Full Length Research Paper

ABSTRACT
Taro (Colocasia esculenta (L.) Schott) is a food crop mainly produced for its nutrient-rich corms. This study aims to characterize the nutritional and antioxidant potential of fifty-nine genotypes of taro to identify the most interesting chemotypes. The traits evaluated were: starch, proteins, soluble sugars, total phenolics, flavonoids, and antioxidant capacity, assessed by Ferric reducing antioxidant power (FRAP) and 2,2-diphenyl-1-picrylhydrazyl (DPPH) methods. Results indicate that taro corms are a good source of nutritional and bioactive compounds. Also, significant variations were recorded between genotypes for the different parameters evaluated. Total phenolics content varied from 4.37± 0.02 to 115.21 ± 8.24 mg GAE/100 g dw, flavonoids from 0.25 ± 0 to 1.47 ± 0.07 mg quercetin equivalent (QE)/100 g dw, starch content from 0.31 ± 0.17 to 17.56 ± 0.27 mg/100 mg dw, proteins from 0.03 ± 0.03 to 0.80 ± 0.02 mg/100 mg dw and soluble sugars from 0.02 ± 0 to 8.10 ± 0.38 mg/100 mg dw. Antioxidant activities ranged from 39.20 ± 1.74 to 98.67 ± 1.62% for the ability to reduce DPPH and from 0.06 ± 0.03 to 12.16 ± 0.72 mg AAE/100 g dw for the ability to reduce iron (FRAP). These variations have allowed us to distinguish four chemotypes.
Key words: Colocasia esculenta, taro, nutritional compounds, bioactive compounds, breeding, Burkina Faso.
INTRODUCTION
Taro [Colocasia esculenta (L.) Schott] is widely grown in the tropics and subtropics for its edible corms and leaves (Huang et al., 2010; Chaïr et al., 2016). It is an important staple for strengthening food security for poor rural populations in Africa, Asia, and the Pacific Islands (Gebre et al., 2015; Ganança et al., 2017). The average global production is 10.1 million tonnes on 1.4 million hectares (FAOSTAT, 2016). The corms, which are the main edible parts with high economic potential, have a nutritional value similar to sweet potato. Indeed, corms are an excellent source of starch, sugars, proteins, fibers, and minerals (Krishnapriya and Suganthi, 2017). Taro is a source of thiamin, riboflavin, iron, phosphorus, zinc, and a very good source of vitamin B6, vitamin C, niacin, potassium, copper, and manganese (Quach et al., 2003). Also, the plant has a range of biologically active compounds such as flavonoids, glycosides, and other micronutrients (Lebot et al., 2017). Taro corms contain phenolic compounds such as tannins, flavonoids and saponins (Alcantara et al., 2013). With their high antioxidant potential, these compounds have protective properties for the body against some chronic diseases and other health-promoting properties. Epidemiological studies have shown that consuming food rich in phenolic antioxidant compounds protects the body against chronic non-communicable diseases such as cancer, aging, diabetes, and cardiovascular diseases (Diego et al., 2017). These phenolic compounds act as radical scavengers, lowering agents, and chelators of metal ions (iron, for example). The nutritional potential and bioactive properties of taro can vary considerably depending on the cultivar. Several studies carried out on taro germplasm collections (Lebot and Legendre, 2015; Lebot et al., 2017)and other root and tuber crops, reported variability in phytochemical compositions and biological properties depending on genotype (Champagne et al., 2011; Senanayake et al., 2013). Thus, some varieties would present better content of nutritional and bioactive compounds. This genetic diversity could be interesting to initiate selection and varietal improvement through breeding to offer to consumers, varieties with the best potentials to strengthen human health (Alcantara et al., 2013).
In Burkina Faso, in addition to the long-cultivated endogenous taro cultivars, several (50) exogenous varieties were introduced in favor of ACPCCCC EU funded project (Adapting Clonally Propagated Crops to Climatic and Commercial Changes). All these varieties (endogenous and exogenous) were characterized agro-morphologically (Traoré et al., 2013; Ouédraogo et al., 2018)and molecularly (Traoré, 2014). However, the characterization of their nutritional and bioactive potentials is necessary to identify the best genotypes for breeding and varietal improvement programmes. This study, which aimed to characterize the nutrients and phytochemical compounds contents of taro genotypes corms, is an essential contribution to the conservation, management, promotion and efficient use of taro genetic resources in Burkina Faso which could be used as a reference material for West Africa.
