Full Length Research Paper

ABSTRACT
Chickpea is one of the main annual crops in Ethiopia both in terms of its total cultivated area of pulses and its role in direct human consumption. The major problem to increase pulse production is the loss of seed viability and seeds damage from insect infestation particularly Adzuki bean beetle. Research works conducted on diversity study on chickpea varieties grown under different soil fertility levels were scanty. To fill such information gap, hundred chickpea genotypes that were managed under different soil fertility levels for Adzuki bean beetle resistance were screened under laboratory condition at Holetta and Debre Zeit. Cluster analysis was used to group the genotypes into five clusters with and without application of rhizobium and phosphorus, but into six clusters with the application of both rhizobium and phosphorus. Inter-cluster D2 average values of 11.50 (between clusters C4 and C5) to 62.94 (between clusters C2 and C5), 14.85 (between clusters C1 and C2) to 72.12 (between clusters C3 and C5) and 8.61 (between clusters C2 and C3) to 166.25 (between clusters C2 and C6) were found when genotypes grown under neither rhizobium nor phosphorus, only with rhizobium and with rhizobium and phosphorus conditions, respectively. The more divergent the two genotypes are the more will be the probability of improving through selection and hybridization. The current study showed presence of moderate level of resistance in chickpea genotypes against Adzuki bean beetle regardless of soil fertility levels. In the future, it would be good approach to take up those moderately resistant genotypes in comprehensive breeding programs as a parent for hybridization.
Key words: Adzuki bean beetle, chickpea, cluster, diversity, phosphorus, rhizobium.
INTRODUCTION
Ethiopia is one of the secondary centers of diversity for chickpea, while South Asia and the Mediterranean are considered as the primary centers of diversity (van der Maesen, 1987; Upadhyaya, 2003).
Chickpea is among the most important cool season food legumes grown worldwide (FAO, 2008; Gaur et al., 2010). Among the pulse crops, chickpea has consistently maintained a much more significant status, ranking second in area and production after common bean (Phaseolus vulgaris L.) and field pea (Pisum sativum L.) (Gaur et al., 2010).
There are two main chickpea botanical subtypes grown globally, namely Kabuli and Desi, representing two diverse gene pools. The Desi chickpeas are characterized by small angular seed shape, dark seed coat, pink flowers, anthocyanin pigmentation of stem, rough seed surface, and either semi-erect or semi-spreading growth habit. The Desi types account for about 85% of world production. This subtype is the principal type grown in India, Pakistan, Iran, Afghanistan and Ethiopia, whereas the Kabuli types generally have large oval shaped seeds, white flowers, smooth seed surface, lack of anthocyanin pigmentation, semi-spreading growth habit and more suited to the temperate climates (Pundir et al., 1991; Reddy et al., 2007). It is grown mainly in the Middle East, India, Mexico as well as in North America, Australia and Spain. Kabuli types have been recently introduced to Ethiopia but the Desi types are dominating the production (over 80%) in the country in terms of both area coverage and volume of production (Kinfe et al., 2015).
Chickpea serves as a multipurpose crop valuable as an export commodity, as it fixes atmospheric nitrogen, a good "break" crop to different diseases and pests, when grown in rotation with cereals (Kirkegaard et al., 2008; Shiferaw and Teklewold, 2007; Bejiga and Daba, 2006). Moreover, it reduces malnutrition and improves human health, especially for the poor who cannot afford livestock products, and increases livestock productivity as the residue is rich in digestible crude protein content compared to residue from cereals (Wood and Grusak, 2007).
Because of its susceptibility to several abiotic (drought, poor soil fertility, and poor cultural practices) and biotic (diseases, insect pests and weeds) factors, the production of chickpea in Ethiopia has remained constantly low, with a national average yield of 2.01 tons ha-1 (CSA, 2020), which is far below the potential yield of 4-5 tons ha-1 (Fikre, 2016). Among the major problems to increase chickpea production in Ethiopia include the damage inflicted by storage insects. The most important pests of stored grain legume seeds are Callosobruchus chinensis L., Callosobruchus maculates Fabricius, Callosobruchus analis Fabricius, Acanthoscelides obtectus Say, and Bruchus incarnates (Desroches et al., 1995).
Among the bruchid beetles, Adzuki bean beetle is one of the most devastating storage pests throughout the world causing substantial loss during storage (Gowda et al., 1982; Sing et al., 1994; Desroches et al., 1995; Gemechu et al., 2012). Reports indicate that Adzuki bean beetle in chickpea may cause losses of up to 50% in Ethiopia and 28% in Eritrea (Kemal et al., 1994; Haile, 2006).
It is widely agreed that food losses after harvest can be substantial and are important in terms of quantity, quality, and nutritional and economic values (Homan and Yubak, 2011). Commercial grain buyers usually reject or refuse to accept delivery of insect contaminated grain or may pay very low price for it (Hill, 1990; Espinal, 1993; Nchimbi-Mosolla and Miswangu, 2001). Genetic enhancement of insect and disease resistance in chickpea can increase its yield potential by as much as three times (ICRISAT, 1992).
