Full Length Research Paper

ABSTRACT
The conversion of tropical field corn genotypes into sweet corn could broaden the genetic base and improve yield and adaptation of sweet corn varieties. In this study, the performance of shrunken-2 (sh2) super-sweet corn populations derived from crosses between a sh2 population and tropical field corn genotypes were evaluated in Ibadan. Experiments were conducted using randomised complete block design with three replicates. Data were collected on agronomic and fresh ear yield traits, and then subjected to analysis of variance. Significant genotypic differences were observed among the populations with most of the derived populations significantly superior to the donor population for most of the traits. Yield of marketable cobs ranged from 5.80 to 7.63 t/ha (mean = 6.84 t/ha). Six derived populations had significantly higher yield of marketable cobs than the donor population. On the average, 83.1% of the number of cobs harvested was marketable. Husk cover scores ranged from 2.8 to 6.8, with all the derived populations having significantly lower husk cover scores than the donor population. The results indicated that the conversion of the field corn genotypes into super-sweet corn was effective in the development of new super-sweet corn populations. The observed genetic differences could be exploited in further breeding programmes.
Key words: Fresh ear yield, husk cover, shrunken-2 super-sweet corn, yield of marketable cobs
INTRODUCTION
Maize (Zea mays L.) is an important and highly diversified crop in the world, and a cereal food staple cultivated in every agro-ecological zone in West and Central Africa. It is made up of different types, classified by their kernel endosperm characteristics. The most common types are; flint, dent, flour, pop, pod, waxy and sweet. In West and Central Africa, the flint and dent types are the most widely cultivated (Kim et al., 1987; Kim and Ajala, 1996). These field maize types, in addition to the dry grains being consumed in different processed forms, are harvested and consumed fresh as green maize (after roasting, steaming or boiling on the cob). This contrasts with developed countries where human consumption of fresh maize is of sweet and super-sweet corn (Osayintola et al., 1992; Lee et al., 1999; Zan and Brewbaker, 1999). The widely grown field maize varieties in West and Central Africa were however not specifically developed for direct fresh human consumption (Ogunbodede, 1999).
Sweet corn (Zea mays L. var saccharata), is a type of maize with kernels that are sweet as a result of high sugar content which when consumed in the fresh immature stage, has high levels of total sugars than field maize, rich in fibre, minerals, certain vitamins (Tracy, 1997; Lertrat and Pulam, 2007), and significant antioxidant properties (Dewanto et al., 2002). Sweet corn has its origin from a mutation that influences carbohydrate biosynthesis in the endosperm. In the genome of sweet corn, at least one of the eight mutant genes preventing the conversion of sugars to starch is present. These genes, which include shrunken-2 (sh2) on chromosome 3; brittle (bt), and amylose extender (ae) on chromosome 5; sugary enhancer (se), sugary (su), and brittle-2 (bt2) on chromosome 4; dull (du) on chromosome 10; and waxy (wx) on chromosome 9 (Tracy et al. 2006, Qi et al., 2009) are monogenic and recessive (Santos et al., 2014). Of all the mutant types, sh2 (Yousef and Juvik, 2002) and bt2 (Brewbaker, 1977) have the greatest commercial value.
The sh2 types of sweet corn contain about 29.9% sucrose, which is about ten times that of field maize. These types of sweet corn, generally referred to as super-sweet or extra-sweet corn, have extended shelf life due to the slow conversion of sugars to starch after harvest (Tracy, 1997), but with highly reduced levels of total carbohydrate. The sh2 gene reduces endosperm content of starch and water soluble polysaccharides which leads to a reduction in the energy content of maize kernels and the markedly collapsed physical appearance of the kernels when dry. Due to these features, sh2 varieties generally have a significantly reduced germination, seedling emergence, seedling vigour, plant development and growth and poor stand establishment.
In Nigeria, maize is grown throughout the country from the high rainfall forest of the southeast to the low rainfall Sudan savanna of the north; and with supplemental irrigation, maize can be grown throughout the year. Nigeria produces about 40% of the maize production in West and Central Africa (FAO, 2016). Great potential therefore exists for the production of super-sweet corn in Nigeria. Sweet corn production is usually targeted at three distinct and largely independent markets, namely; fresh, canning and freezing, with the fresh market component accounting for more than 70% of the total (Lizaso et al., 2007; USDA, 2017). Imports of canned sweet corn to Nigeria have increased in recent years largely due to widening food preferences. Therefore, the development and production of super-sweet corn in large quantity and quality would impact positively on the social and economic life of Nigeria. However, sweet corn cultivars are virtually nonexistent in Nigeria, and coupled with poor adaptation to tropical environments, its cultivation and utility is limited.