MATERIALS AND METHODS
Plant
The plant materials are composed of fifty-nine genotypes, including twenty-nine exotic varieties and thirty local accessions of taro. The twenty-nine exotic varieties were received as in vitro-plants through an international EU funded project. They were chosen from the taro genetic resources held by the Secretariat of the Pacific Community (PCS) in Fiji, South Pacific. The thirty local accessions were collected in the provinces of Comoé and Kénédougou, respectively, in Cascades and Hauts Bassins regions of Burkina Faso. These two provinces are the largest taro producing regions in Burkina Faso (Traoré, 2014).
Production of corms
The analyzed corms were harvested at the Gampela experimental station. Genotypes were established following a fully randomized Fisher block design. After being fertilized with organic fertilizer, and the plants were transplanted into seeds pits dug in soil plowed by a tractor.
The cubic-shaped pits with a 30 cm ridge were filled to 2/3 of their volume with compost mixed with the organic fertilizer fifteen days after planting. Watering was provided by rainwater and supplemented by borehole watering during the dry season. Weeding was carried out by hand whenever necessary.
Samples collection
Harvesting was done at maturity for each genotype as determined by the appearance of signs such as yellowing and wilting of the aerial parts, petioles, and laminas. Each sample was a mixture of corms from three blocks. The newly collected taro corms were carefully washed with running tap water and peeled. The thickness of the peelings was about 5 mm. The peeled corms were then cut into pieces (or chips) between 3 and 5 mm thick. The chips were dried at 70°C for 24 h in a PROLABO oven of the JOUAN brand, model EB170 (Château-Gontier, France). The dried corms were ground into a flour using a blender and mill (Binatone brand, model BLG-460, China).
Nutritional and nutraceutical potentials of corms
Proteins content
Taro corm flours of each genotype (500 mg) were homogenized in 5 ml of NaCl 0.1 M and centrifuged at 4500 rpm for 15 min. After centrifugation, the supernatant was used to determine the total proteins content. The total proteins content were determined according to the method described by Bradford (1976) using bovine serum albumin (BSA) as standard, and the content was expressed as mg/100 mg of dry flour weight (dw).
Soluble sugars content
Taro flours of each genotype (500 mg) were homogenized in 5 ml of 80% ethanol and boiled for 30 min. After cooling, the solution was centrifuged at 4000 rpm for 10 min. The supernatant was used to determine the soluble sugars content. Total soluble sugars were evaluated using the phenol-acid sulfuric method as described by Sombié et al. (2019), and the absorbance was read at 490 nm. The total sugar content was expressed as mg glucose equivalent/100 g of corms.
Starch content
The starch content of corms was determined according to the method described by Jarvis and Walker (1993). 100 mg of taro corm flour were homogenized in 5 ml of 1 N KOH at the ambient temperature then neutralized with 5 ml of 1 N HCl. The mixture was put in boiling in a bain-marie for 15 min, and the volume was adjusted at 10 ml with distilled water. After cooling, the solution was centrifuged, and the supernatant was used for the dosage of starch. The starch content of corms was expressed as mg of starch/100 mg of flour.
Content of bioactive compounds and antioxidant activities
Extraction method
The flour of corms of each genotype (0.5 g) has been extracted with 10 ml of acetone: water (80: 20, v/v). The mixing was put in agitation for 24 h and centrifuged at 4500 rpm for 15 min. The supernatant was used to quantify total phenolics, total flavonoids and assess the antioxidant activities.
Total phenolics content
Total phenolics content of corms was determined at 760 nm according to the method described by Singleton et al. (1999) by using gallic acid as standard. The content in total phenolics was expressed in mg of gallic acid equivalent per 100 g of corm flour (mg GAE/100 g dw).
Total flavonoids content
The total flavonoids content of corms extracts was determined at 415 nm using the method described by Arvouet-Grand et al. (1994). The total flavonoids content was determined on a quercetin calibration curve and expressed as mg of quercetin equivalent (QE) per 100 g of corm flour (mg QE/100 g dw).