In different countries, effective storage insect pest management through genetic improvement of the host for resistance has thoroughly been reviewed for various crops (Shaheen et al., 2006; Somta et al., 2007, 2008). For example, Ahmed and Yusuf (2007) found some level of insect resistance in groundnut (Arachis hypogea) to both Indian meal moth (Plodia interpunctella) and rust red flour beetle (Tribolium castaneum). Derera et al. (2001) also observed a kind of partial resistance in maize genotypes to the weevil (Sitophilus zeamais). Upadhyaya et al. (2011) reported the importance of utilizing this diversity in chickpea breeding programs in order to develop varieties with high yielder and resistance to stress. Similarly, Gemechu et al. (2012) suggested the importance of exploiting chickpea genetic resources with proven performance for better resistance to Adzuki bean beetle in chickpea breeding programs in Ethiopia. However, chickpea breeding approach in Ethiopia mainly focused on improved seed quality, phenological components and grain yield; while the need for genetic enhancement for resistance to Adzuki bean beetle under different soil fertility levels received less attention.
Genetic diversity resistance to pests may be a reflection of differences in plant health, as mediated by soil fertility management (Phelan et al., 1995). Different authors reported the ability of crop plants to resist insect pests and diseases when grown under optimal (physical, chemical and biological) soil conditions. Again, evidence suggests that resistance is linked directly to the physiology of the plant and thus any factor that affects the physiology of the plant may lead to changes in resistance to insect pests (Slansky, 1990).
Resistant varieties have become a crucial element in the success of many on-going insect pest management programmes, which is an effective, feasible, economical and environmentally safe pest management approach (Pedigo and Higley, 1996; Gemechu et al., 2012). Dobie (1984) suggested that the use of improved grain cultivars with resistance to storage pests could provide a key element in integrated pest management for stored grains. The objective of this study was to understand the diversity of Chickpea genotypes managed under different soil fertility levels for Adzuki bean beetle resistance in Ethiopia.
MATERIALS AND METHODS
Description of Experimental Sites
Both experiments were conducted in Holetta and Debre Zeit Agricultural Research Centers, Ethiopia. Debre Zeit Agricultural Research Center (DZARC) is located in East Shewa Zone of Oromia Regional State in Central Ethiopia, at 08°44’N, 38°58’E and an altitude of 1900 m.a.s.l.. It is characterized by long term mean annual rainfall of 851 mm and mean maximum and minimum temperatures of 28.3 and 8.9°C, respectively. Holetta Agricultural Research Center (HARC) is located in West Shewa Zone of Oromia Regional State in Central Ethiopia, at 09°04'12"′N, 38o29'45"E and an altitude of 2400 m.a.s.l. It is characterized by long term mean annual rainfall of 1064 mm and mean maximum and minimum temperatures of 22.5 and 6.4°C, respectively.
Genetic materials
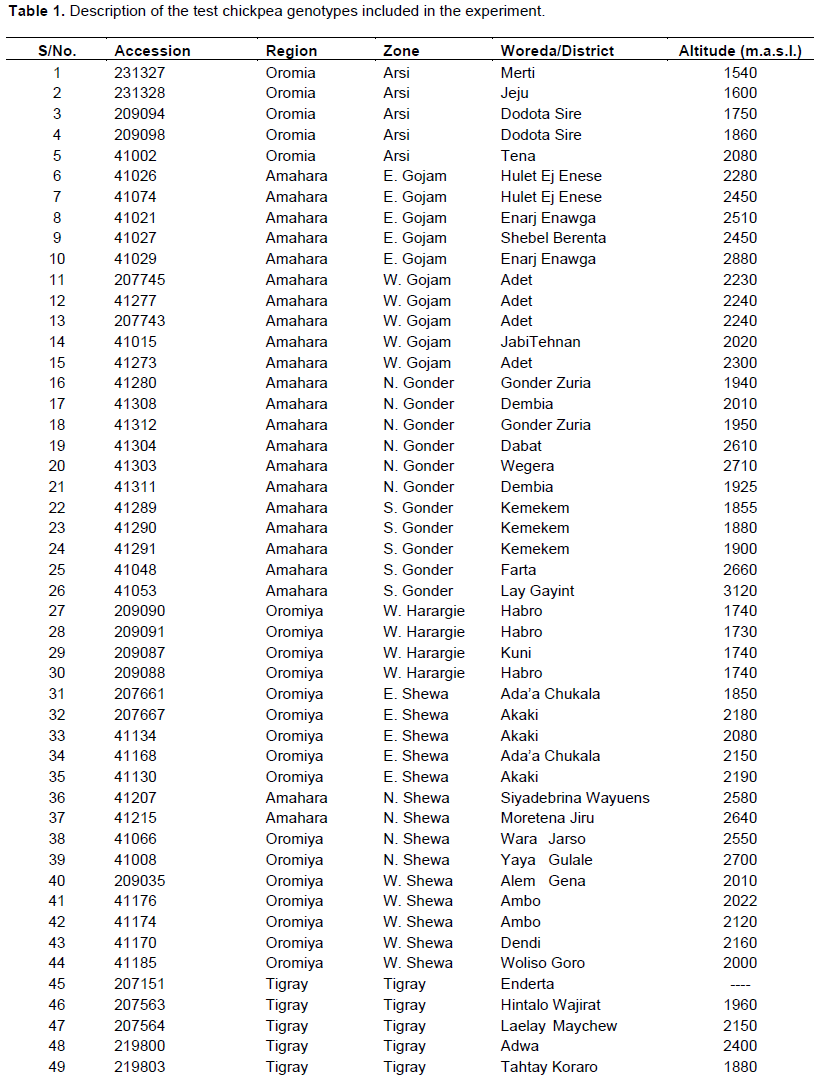
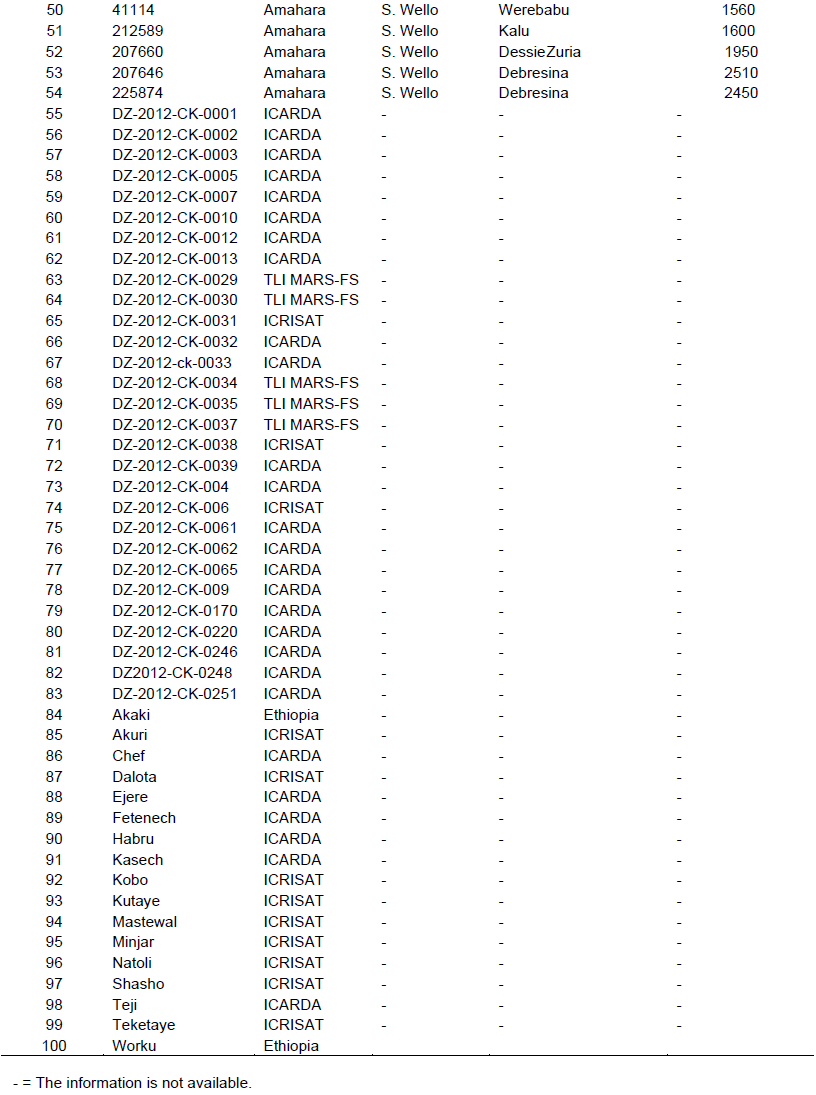