To bridge the gap created by these challenges, a broad-base temperate super-sweet sh2 corn population was introduced into the country and adapted to the prevailing tropical environmental conditions by four cycles of mass selection (Adetimirin, 2008). This was meant to serve both as an open pollinated variety, as well as basis for inbred line development and hybrid production. The performance of this sh2 super-sweet population in the growing conditions of Nigeria could be improved. One way of achieving this is by converting tropical field maize genotypes into sweet corn through backcrossing and selection for the sweet corn trait. This strategy will lead to improvement in the yield of the super-sweet corn varieties, broaden the narrow genetic base characteristic of sh2 (Tracy, 2001; Teixeira et al., 2013), and facilitate the development of sh2 inbred lines and hybrids. According to Entringer et al. (2017), the backcross method of breeding is efficient for obtaining super-sweet corn populations with good agronomic performance. Previous studies Cartea et al. (1996), Malvar et al. (1997, 2001), Tracy (2001), Butrón et al. (2008) and Entringer et al. (2017) reported the use of field corn to improve the agronomic performance of sweet corn. These studies have also shown that field maize genotypes could differ in their ability to improve the agronomic performance and quality of sweet corn. Therefore, tropical normal endosperm field maize genotypes have the potential to improve the adaptation and productivity of super-sweet corn. The objective of this study was to evaluate the agronomic performance of some sh2 super-sweet corn populations derived from crosses between a tropicalised sh2 population and tropical normal endosperm field corn genotypes.
MATERIALS AND METHODS
Location of experimental site
The study was conducted at the experimental field of the Department of Agronomy, Faculty of Agriculture, along Parry road, University of Ibadan (7°26' N, 3°54' E), Ibadan, Nigeria.
Generation of new homozygous sh2 super-sweet corn populations
A broad-based temperate super-sweet sh2 maize population, which was introduced and adapted to tropical environmental conditions of Ibadan, Nigeria after four cycles of mass selection (Adetimirin, 2008), was crossed as male (using bulk pollen) to six tropical field maize inbred lines (1368, 4001, 4008, 9613, KU1409 and KU1414) and five commercial/experimental hybrids [1368/9071 (Oba super 1), 4001/KU1414 (Oba super 2), KU1409/9613, KU1409/4008 and 4001/4008], all developed at the International Institute of Tropical Agriculture (IITA), Ibadan, Nigeria
The resulting F1 populations from each of these initial crosses were sib-mated to generate F2 populations. The F2 seeds were thereafter selected for kernels homozygous for the sh2 phenotype. The selected homozygous sh2 derived from each of the F2 populations were backcrossed to the recurrent (field maize) parents. The BC1 plants from each population were sib-mated to generate BC1S1 which were thereafter selected for kernels homozygous for the sh2 phenotype. The selected homozygous sh2 derived from each of the BC1S1 populations were then bulked to form new homozygous sh2 populations as shown in Table 1.
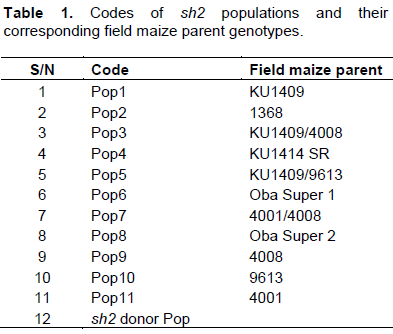
Evaluation of sh2 super-sweet corn populations
The eleven new homozygous derived sh2 populations alongside the original donor sh2 population were evaluated in replicated field trials under rain-fed conditions during the 2016 and 2017 cropping seasons. The soil at the experimental site is sandy-loam with 15.20 g kg-1 organic carbon, 0.98 g kg-1 total nitrogen, 14.79 mg kg-1 available P (Bray-1), 0.26 cmol kg-1K and a pH(H2O) of 6.1. The twelve sh2 populations were arranged in a randomized complete block design with three replicates. Each plot consisted of four 5.0 m long rows, with plants spaced 0.50 m and rows 0.75 m apart. Planting was done on the flat. Plots were over-sown to ensure good plant stand (since germination failure is a common feature in super-sweet corn with the sh2 gene) and later thinned to two plants at two weeks after planting (WAP) to give a plant population of approximately 53,333 plants per hectare.