Antioxidant activities
Ferric reducing antioxidant power (FRAP) assay
The capacity of taro corms extracts to reduce the iron (III) to iron (II) was measured at 700 nm according to the procedure described by Sombié et al. (2011). Iron (III) reducing activity has been determined in mg of ascorbic acid equivalent for 100 g of weight of corm flour (mg AAE/100 g dw).
DPPH radical scavenging activity
The capacity of taro corms to trap the radical 2,2-diphenyl-1-picrylhydrazyl (DPPH) was assessed at 517 nm as described by Sombié et al. (2011). The activity of taro extracts was expressed in the percentage of inhibition.
Physical parameters
Corm flesh color (CFC) was recorded to establish the relation between the different color modalities and the studied parameters. Also, the mass of the main corm (MPC) was measured to include this variable in the characterization of chemotypes. A Mettler Toledo digital electronic balance, model PL3001-S (Greifensee, Switzerland) was used for the weighing.
Statistical analyses
The data were entered, organized in an Excel sheet and analyzed using XL-STAT software. The analysis of variance (ANOVA), at the 5% threshold was performed to determine the most discriminant traits. Tukey test at the 5% threshold was used to compare the means. Pearson correlation test was used to study the relations between quantitative characters. The principal component analysis (PCA) was performed to determine the associations between the studied variables by integrating additional data, which are the mass of main corm (MPC) and the corm flesh color (CFC). The hierarchical ascendant clustering (HAC) was conducted using the Euclidean distance and the method of aggregation of Ward to define the structuration of the variability of the collection. The organized groups for the HAC were characterized by the discriminating factorial analysis (DFA).
RESULTS
Nutritional and nutraceutical characteristics
The coefficient of variation of nutritional and nutraceutical characteristics of the studied genotypes of taro are shown in Table 1. These results confirm significant variation between genotypes for all the quantitative studied traits. Total phenolic content varied from 4.37 ± 0.02 to 115.21 ± 8.24 mg GAE/100 g dw. The total flavonoids content varied from 0.25 ± 0 to 1.47 ± 0.07 mg QE/100 g dw. The primary metabolites quantification gave an average of 0.36 ± 0.20 mg/100 mg dw for proteins, 1.65 ± 1.65 mg/100 mg dw for soluble sugars; 12.96 ± 3.65 mg/100 mg dw for starch. The coefficient of variation are higher than 30% for the traits of polyphenols content (115.85%), proteins content (55.36%) and the antioxidant activity by the FRAP method (87.95%).The Appendix shows the average comparisons of Tukey’s test of all studied parameters of the genotypes collection.
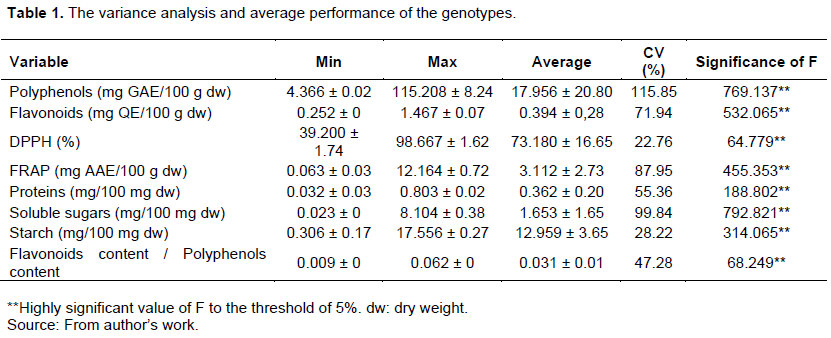
Antioxidant activities
The different genotypes showed generally good antioxidant activities. Thus, their ability to reduce DPPH is assessed on average at 73.18 ± 16.65%, with a maximum value reaching 98.67 ± 1.62%. For the capacity to reduce iron, we note an average of 3.11 ± 2.73 mg AAE/100 g and a maximum value of 12.16 ± 0.72 mg AAE/100 g.