A total of 100 genotypes were used in the study: 54 chickpea germplasm accessions collected from the major chickpea production areas all over the country (Arsi, East Gojam, West Gojam, North Gonder, South Gonder, West Harerge, East Shewa, North Shewa, West Shewa, Tigray, and Wello), 29 pipeline materials and 17 improved varieties. The pipeline materials and released varieties were originally from the Ethiopian Institute of Agricultural Research (EIAR), the International Center for Agricultural Research in the Dry Areas (ICARDA) and the International Crops Research Institute for the Semi-Arid Tropics (ICRISAT). The test genotypes are described in Table 1.
Experimental field layout and management of treatments
All genotypes were grown in 2016 main cropping season in a randomized complete block design with 3 replications in vertisol at Holetta and Debre Zeit under three different soil fertility levels, that is, neither rhizobium nor with phosphorus, only with rhizobium and with phosphorus and rhizobium. Phosphorus was applied at the rate of 20 g per plot (1.2 m2) in the form of triple supper phosphate (TSP) as recommended (Eshete, 1994). An effective isolate of Rhizobium, CP EAL 004, was inoculated at the rate of approximately 1 g of inoculum for 40 seeds using 40% gum Arabic as an adhesive (Somasegaran and Hoben, 1985).
Freshly harvested seeds of each genotypes were cleaned manually from foreign materials and adjusted to 9.0-10% moisture contents by sun-drying and disinfected in a deep freeze at about -20°C for a month prior to the study to eliminate any pre-storage infestation (eggs, larvae and adult bruchids).
Mass-rearing of the insects
Adult beetles were mass-reared using a susceptible chickpea variety Shasho as suggested by Gemechu et al. (2012). The beetles were introduced into 10 kg of seeds from the susceptible variety and kept at ambient temperature and relative humidity for seven days to allow oviposition. Mass-rearing was made at Holetta and Debre Zeit Agricultural Research Centers, Entomology Laboratories. To standardize the age of the progeny, the parent insects were sieved out after seven days. After parent removal, the progenies that emerged were used for re-culturing and, subsequently, 1-2 day old adult insects that emerged were used for the purpose of infestation.
Laboratory experimental design and infestation
The experiment was conducted under ambient room temperature and relative humidity in a randomized complete block Design (RCBD) with 3 replications. Two hundred seeds of each genotypes were allocated per experimental unit (a plastic jar of 250 ml; 6 cm x 7 cm). The chickpea genotypes were assigned to jars at random within each block. Fourteen 1-2 days old unsexed adults of Adzuki bean beetles were collected from the maintained culture and randomly selected and released in each jar. The male to female ratio in this insect being nearly 1:1 (Lemma, 1990), was assumed that each jar received 7 males and 7 female. The ovipositing adults were kept in the jars for 7 days after introduction and then were removed from the jars. The plastic jars containing seeds were inspected on daily basis for the emergence of first progeny. When emergence of the first progeny was completed, the first progeny was removed from the jars for evaluation of the level of attack and loss incurred by the first progeny. With the help of portable thermo-hygrometer, temperature and relative humidity of the room was recorded daily until the end of the experiment to observe the daily fluctuation. The level of temperature and humidity of the laboratory is sometimes observed to be influenced by external environmental situations during the course of experimental periods (Appendix Figure 1).