Data collection
Data were collected on days to anthesis (DA) as number of days from planting to when 50% of the plants in a plot shed pollen, and days to silking (DS) as number of days from planting to when 50% of the plants in a plot have emerged silks. Anthesis-silking interval (ASI) was calculated as the difference in days between DS and DA. Plant height (PH) and ear height (EH) were measured in meters 2 weeks after silking on all plants in the two middle rows of a plot, as the average distance from the soil level to the collar of the uppermost leaf and collar of the leaf bearing the uppermost ear, respectively. Husk cover (HC) was scored on plot basis on a scale of 1 to 9 (1 = husk tightly covers ear tip and extends beyond it; 9 = poor husk cover with ear tip clearly exposed). Harvesting for yield data was carried out between stages R4 and R5 when the ears were still green and fresh, 21 days after silking using plants in the two middle rows of a plot. Yield data included: (i) number of cobs (NC) recorded as total number of ears (fresh ears with husk removed) harvested per plot expressed per hectare; (ii) yield of cobs (YC) recorded as total weight of cobs (fresh ears with husk removed) harvested per plot expressed in tonnes per hectare; (iii) number of marketable cobs (NMC) recorded as number of cobs with approximately 250 filled edible kernels per plot, expressed per hectare; (iv) yield of marketable cobs (YMC) recorded as total weight of marketable cobs per plot expressed in tonnes per hectare; (v) number of kernel rows (NKR) recorded as the average number of kernel rows of 10 top cobs; (vi) cob length (CL) measured in cm as the average length of 10 top cobs; (vii) cob diameter (CD) was measured using an electronic 6 in digital calliper (Pittsburgh®, Item #47257) as the average diameter of 10 top cobs taken at the middle portion of the cob.
Data analyses
All data were subjected to analysis of variance for a randomized complete block using the PROC. GLM procedure in SAS (SAS Institute, 2003) assuming a mixed model with genotype and the interaction between genotype and environment were considered random. The years were considered as separate environments. Significant means were separated using DMRT (p=0.05).
RESULTS
Evaluation of sh2 super-sweet corn populations
The eleven new homozygous derived sh2 populations alongside the original donor sh2 population were evaluated in replicated field trials under rain-fed conditions during the 2016 and 2017 cropping seasons. The soil at the experimental site is sandy-loam with 15.20 g kg-1 organic carbon, 0.98 g kg-1 total nitrogen, 14.79 mg kg-1 available P (Bray-1), 0.26 cmol kg-1K and a pH(H2O) of 6.1. The twelve sh2 populations were arranged in a randomized complete block design with three replicates. Each plot consisted of four 5.0 m long rows, with plants spaced 0.50 m and rows 0.75 m apart. Planting was done on the flat. Plots were over-sown to ensure good plant stand (since germination failure is a common feature in super-sweet corn with the sh2 gene) and later thinned to two plants at two weeks after planting (WAP) to give a plant population of approximately 53,333 plants per hectare.
Data collection
Data were collected on days to anthesis (DA) as number of days from planting to when 50% of the plants in a plot shed pollen, and days to silking (DS) as number of days from planting to when 50% of the plants in a plot have emerged silks. Anthesis-silking interval (ASI) was calculated as the difference in days between DS and DA. Plant height (PH) and ear height (EH) were measured in meters 2 weeks after silking on all plants in the two middle rows of a plot, as the average distance from the soil level to the collar of the uppermost leaf and collar of the leaf bearing the uppermost ear, respectively. Husk cover (HC) was scored on plot basis on a scale of 1 to 9 (1 = husk tightly covers ear tip and extends beyond it; 9 = poor husk cover with ear tip clearly exposed). Harvesting for yield data was carried out between stages R4 and R5 when the ears were still green and fresh, 21 days after silking using plants in the two middle rows of a plot. Yield data included: (i) number of cobs (NC) recorded as total number of ears (fresh ears with husk removed) harvested per plot expressed per hectare; (ii) yield of cobs (YC) recorded as total weight of cobs (fresh ears with husk removed) harvested per plot expressed in tonnes per hectare; (iii) number of marketable cobs (NMC) recorded as number of cobs with approximately 250 filled edible kernels per plot, expressed per hectare; (iv) yield of marketable cobs (YMC) recorded as total weight of marketable cobs per plot expressed in tonnes per hectare; (v) number of kernel rows (NKR) recorded as the average number of kernel rows of 10 top cobs; (vi) cob length (CL) measured in cm as the average length of 10 top cobs; (vii) cob diameter (CD) was measured using an electronic 6 in digital calliper (Pittsburgh®, Item #47257) as the average diameter of 10 top cobs taken at the middle portion of the cob.