Relations between traits
Pearson correlation matrix (Table 2) showed significant correlations between several studied traits. The content in polyphenols is strongly and positively correlated with the content in flavonoids (r = 0.883**), soluble sugars (r = 0.681**), as well with the ferric reduction ability FRAP (r = 0.625**). Its correlation with the DPPH free radical scavenging capacity is positive and medium (r = 0.545**). However, it is negatively and weakly correlated with protein content (r = 0.378**) but moderately correlated with the mass of the main corm (r = 0.511**). Similarly, the correlation of the yellow color of the corm flesh is positively and strongly correlated with the flavonoids content (r = 0.642**), positively correlated with the polyphenols content (r = 0.402**) and with the reduction power of iron FRAP (r = 0.473**). For the white color of the corm flesh, it is negatively and weakly correlated to the flavonoids content (r = 0.357**) and to the antioxidants by the DPPH method (r = -0.309*) and FRAP method (r = -0.277*).
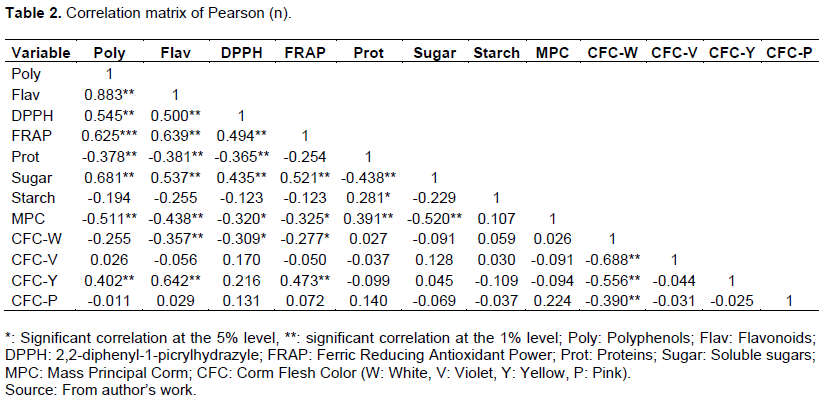
Thus, the different correlations observed justify the projection of the variables in the circle of the principal component analysis. The first factorial plane (Figure 1), with total inertia of 67.66%, made up of the F1 and F2 axes, is sufficient to reflect the greatest part of the variation. The F1 axis of inertia is 52.96% positively associated on one hand with the variables polyphenol, flavonoid and soluble sugar contents, and the degree of anti-oxidant activities and the other hand, negatively associated with the protein content and the corm mass. The F2 axis of inertia with 14.71% is positively associated with the starch content.
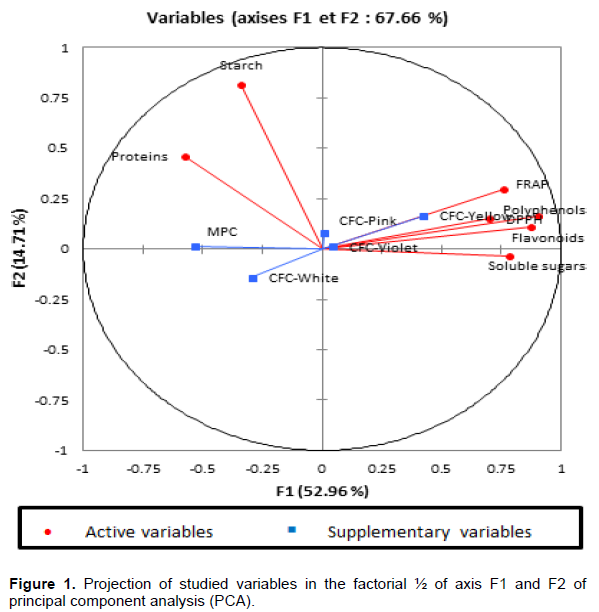
Structuration of the diversity
The ascending hierarchical clustering (AHC) using the Euclidean distance and the method of aggregation of Ward resulted in a dendrogram identified at a level of truncation level of about 30, four groups of different physico-chemical characteristics (Figure 2). Group 4 comprises four (4) exotic genotypes from the SPC collection and group 2 is composed of six (6) genotypes from Burkina Faso.