Data collection
Total number of eggs: Total number of eggs laid on the surface of seeds of each genotype was counted on a daily basis starting from the 4th day to the 14th day of infestation.
Days to adult emergence: The number of days required to adult emergence was recorded on a daily basis starting from the 25th day of infestation until the first adult emerged from seeds.
Number of adults emerged: Total number of adults that emerged from each genotype was counted on a daily basis starting from the 25th day of infestation to until the last adult of the first progeny emerged from seeds.
Susceptibility index (SI): Susceptibility index calculated after Howe (1971) as modified by Dobie (1977) using the formula:
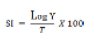
Where SI = susceptibility index, Log Y= log number of first emerged adults, T = mean developmental periods (days), estimated as the time from the middle of oviposition period to 50% emergence of the first progeny. The values of the susceptibility indices were used to rank genotype susceptibility to the bruchids into five categories according to Mensah (1986) as follows:
i. Genotypes with values ranging from 0.0-2.5 were considered resistant genotypes (R).
ii. Genotypes with values ranging from 2.6-5.0 were considered moderately resistant (MR).
iii. Genotypes with value ranging from 5.1-7.5 were considered moderately susceptible (MS).
iv. Genotypes with values ranging from 7.6-10.0 were considered susceptible (S).
v. Genotypes with values greater than 10.0 were considered highly susceptible (HS).
Percentage of seed damage: The percent damage of each genotype was calculated by separating healthy grains (without holes) from the sieved samples and used for percent damage calculations using the formula described by Khattak et al. (1987) as:

Where Nds = number of damaged seed, Tns = total number of seeds.
Adult recovery (%): The actual number of adults that emerged compared with the actual number of eggs laid on the surface of seeds. That is, the ratio of number of adults emerged to number of eggs multiply by one hundred.
Thousand seed weight (g): Cleaned grains sample was taken from each genotype and 1000-grains were weighed in grams after adjusting the moisture content to the standard level (10%).
Proportion of seed coat by weight (%): Seed coat weight as percent of total seed weight of the same genotypes grown under the same conditions was taken from the replicated field trial. That is, ratio of seed coat weight to total weight of the seed multiply by one hundred.
Seed weight loss (g): The seeds were separated into damaged and undamaged categories and weight loss was adjusted to 10% moisture content. The damaged and undamaged seeds were counted and weighed. Percent weight loss was calculated using the formula given by Adams (1976) as follows:
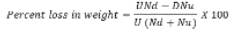
Where U = weight of undamaged grain; D = weight of damaged grain; Nd = number of damaged grain; Nu = number of undamaged grain.
Data analyses
Count data including total number of eggs, number of adults emerged and mean number of holes per seed were log-transformed. Likewise, percent data, adult recovery, proportion of seed coat weight by weight, percent of damage seeds, percent seed weight loss and index of susceptibility were angular-transformed (arcsine proportion) in order to stabilize the variance (Gomez and Gomez, 1984). Data on thousand seed weight and days to adult emergence were not transformed because variance heterogeneity was not observed.
Cluster and distance analyses
Cluster analysis based on the average linkage method was made to examine the assembling pattern of the 100 chickpea genotypes managed under different soil fertility levels. The number of cluster was determined based on Pseudo F and Pseudo t2 statistics. The points where local peaks of the pseudo F-statistic join with small values of the pseudo t2 statistic followed by a larger pseudo t2 for the next cluster combination was used to determine the number of clusters. Dendrograms were constructed based on the Euclidean distance used as a measure of dissimilarity (the distance).
Genetic distances between clusters as standardized Mahalanobis’s D2 statistics (Mahalanobis, 1936) were calculated using the formula:
Dij2 = (xi – xj)’ cov-1(xi – xj) (11)
Where, Dij2 = the distance between cases i and j; xi and xj = vectors of the values of the variables for cases i and j; and cov-1 = the pooled within groups variance-covariance matrix.
Both cluster and distance analyses were done using SAS statistical package (SAS Institute, 2002), whereas the dendrograms were built using the MINITAB 14 statistical package (Minitab, 2010).
RESULTS AND DISCUSSION
Cluster analysis
Cluster analysis was used to group the genotypes into five clusters with and without application of rhizobium and phosphorus, and into six clusters under the application of both rhizobium and phosphorus (Figure 1 and Table 2). This indicated that the higher number of clusters was formed when the crop was grown under the application of both rhizobium and phosphorus, and suggested that the better the soil fertility the better would be the higher genetic expression in terms of genetic diversity. Interestingly, these results are in-agreement with the report of Gemechu et al. (2012) in 155 chickpea genotypes, who reported that cluster analysis grouped the genotypes into five clusters in the absence and six clusters in the presence of phosphorus fertilizer. Similarly, on the same crop, Mussa (2017) reported that the cluster analysis has grouped the 210 chickpea genotypes into 12 distinct clusters under full-water condition and 10 under water-stressed condition. Under the three soil fertility management levels, different cluster contained different number of genotypes.