Data analyses
All data were subjected to analysis of variance for a randomized complete block using the PROC. GLM procedure in SAS (SAS Institute, 2003) assuming a mixed model with genotype and the interaction between genotype and environment were considered random. The years were considered as separate environments. Significant means were separated using DMRT (p=0.05).
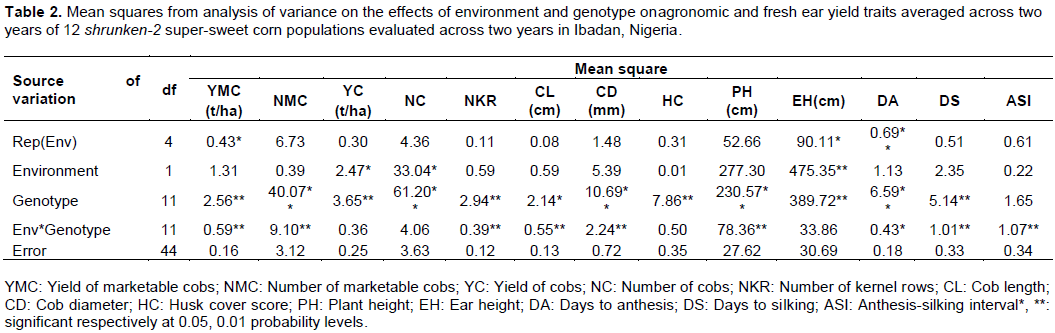
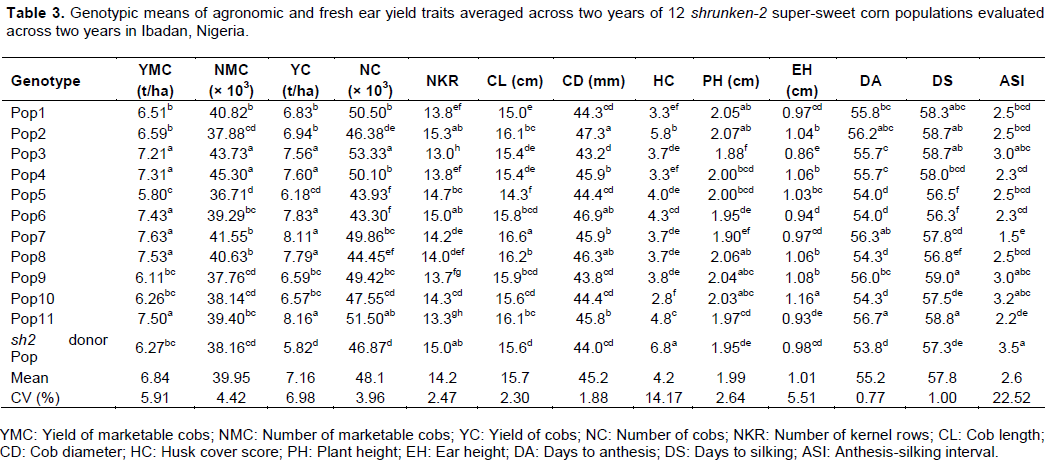
The YC ranged from 5.82 to 8.16 t/ha. The range in NC and NMC were 43,290 to 53,330 and 36,710 to 45,300 cobs/ha, respectively. On the average, 83.1% of the total NC harvested was marketable. The NKR, CL and CD averaged 14.2, 15.7 cm and 45.2 mm, respectively.
One important quality attribute in fresh market sweet corn production is HC. In the present study, HC scores ranged from 2.8 to 6.8 with a mean of 4.2. All the derived sh2 super-sweet corn populations had significantly lower HC scores than the donor sh2 population, eight of which had scores lower than the mean. The PH and EH of the populations ranged from 1.9 to 2.1 m (mean = 2.0 m) and 0.9 to 1.2 m (mean = 1.0 m), respectively. On the average, ears were placed mid-way the height of the plants.
All the population including the sh2 donor population exhibited intermediate maturity with DA and DS ranging from 53.8 to 56.7 days and 56.3 to 59.0 days, respectively. However, nine of the derived populations shed pollen significantly later than the donor sh2 population. On the average, the derived sh2 super-sweet corn populations flowered significantly later than the donor sh2 population. The ASI, which is an indication of the extent of synchrony in flowering ranged from 1.5 to 3.5 days with a mean of 2.6 days. Eight of the derived sh2 populations had significantly lower ASI that the donor sh2 composite population.