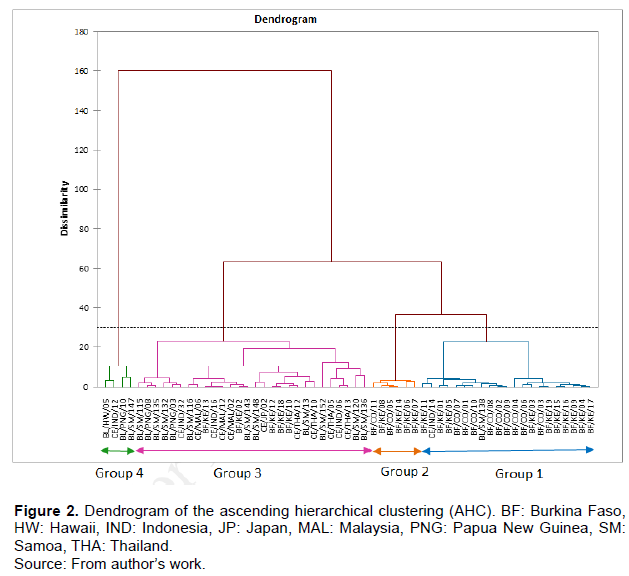
Group 1 with twenty-one (21) genotypes has only two (2) exotic genotypes, and group 3 has a total of twenty-five (25) genotypes which contain only five (5) local genotypes.
Characterization of groups from (HAC)
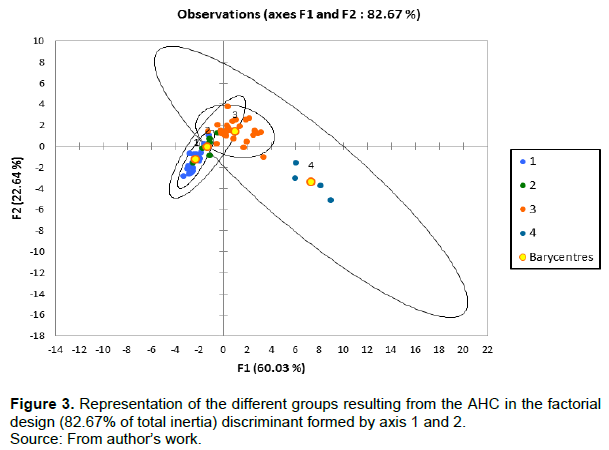
The factorial analysis of the studied traits of taro genotypes according to axis 1 and 2 (Figure 3) with 82.6% total inertia allows to characterize the different groups. Axis 1 with 60.03% of inertia is positively associated with group 4 and negatively with groups 1 and 2. At the same time, axis 2 with 22.64% of inertia is positively associated with group 3. Then, the different groups according to their distribution in factorial design ½ and the defining axis of PCA correlation circle are characterized as follows:
(1) Group 1 comprises genotypes with low polyphenols, flavonoid, and soluble sugars contents and low antioxidant activity. However, these genotypes showed the highest protein and starch contents and the highest weight of corm. The genotypes of this group have white corm flesh.
(2) Group 2 is composed of genotypes with a low content in starch. This group is close to group 1 in terms of polyphenols, flavonoids, and soluble sugars contents and their antioxidants' importance. The corm flesh color of this group of genotypes is also white.
(3) Group 3 is the group with the highest content in starch. In this group, the contents of polyphenols, flavonoids and antioxidants activities are lower than in group 4 but are much higher than in groups 1 and 2. All modalities of the variable color of the corm flesh are present in this group.
(4) Group 4 has the highest polyphenol contents, flavonoids, soluble sugars, and the highest antioxidants activities. Among its genotypes, the variety BL/PNG/10 which is distinguished by the yellow color of the corm flesh has the highest flavonoid content (1.467 ± 0.07 mg EQ/100 g dw), while the corm flesh of the others is white. The group also has the lowest content in proteins and the lowest mass of the main corm.
Groups 1 and 2 are composed of local accessions from Burkina Faso. They presented similar characteristics by considering their low content in secondary metabolites. Groups 1 and 2 contrast groups 3 and 4, which have a high content in secondary metabolites.
DISCUSSION
This study focused on the characterization of the nutritional, nutraceutical and antioxidant potentials of taro genotypes from Burkina Faso germplasm collection. Taro is a nutritious plant widely distributed in the world. This study showed that the contents in proteins, soluble sugars and starch varied according to the variety. The recorded values are similar to those reported in other studies (Krishnapriya and Suganthi, 2017). These values demonstrate that some accessions are an important source of proteins, sugars and starch. For comparison, the contents in proteins and the starch of sweet potato (Ipomoea batatas Lam.) root tubers are between 1.2 to 3.3 mg/100 mg and 33.7 to 64.1 mg/100 mg of dry matter (Senanayake et al., 2013). As for yams, the content in proteins varies from 1 to 1.8 mg/100 mg of dry matter (Ike, 1972).