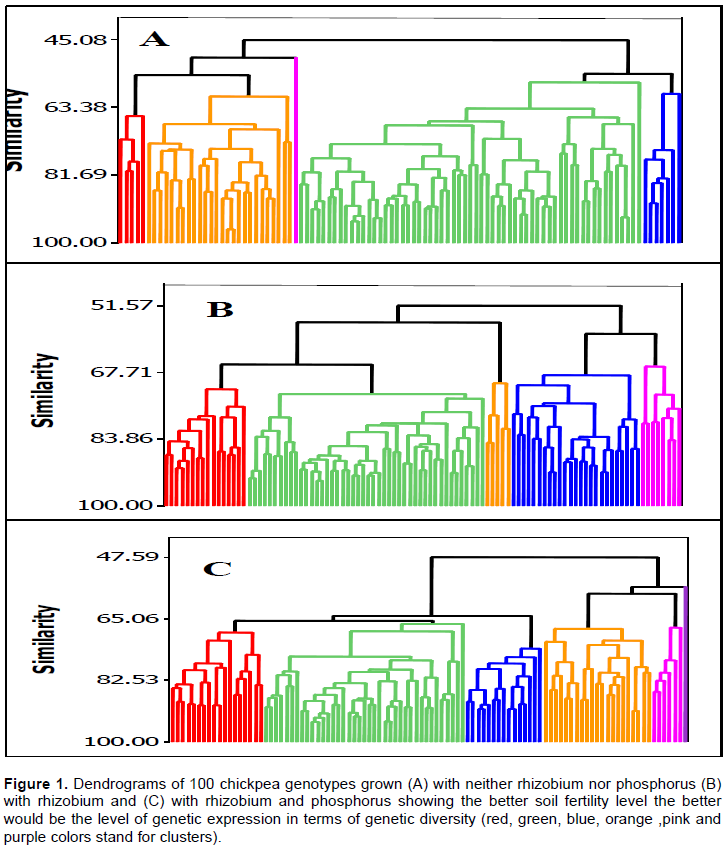

When genotypes were grown without rhizobium and phosphorus, the first cluster (C1) consisted of 5 large-seeded and improved Desi genotypes with relatively thicker seed coat. The second cluster (C2) under the same soil fertility management level consisted of a total of 26 large-seeded and improved Kabuli genotypes with relatively thinner seed coat. Likewise, the third cluster (C3) also consisted of only a single improved Kabuli genotype with large seed size and relatively thinner seed coat. The fourth cluster (C4) under the same soil fertility management was the largest with 61 Desi genotypes with relatively thicker seed coat. The fifth cluster (C5) consisted of seven small-seeded landrace Desi genotypes with relatively thicker seed coat.
When the genotypes were grown under rhizobium inoculation, the first cluster (C1) consisted of 16 genotypes, and the second cluster (C2) 46 small-seeded improved Desi genotypes with relatively thinner seed coat. The third cluster (C3) consisted of 5 Desi genotypes with relatively thicker seed coat, the fourth cluster (C4) 25 genotypes and the fifth cluster (C5) 8 improved large-seeded Kabuli genotypes with relatively thinner seed coat. Except in cluster five where improved Kabuli genotypes were grouped, landraces and improved genotypes were grouped into other clusters.
When the genotypes were grown under rhizobium and phosphorus condition, the first cluster (C1) consisted of 18 landraces and improved Desi genotypes with relatively thicker seed coat. The second cluster (C2) consisted of 39 small-seeded landrace Desi genotypes with relatively thicker seed coat. The third cluster (C3) consisted of 15 small- and medium-seeded Desi genotypes with relatively thinner seed coat. The fourth cluster (C4) consisted of 21 (20 landrace and one improved) Desi genotypes including landraces and improved ones with relatively thicker seed coat. The fifth cluster (C5) consisted of six Kabuli and Desi improved genotypes while the sixth cluster (C6) consisted of only a single improved Kabuli large-seeded genotype with relatively thinner seed coat.
Distance analysis
Genetic distances (D2) among the clusters of the 100 chickpea genotypes grown under different soil fertility levels for response characters to infestation by Adzuki bean beetle are presented in Table 3. The inter-cluster distances (D2) between the possible pairs of clusters were found to be mostly significant (P ≤ 0.05 or P ≤ 0.01) regardless of different soil fertility levels. Inter-cluster D2 average values of 11.50 (between clusters C4 and C5) to 62.94 (between clusters C2 and C5), 14.85 (between clusters C1 and C2) to 72.12 (between clusters C3 and C5) and 8.61 (between clusters C2 and C3) to 166.25 (between clusters C2 and C6) was found when genotypes were grown under neither rhizobium nor phosphorus, only with rhizobium and with rhizobium and phosphorus conditions respectively.

When the three soil fertility management levels were compared, the maximum genetic divergence (D2 = 166.25) was observed when the genotypes were grown under the best soil fertility management level, that is, with both rhizobium inoculation and phosphorus application.
It is generally believed that more conducive environments may be expected to result in better expression of the genetic potential of the genotypes for the traits under consideration (Rosielle and Hamblin, 1981; Simmonds, 1991; Singh, 2002) despite the controversy that there may be no interrelationship between the type of the environment and the magnitude of genetic variation (Ceccarelli and Grando, 1996). Hybridization of parental lines selected from clusters C2 with those selected from cluster C6 (D2 = 166.25) in the genotypes grown with both rhizobium inoculation and phosphorus application is expected to result in good progenies. It is generally believed that crosses between parental genotypes selected from the most divergent clusters would result in maximum genetic recombination and segregation in the resultant progenies.
