. Retrieved on 6 May 2020. |
|
|
Bonneau L, Eli D, Vovola P, Virk DS (2017). Comparing pollination bag types for micro-environmental parameters influencing seed production in oil palm. Journal of Oil Palm Research 29(2):168-179. |
|
|
Brown J, Caligari P, Campos H (2014). Plant Breeding, 2nd edition. Wiley Blackwell, West Sussex, UK, P. 271. |
|
|
Chen L, Liu Y-G (2014). Male Sterility and Fertility Restoration in Crops. Annual Review of Plant Biology 65:579-606. |
|
|
Clifton-Brown JC, Senior H, Purdy SJ, Horsnell R, Lankamp B, MuEennekhoff A K, Virk D, Guillemois E, Chetty V, Cookson A, Girdwood S, Clifton-Brown G, Tan ML, Awty-Carroll D, Bentley AR (2018). Investigating the potential of novel nonwoven fabrics for efficient pollination control in plant breeding. PLoS ONE 13(9):1-21, e0204728. |
|
|
Darmency H, Klein EK, De Garanbé TG, Gouyon PH, Richard-Molard M, Muchembled C (2009). Pollen dispersal in sugar beet production fields. Theoretical and Applied Genetics 118(6):1083-92. |
|
|
Gaddameedi A, Kumar AA, Madhavrao PR, Virk DS, Senior H (2017). Evaluating the efficacy of synthetic fibre pollination control bags in Sorghum during the rainy season. International Journal of Plant Breeding and Genetics 11(1):39-54. |
|
|
Gitz DC, Baker JT, Xin Z, Burke JJ, Lascano RJ (2015). The microenvironment within and pollen transmission through polyethylene sorghum pollination bags. American Journal of Plant Science 6:265-274. |
|
|
Hayes C, Virk DS (2016). Assessing the relative efficacy of polyester pollination bags and crossing tents, and isolation chambers for seed harvest in Miscanthus crosses. International Journal of Plant Breeding and Genetics 10(2):79-90. |
|
|
Hecker RJ (1988). Pollen Characteristics of Diploid and Tetraploid Sugarbeet. Journal of Sugar Beet Research 25(1):55-62. |
|
|
Kniss A, Schambow T (2016). Shade avoidance. Sugarbeet Grower 55(4):4. |
|
|
McGarth JM, Panella L (2018). Sugar beet breeding. In: Irwin Goldman (Ed), Plant Breeding Reviews, Volume 42 Chapter 5, pp.167-218. John Wiley & Sons. Print ISBN: 9781119521310 |Online ISBN:9781119521358| |
|
|
PBS International (2020a). Intelligent design-why our bags are best. |
|
|
PBS International (2020b). Grasses research-why our bags are best. |
|
|
PBS International (2020c).Pollination Control just got Bigger! |
|
|
Schaffert RE, Virk DS, Senior H (2016). Comparing pollination control bag types for sorghum seed harvest. Journal of Plant Breeding and Crop Science 8(8):126-137. |
|
|
Schaffert RE, Virk DS, Senior H (2018). Are nonwoven synthetic pollination bags a better choice for sorghum breeding? Journal of Plant Breeding and Crop Science 10(3):58-68. |
|
|
Schaffert RE, Virk DS, Senior H (2019). Are Nonwoven Synthetic Pollination Bags a Better Choice for Sorghum Breeding? In Current Research in Agriculture and Horticulture Vol. 1 (Chapter 9):82-95. Print ISBN: 978-81-940613-7-3, eBook ISBN: 978-93-89246-27-8. |
|
|
Sokal RR, Rohlf FJ (2011). Biometry: The Principles and Practice of Statistics in Biological Research. 4th Edn., W.H. Freeman and Co., New York, ISBN-13: 978-0-7167-8604-7, Pages: 937. |
|
|
Trammell M, Pittman DG, Virk DS, Senior H (2020). Assessing the effectiveness of nonwoven fabric pollination tents for improved grass breeding. Journal of Plant Breeding and Crop Science 12(3):200-218. |
|
|
Vogel KP, Sarath G, Mitchell RB (2014). Micromesh fabric pollination bags for switchgrass. Crop Science 54:1621-1623. |
|
|
Wang J, Feng Y (2004). The effect of light intensity on biomass allocation, leaf morphology and relative growth rate of two invasive plants. Acta Phytoecological Sinica 28(6):781-786. |
|
|
Wang WH, Gong RH (2006). Thermally Bonded Nonwoven Filters Composed of Bi-Component Polypropylene/Polyester Fiber. II. Relationships between Fabric Area Density, Air Permeability, and Pore Size Distribution. Journal of Applied Polymer Science 102:2264-2275. |
|
|
Zhuzhzhalova TP, Podvigina OA, Znamenskaya VV, Vasil'chenko EN, Karpechenko NA, Zemlyanukhina OA (2016). Sugar beet (Beta vulgaris L.) haploid parthenogenesis in vitro: Factors and diagnostic characters. Sel'skokhozyaistvennaya Biologiya [Agricultural Biology] 51(5):636-644. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