Total phenolics and flavonoids contents also varied respectively according to the variety to 4.37 ± 0.02 to 115.21 ± 8.24 mg GAE/100 g of dry matter and from 0.25 ± 0 to 1.47 ± 0.07 mg QE/100 g of dry matter. These contents are close to those reported in other studies. Phenolics content of taro corms of 187 ± 53 mg GAE/100 g have been reported (Simsek and El, 2015). Similarly, contents ranging from 12 mg GAE/100 g for white-fleshed taro to 20 mg GAE/100 g for yellow-fleshed corms are noted (Lako et al., 2007). Overall, phenolics and flavonoids contents of the taro corms place this food plant among the richest in antioxidant compounds.
This study indicates that variability in studied parameters of the taro germplasm depends on the genotypes. Several studies on the quantification of phenolic compounds in taro corms have concluded that there is variability in phenolic compounds content between corms of different genotypes due to soil and climatic conditions of the growing area of the culture, sample processing, extraction conditions but also genetic factors (Champagne et al., 2010; Simsek and El, 2015). However, in our study, the influence of environmental factors, sample processing and extractions conditions on the variability of contents seems less plausible as genotypes were grown under the same pedoclimatic conditions within a fully randomized complete block design and the processing and the extraction were also carried out under the same conditions. Therefore, the observed variability in chemical composition seems to have a genetic determinism. In this regard, the four groups identified with the Hierarchical Ascending Clustering correspond to four distinct chemotypes. Group 4 is composed of exotic genotypes (BL/HW/05; BL/PNG/10; BL/SM/147; CE/IND/12) and is characterized by high phenolics and flavonoids contents and by the highest antioxidant activities. This chemotype is a material of choice for the production of taro corms beneficial for human health. Several recent studies emphasize the therapeutic proprieties of phenolic compounds, which are reported to be antiallergic, anti-inflammatory, anti-bacterial, antithrombotics, and vasodilators (Singh et al., 2014; Diego et al., 2017; Yildirim et al., 2017).
The study also showed high correlations between phenolics and flavonoids contents and the antioxidant activities (DPPH, FRAP). Similar results were reported by other authors on seeds of Pisum sativum L. (Zhao et al., 2020). The phenolic compounds would be responsible for the antioxidant properties of the extracts. The known benefits properties of phenolic compounds such as anti-inflammatory effects, antiallergic, hepatoprotective, anti-viral and antioxidant activities coincide with taro physiological properties (Diego et al., 2017). Therefore, taro is more than a food plant that can contribute to the fight against hunger and poverty in developing countries (Traoré et al., 2013) as it is also a food plant with very interesting antioxidant properties. High positive correlations were noted between the yellow color of the corm flesh and flavonoids content. Flavones, one of the main categories of flavonoids, are responsible for the yellow color of taro corms flesh and fibers (Lebot et al., 2015). These compounds exhibit antioxidant properties confirmed by the high positive correlations between their contents and antioxidant activities of extracts. The results pointed out that the difference in connection of the content in flavonoids on the content of total polyphenols of extracts is highly significant. Similar results have been reported (Takebayashi et al., 2013). However, a recent study mentioned a stable relationship of about a quarter (Simsek and El, 2015). These differences can be explained by the pigmentation of some corms, richer in flavonoids than other white-fleshed accessions.
A high positive correlation was observed between the total phenolics content and soluble sugars content. This later revelation might not worry the consumers who would fear the consuming of aliments rich in sugars because the highest sugars content has been estimated at 8.104 ± 0.38% of dry matter. Taro has a very low glycemic index as reported by Simsek and El (2015). The high positive correlations between the variables could be explained by genetic linkage reasons or a pleiotropic effect. Two situations arise for plant breeders: the possibility of simultaneous improvement of some traits and the difficulty of separating others. Conversely, it will be easier to select varieties with specific characteristics when the correlations are negative.