However, parental selection should take into consideration not only genetic divergence for multiple traits but also specific merits of the particular genotype for the particular trait of interest like disease resistance and desirable agronomic and quality performances (Singh, 1990; Chahal and Gosal, 2002). There were no significant (P > 0.05) genetic divergence between some of the clusters regardless of the soil fertility management levels, indicating close relation among the member genotypes in these clusters for the traits considered in this study. It should be noted that crosses of parental pairs selected from the respective clusters with minimal divergence would not produce good level of genetic recombination and segregation in progenies for effective selection.
Average performances of genotypes in different clusters
Cluster mean performances showed existence of considerable variation among the different clusters for individual traits to infestation by Adzuki bean beetle under the three soil fertility management levels (Table 4). The first cluster (C1) under neither rhizobium nor phosphorus application, the third cluster (C3) under rhizobium inoculation and the second cluster (C2) under the application of both rhizobium and phosphorus were characterized by the least number of eggs laid, adults emerged, percentage of adult recovery, number of holes per seed, percentage of damage seed, and seed weight loss. This indicated that these clusters consisted of small to medium seeded genotypes with thick seed coats and moderate resistance to Adzuki bean bettele. When compared to the other clusters, those clusters were totally dominated by Desi type genotypes which showed better performance for resistance may be due to seed related traits that hindered reproduction and establishment of the insect.
On the contrary, under neither rhizobium nor phosphorus condition the third cluster (C3), under rhizobium and with rhizobium and phosphorus the fifth cluster (C5) were characterized by the highest number of eggs laid and adults emerged, highest percentage of adult recovery, highest mean number of holes per seed, highest percentage of damaged seed by number, highest seed weight loss, highly susceptible to bruchid attack, and largely seeded with thin seed coat. When compared to the other clusters those clusters were dominated by Kabuli type genotypes which showed least performance in most insect and seed related traits.
The remaining clusters were found to have exhibited intermediate mean value for both seed and insect related traits. Thus, the current result was in line with earlier works which reported that the susceptible genotypes with soft smooth seed coat, white seed color and bigger grain size resulted to higher insect damage (Gemechu et al., 2012; Sarwar, 2012). Similarly, Pacheco et al. (1994) reported that the level of resistance against insect damage related with physical characteristics of the seed coat such as dark color, roughness, and thickness. Again, Winn (1988) suggested that seed size is known as one of the factors for determining the level of resistance to bruchid species while predation is considered to result in selection for smaller seeds in wild legumes. Likewise, Nwanze and Horber (1975) demonstrated that cowpea beetle (C. maculates) prefers smooth seeded to rough seeded cowpeas.
Geographic pattern of genetic diversity
The large majority of the genotypes, including released varieties, tended to concentrate on a limited number of clusters but, generally, the pattern of distribution of the genotypes originating from different eco-geographical regions of the country across different zones (Arsi, East Gojam, West Gojam, North Gonder, South Gonder, West Harargie, East Shewa, North Shewa West Shewa, Tigray and South Wello) showed that genotypes from the different origins may be grouped together into the same clusters and genotypes from the same origins were grouped into different clusters (Table 5). The concentration of the large majority of the genotypes into a few clusters may indicate existence of limited genetic diversity for response characters to infestation by Adzuki bean beetle. Similar results were also reported from an earlier study of chickpea genotypes mostly collected from Ethiopia and a few from exotic sources (Gemechu et al., 2012). The tendency of most of the released varieties to be concentrated into a few clusters may indicate that breeders have emphasized similar characters during selection and might develop genetic materials with similar genetic background as in most modern varieties (Heisey and Brennan, 1991).
CONCLUSION
Application of both rhizobium and phosphorus received higher number of clusters than crops grown without application of rhizobium and phosphorus. This showed that the better the soil fertility the better would be the higher genetic expression in terms of genetic diversity. Singh (2002) also suggested that the more conducive the test environment, the better would be the level of genetic expression of the multiple traits and the level of genetic diversity, not only, in terms of the magnitude of the D2 values, but also, in terms of the number of clusters formed.
Under the three soil fertility levels, genetic diversity for individual traits to infestation by Adzuki bean beetle was observed. For instance, Desi type genotypes under each cluster showed relatively least level of infestation against the beetle may be due to seed related traits like least mean number of holes per seed, low percentage of seed damage, least seed weight loss, small seed size with thin seed coat. that hindered reproduction and establishment of the insect. Similarly, Pacheco et al. (1994) reported that the level of resistance against insect damage related with physical characteristics of the seed coat such as dark color, roughness, and thickness.
Generally, to broaden the source of genetic variability of chickpea in Ethiopia, it would be good approach take up moderately resistant genotypes in a comprehensive breeding program as a parent for hybridization.
CONCLUSION
The authors have not declared any conflict of interests.
ACKNOWLEDGMENTS
The authors are grateful to Ethiopian Institute of Agricultural Research for financial support and staff members of Holetta and Debre Zeit Agricultural Research Centers, and Jimma University for assistancing the research work.
REFERENCES
|
Adams JM (1976). A guide to the objective and reliable estimation of food losses in a small scale farmer storage. Tropical Stored Product 32:5-12. |
|
|
Ahmed BI, Yusuf AU (2007). Host-plant resistance as viable non-chemical and environmentally friendly strategy of controlling stored products pests. Emirates Journal of Food and Agriculture pp. 1-12. |
|
|
Bejiga G, Daba K (2006). Breeding chickpea for wide adaptation. Ali K, Keneni G, Ahmed S, Malhotra R, Beniwal S, Makkouk K, Halila. |
|
|
Ceccarelli S, Grando S (1996). Importance of specific adaptation in breeding for marginal conditions. In: Barley Research Review Workshop Proceeding, 16-19 October 1993, Addis Abeba, Ethiopia pp. 101-111. |
|
|
Central Statistical Authority, CSA (2020). Agriculture Sample Survey, Area and Production of Major Crops. Statistical Bulletin 13p. |
|
|
Chahal GS, Gosal SS (2002). Principles and procedures of plant breeding.Biotechnological and conventional approaches.Alpha Science, Oxford, United Kingdom 412 p. |
|
|
Derera J, Pixley KV, Denash G (2001). Resistance of maize to the maize weevil l: I. Antibiosis. |
|
|
Desroches P, Shazly E, Mandon N, Duc G, Huignard J (1995). Development of Callosobruchus chinensis (L.) and Callosobruchus maculatus (F.) (Coleoptera: Bruchidae) in seeds of Vicia faba L. differing in their tannin, vicine and convicine contents. Journal of Stored Products Research 31(1):83-89. |
|
|
Dobie P (1977). The contribution of the Tropical Stored Products Centre to the study of insect resistance in stored maize. Journal of Stored Products Research 14:87-93. |
|
|
Dobie P (1984). Biological methods for integrated control of insects and mites in tropical stored products. In. McFarlane JA (Eds), The use of resistant varieties. Tropical Stored Products Information, Natural Resources Institute, Chatham, UK. P 28. |
|
|
Eshete M (1994). Chickpea and Lentil Agronomy Research. In: The First Review Conference National Cool-Season Food Legumes Conference Proceeding, 16-20 Dec 1993, Addis Abeba ,Ethiopia pp. 230-250. |
|
|
Espinal JR (1993). Economic losses associated with Zabrotes subfasciatus and Acanthoscelides obtectus infestations of stored dry red beans (Phaseolus vulgaris L.) in southeastern Honduras. Doctoral dissertation, Kansas State University. |
|
|
FAO ( 2008). Food and Agriculture Organization of the United Nations Statistical, Rome, Italy. |
|
|
Fikre A (2016). Progresses of chickpea research and development in Ethiopia. In. Korbu L, Damte T, Fikre A (Eds.). Harnessing Chickpea Value Chain for Nutrition Security and Commercialization of Smallholder Agriculture in Africa, Debre Zeit, Ethiopia. pp. 25-39. |
|
|
Gaur PM, Tripathi S, Gowda CLL, Ranga Rao GV, Sharma HC, Pande S, Sharma M (2010). Chickpea Seed Production Manual. Andhra Pradesh, India. 28p. |
|
|
Gemechu K , Bekele E, Getu E, Imtiaz M, Dagne K, Assefa F (2012). Genetic Gain for Adzuki Bean Beetle (Callosobruchus chinensis L.) Resistance in Ethiopian Chickpea (Cicer arietinum L.) Genotypes. East African Journal of Sciences 6(1):43-54. |
|
|
Gomez KA, Gomez A (1984). Statistical Procedures for Agricultural Research. John Wiley Sons, New York. |
|
|
Gowda CLL, Kaul AK (1982). Pulses in Bangladesh, Bangladesh Agricultural Research Institute, FAO, UN. 172p. |
|
|
Haile A (2006). On-farm storage studies on sorghum and chickpea in Eritrea. African Journal of Biotechnology, 5(17): 57-79 |
|
|
Heisey PW, Brennan JP (1991). An analytical model of farmers' demand for replacement seed. American Journal of Agricultural Economics 73(4):1044-1052. |
|
|
Hill DS (1990). Pests of stored product and their control, 1st edition, Cambridge University press, London 780 p. |
|
|
Homan R, Yubak GC (2011). Eco-friendly management of pulse beetle. Journal of Agriculture and Environment 12:1-10. |
|
|
Howe RW (1971). A parameter for expressing the suitability of an environment for insect development. Journal of Stored Products Research 7(1):63-65. |
|
|
ICRISAT (1992). The Medium Term Plan.International Crops Research Institute for the Semi-Arid Tropics. Patancheru, Andhra Pradesh, India. 229p. |
|
|
Kemal A, Tibebu H, Asfaw T, Geletu B, Saxena MC, Solh MB (1994). Cool-Season Food Legumes of Ethiopia. |
|
|
Khattak SUK, Hamed M, Khatoon R, Mohammad T (1987). Relative susceptibility of different mungbean varieties to Callosobruchus maculates. Journal of Stored Products Research 23(3):139-142. |
|
|
Kinfe E, Singh P, Fekadu T (2015). Physicochemical and Functional Characteristics of Desi and Kabuli Chickpea (Cicer arietinum L.) Cultivars Grown in Bodity, Ethiopia and Sensory Evaluation of Boiled and Roasted Products Prepared Using Chickpea Varieties. International Journal of Current Research in Biosciences and Plant Biology 2(4):21-29. |
|
|
Kirkegaard J, Christen O, Krupinsky J, Layzell D (2008). Break crop benefits in temperate wheat production. Field Crops Research 107(3):85-195. |
|
|
Lemma T (1990). The biology and control of the Adzuki bean beetle (Callosobruchus chinensis L.) on chickpea (Cicer arietinum L.). MSc Thesis, Alemaya University, Ethiopia. |