In the case of the present study, the strong negative correlations between the phenolics and proteins contents allowed the structuration of the variability to distinguish the groups 1 and 4 with opposite characteristics. It would be important to consider the improvement of varieties that combine these two opposite features and yet contribute to good nutrition.
CONCLUSION
The assessment of the nutritional value of corms of genotypes from the Burkina Faso germplasm collection confirms the tremendous nutritional potential of taro corms, as already reported by previous studies. However, the high variability of the different traits measured in the present study, allowed us to distinguish four distinct groups whose characteristics allow a specific use. The accessions of group 4 are distinguished from the others by the high content of phenolic compounds and an important antioxidant activity of their corm extracts.
These accessions might be promoted and valorized as functional foods. Likewise, it would be important that elite varieties are created through hybridization and phenotypic recurrent selection that will accumulate the characteristics desired by consumers. The adoption of markers assisted selection could also ease the process.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Alcantara RM, Hurtada WA, Dizon EI (2013). The Nutritional Value and Phytochemical Components of Taro [Colocasia esculenta (L.) Schott] Powder and its Selected Processed Foods. Journal of Nutrition and Food Sciences 3(3). |
|
|
Champagne A, Bernillon S, Moing A, Rolin D, Legendre L, Lebot V (2010). Carotenoid profiling of tropical root crop chemotypes from Vanuatu, South Pacific. Journal of Food Composition and Analysis 23:763-771 |
|
|
Champagne A, Hilbert G, Legendre L, Lebot V (2011). Diversity of anthocyanins and other phenolic compounds among tropical root crops from Vanuatu, South Pacific. Journal of Food Composition and Analysis 24(3):315-325. |
|
|
Arvouet-Grand A, Vennat B, Pourrat A, Legret P (1994). Standardisation d'un extrait de propolis et identification des principaux constituants. Journal de Pharmacie de Belgique Catalogue en ligne 49:462-468. |
|
|
Bradford MM (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry 72(1-2):248-254. |
|
|
Chaïr H, Traoré RE, Duval MF, Rivallan R, Mukherjee A, Aboagye LM (2016). Genetic Diversification and Dispersal of Taro (Colocasiaesculenta (L.)Schott). Plose One 11:0157712. |
|
|
Diego dSB, Cyntia SdF, Laidson PG, Davi dS, Anna CNTFC, Patricia RP, Eduardo MDA, Vania MFP (2017). Polyphenols from root, tubercles, and grains cropped in Brazil: chemical and nutritional characterization and their effects on human health and diseases. Nutrients 9:1044. |
|
|
FAOSTAT (2016). |
|
|
Ganança JFT, Freitas JGR, Nóbrega HGM, Rodrigues V, Antunes G, Gouveia CSS, Rodrigues M, Cha?r H, Pinheiro de Carvalho MÂA, Lebot V (2017). Screening for Drought Tolerance in Thirty Three Taro Cultivars. Notulae Botanicae Horti Agrobotanica Cluj-Napoca 46(1):65-74. |
|
|
Gebre A, Tesfaye B, Kassahun BM (2015). Effect of corm size and plant population density on corm yield of Taro (Colocasia Esculenta L.). International Journal of Advanced Biological and Biomedical Research 3(4):405-412. |
|
|
Huang CC, Lai P, Chen IH, Liu YF, Wang CCR (2010). Effects of mucilage on the thermal and pasting properties of yam, taro, and sweet potato starches. LWT - Food Science and Technology 43(6):849-855. |
|
|
Krishnapriya TV, Suganthi A (2017). Biochemical and phytochemical analysis of Colocasia esculenta ( L .) Schott tubers. International Journal of Research in Pharmacy and Pharmaceutical Sciences 2(3):21-25. |
|
|
Ike OL (1972). Yam - A Valuable Source of Food and Drugs. World Review of Nutrition and Dietetics 15:156-184. |
|
|
Jarvis CE, Walker JRL (1993). Simultaneous, Rapid, Spectrophotometric. Determination of Total Starch, Amylose, and Amylopectin. Journal of the Science of Food and Agriculture 63:53-51 |