|
|
Mahalanobis PC (1936). On generalized distance in statistics. National Institute of Science Proceeding, 16 April 1936, India. pp. 49-55. |
|
|
Mensah G (1986). Infestation potential of Callosobruchus maculatos on cowpea cultivars stored under subtropical conditions. International Journal of Tropical Insect Science 7(06):781-784. |
|
|
Minitab Inc (2010). Statistical Software Language Pack English Version 16.1.1.1. Minitab, Inc., PA, USA. |
|
|
Mussa J (2017). Genetic Improvement Of Adapted Ethiopian Chickpea (Callosobruchus maculatus L.) Cultivar For Drought Tolerance Through Conventional And Markerassisted Backcross Breeding Methods. Doctoral Dissertation, Addis Ababa University, Ethiopia. |
|
|
Nchimbi-Mosolla S, Misangu RN (2001). Seasonal distribution of common bean bruchid species in selected areas in Tanzania.In Proceedings of the Bean Seed Workshop, Arusha, Tanzania. pp. 12-14. |
|
|
Nwanze KF, Horber E (1975). Laboratory Techniques for Screening Cowpeas for Resistance to Callosobruchus maculatus. Environmental Entomology 4(3):415-419. |
|
|
Pacheco IA, Bolonhezi S, Sartori MR, Turatti, JM, Paula DC, de Lourencao AL (1994). Resistance to bruchids, fatty acid composition and grain texture in genotypes of chickpea. Bragantia 53:61-74. |
|
|
Pedigo LP, Higley LG (1996). Introduction to Pest Management and Economic Thresholds for Integrated Pest Management.University of Nebraska Press, Lincoln, USA pp. 3-8. |
|
|
Phelan PL, Mason JF, Stinner BR (1995). Soil-fertility management and host preference by European corn borer, Ostrinianu bilalis, on Zea mays. A comparison of organic and conventional chemical farming. Agriculture, Ecosystems And Environment 56(1):1-8. |
|
|
Pundir RPS, Reddy KN, Mengesha MH (1991). Genetics of some physio-morphic and yield traits of chickpea (Cicer arietinum L.). Legume Research 14(4):157-161. |
|
|
Reddy AA, Mathur VC, Yadav M, Yadav SS (2007). Commercial cultivation and profitability. Chickpea Breeding and Management. pp.292-321. |
|
|
Rosielle A, Hamblin J (1981). Theoretical aspects of selection for yield in stress and non-stress environments. Crop Science 21:943-946. |
|
|
Sarwar M (2012). Assessment of resistance to the attack of bean beetle Callosobruchus maculates in chickpea genotypes on the basis of various parameters during storage. Journal of Science and Technology 34(3):241-345. |
|
|
SAS Institute (2002). SAS/STAT system guide for personal computers, version 9.0 edition. SAS Institute Inc., Cary, North Carolina, USA |
|
|
Shaheen FA, Khaliq A, Aslam M (2006). Resistance of chickpea (Cicer arietinum L.) cultivars against pulse beetle. Pakistan Journal of Botany 38(4):1237. |
|
|
Shiferaw B, Teklewold H (2007). Structure and functioning of chickpea markets in Ethiopia: Evidence based on analyses of value chains linking smallholders and markets. Nairobi, Kenya 63 p. |
|
|
Simmonds NW (1991). Selection for local adaptation in a plant breeding programme. Theoretical and Applied Genetics, 82(3):363-367. |
|
|
Singh DN, Nandi A, Tripathy P (1994). Genetic variability and character association in French bean (Phaseolus vulgaris). Indian Journal of Agricultural Sciences 2:114-116. |
|
|
Singh KB (1990). Prospects of developing new genetic material and breeding methodologies for chickpea improvement. Mediterraneennes 9:43-50. |
|
|
Singh BD (2002). Plant breeding, principles and methods. Kalyani Publishers, New Delhi Ludhiana. |
|
|
Slansky Jr F (1990). Insect nutritional ecology as a basis for studying host plant resistance. Florida Entomologist 3:359-378. |
|
|
Somasegaran P, Hoben H.J (1985). Methods in Legume-Rhizobium Technology.Department of Agronomy and Soil Science, University of Hawaii, USA 510 p. |
|
|
Somta P, Ammaranan C, Ooi PAC, Srinives P (2007). Inheritance of seed resistance to bruchids in cultivated mungbean (Vignaradiata, L.). Euphytica 155(1-2):47-55. |
|
|
Somta P, Kaga A, Tomooka N, Isemura T, Vaughan DA, Srinives P (2008). Mapping of quantitative trait loci for a new source of resistance to bruchids in the wild species Vigna nepalensis Tateishi & Maxted (Vigna subgenus Ceratotropis). Theoretical and Applied Genetics 117(4):621-628. |
|
|
Upadhyaya HD (2003). Geographical patterns of variation for morphological and agronomic characteristics in the chickpea germplasm collection. Euphytica 132(3):343-352. |
|
|
Upadhyaya HD, Dronavalli N, Gowda CLL, Singh S ((2011). Identification and evaluation of chickpea germplasm for tolerance to heat stress. Crop Science 51(5):2079-2094. |
|
|
Van der Maesen L (1987). Origin, history and taxonomy of chickpea. In. Saxena MC, Singh KB (Eds.). CAB International Cambrian News Ltd, Aberystwyth UK pp. 11-34. |
|
|
Winn AA (1988). Ecological and evolutionary consequences of seed size in Prunella vulgaris. Ecology 69(5):1537-1544. |
|
|
Wood JA, Grusak MA (2007). Nutritional value of chickpea. Chickpea Breeding and Management pp. 101-142. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