|
|
Lako J, Trenerry VC, Rochfort S (2007). Routine analytical methods for use in South Pacific regional laboratories for determining naturally occurring antioxidants in food. International Food Research Journal 15(3):313-323 |
|
|
Lebot V, Lawac F, Michalet S, Legendre L (2017). Characterization of taro [Colocasia esculenta (L.) Schott] germplasm for improved flavonoid composition and content. Plant Genetic Resources: Characterisation and Utilisation 15(3):260-268. |
|
|
Lebot V, Legendre L (2015). HPTLC screening of taro hybrids (Colocasia esculenta (L.) Schott) with high flavonoids and antioxidant contents. Plant Breeding 134(1):129-134. |
|
|
Ouédraogo N, Traoré RE, Kando PB, Sawadogo M, Zongo JD (2018). Agro-morphological diversity of exotic taro varieties ( Colocasia esculenta L . Schott ) introduced in Burkina Faso. Journal of Experimental Biology and Agricultural Sciences 6(2320):370-385. |
|
|
Quach ML, Melton LD, Harris PJ, Burdon JN, Smith BG (2003). Cell wall compositions of raw and cooked corms of taro (Colocasia esculenta). Journal of the Science of Food and Agriculture 81:311-318. |
|
|
Senanayake S, Gunaratne A, Ranaweera KKDS, Bamunuarachchi A (2013). Physicochemical properties of five cultivars of sweet potato (Ipomea batatas Lam) roots grown in Sri Lanka. Tropical Agriculture 90(2):96-105. |
|
|
Simsek S, El SN (2015). In vitro starch digestibility, estimated glycemic index, and antioxidant potential of taro (Colocasia esculenta L . Schott) corm. Food Chemistry 168:257-261. |
|
|
Singh M, Kaur M, Silakari O (2014). Flavones:An Important Scaffold for Medicinal Chemistry. European Journal of Medicinal Chemistry, III:10.1016/j.ejmech.2014.07.013. |
|
|
Singleton VL, Orthofer R, Lamuela-Raventos RM (1999). Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-ciocalteu, reagent. Methods Enzymology 299:152-178. |
|
|
Sombié PAED, Hilou A, Mounier C, Coulibaly A, Kiendrebeogo M, Millogo JF, Nacoulma OG (2011). Antioxidant and anti-inflammatory activities from galls of Guiera senegalensis J.F. GMEL (Combretaceae). Research Journal of Medicinal Plant, 5:448-461. |
|
|
Sombié PAED, Sama H, Sidibé H, Kiendrébéogo M (2019). Effect of Organic (Jatropha Cake) and NPK Fertilizers on Improving Biochemical Components and Antioxidant Properties of Five Cowpea (Vigna unguiculata L. Walp.) Genotypes. Journal of Agricultural Science 11(10):48-62. |
|
|
Takebayashi J, Oki T, Watanabe J, Yamasaki K, Chen J, Sato-furukawa M, Tsubota-Utsugi M, Taku K, Goto K, Matsumoto T, Ishimi Y (2013). Analysis of Hydrophilic antioxidant capacities of vegetables and fruits commonly consumed in Japan and estimated average daily intake of hydrophilic antioxidants from these foods. Journal of Food Composition and Analysis 29(1):25-31. |
|
|
Traoré ER, Nanema RK, Bationo-Kando P, Sawadogo M, Nebié B, Zongo JD (2013). Variation agromorphologique dans une collection de taro (Colocasia esculenta ( L .) Schott ) adapté aux conditions de culture pluviale au Burkina Faso. International Journal of Biological and Chemical Sciences 7(4):1490-1502. |
|
|
Traoré, RE (2014). Etude de la diversité du taro (Colocasia esculenta (L.) Schott):cas d'une collection du Burkina Faso et d'une collection internationale. Thèse de Doctorat Unique, UFR/SVT, Université de Ouagadougou 161 p. |
|
|
Yildirim AB, Guner B, Karakas FP, Turker AU (2017). Evaluation of antibacterial, antitumor, antioxidant activities and phenolic constituents of field-grown and in vitro-grown Lysimachia vulgaris L.. African Journal of Traditional, Complementary, and Alternative Medicines 4(2):177-187. |
|
|
Zhao T, Su W, Qin Y, Wang L, Kang Y. 2020. Phenotypic diversity of pea (Pisum sativum L.) varieties and the polyphenols, flavonoids, and antioxidant activity of their seeds. Biology ISSNe 1678-4596. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