In this study, orthogonal comparison revealed significant differences between the derived sh2 populations and the donor sh2 composite population for all the traits, with the derived sh2 populations being superior to the donor sh2 composite (Table 3).
DISCUSSION
Sweet corn cultivars are virtually non-existent in Nigeria, have narrow genetic base and poor adaptation to tropical environments. One strategy that could be adopted to broaden its genetic base, improve its adaptation and enhance its agronomic performance under tropical conditions is to cross with tropical field maize. In this study, some sh2 super-sweet corn populations derived from crosses between sh2 super-sweet corn population and tropical field maize genotypes were evaluated for their agronomic performance.
The significant effects of genotype and the higher contribution of the main effects of genotype to the total sum of squares observed in this study was a manifestation of the fact that the populations varied greatly in their performance. These genetic differences could be exploited in further breeding programmes. These significant genetic differences also indicated that the conversion of the field corn genotypes into super-sweet corn was effective in the development of new sweet corn populations different from the sh2 donor parent. This was furthermore evidenced in the superiority of most of the derived populations over the sh2 donor parent population for most of the traits studied. In this study, yield of marketable cobs averaged 6.84 t/ha. This was higher than the average yield of marketable fresh cobs of 5.03 t/ha reported by Kim et al. (2007) and 4.72 t/ha reported by Abe and Akinrinola (2015) for open pollinated varieties of normal endosperm tropical field maize. This observed yield potential reflects the great prospects that abound for sweet corn production in Nigeria. The findings of this study confirmed previous reports (Cartea et al., 1996; Malvar et al., 1997, 2001; Tracy, 2001; Butrón et al., 2008; Santos et al., 2014; Entringer et al., 2017) on the potential of tropical field maize genotypes in the improvement of sweet corn.
CONCLUSION
Significant genotypic differences were observed among the populations studied. Most of the derived sh2 populations exhibited significantly superior performance relative to the sh2 donor population for most of the traits. The results of the present study indicated that new super-sweet corn populations different from the donor parent could be developed by converting field corn genotypes into super-sweet corn through backcrossing and selection for the sweet corn trait. The observed genetic differences could be exploited in further breeding programmes. The superior populations could be further improved and used as base populations for inbred line development.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
The authors wish to thank Professor V.O. Adetimirin of the Department of Agronomy, University of Ibadan for generously providing the sh2 donor population used in this study. The normal endosperm tropical field maize genotypes were sourced from the Maize Improvement Programme of IITA, Ibadan. This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
REFERENCES
|
Abe A, Akinrinola TB (2015). Green ear yield potential of tropical field maize maize at two levels of nitrogen fertiliser application in Ibadan, Nigeria. Ibadan Journal of Agricultural Research 11(1):201-209. |
|
|
Adetimirin VO (2008). Stand establishment and early field vigour variation in a tropicalised shrunken-2 maize population. Field Crops Research 108:143-149. |
|
|
Brewbaker JL (1977). 'Hawaiian superweet #9' corn. HortScience 12:355-356. |
|
|
Butrón A, Álvarez A, Revilla P, Malvar RA, Rodriguez VM, Galarreta JIR, Ordás A (2008). Agronomic performance of sweetcorn populations derived from crosses between sweetcorn and field corn. Journal of Agricultural Research 6:378-384. |
|
|
Cartea ME, Malvar RA, Revilla P, Ordás A (1996). Improvement of early vigor and adaptation of sweet corn to European Atlantic coast with open-pollinated field corn populations. Maydica 41:119-125. |
|
|
Dewanto V, Wu X, Liu RH (2002). Processed sweet corn has higher antioxidant activity. Journal of Agriculture and Food Chemistry 50(14):4959-4964. |
|
|
Entringer GC, Vettorazzi JCF, Crevelari JA, Durães NNL, Catarina RS, Pereira MG (2017). Super-sweet corn breeding by backcross: A new choice for the Brazilian market. Brazilian Journal of Agriculture 92(1):12-26. |
|
|
FAO (2016). Food and Agriculture Organization of the United Nations statistical database, |
|
|
Kim SK, Ajala SO (1996). Combining ability of tropical maize germplasm in West Africa II. Tropical vs. Temperate × Tropical origins. Maydica 41:135-141. |
|
|
Kim SK, Adetimirin VO, Yoon ST, Adepoju MA, Gbadamosi BA (2007). Green-maize potential of hybrid and open pollinated cultivars at varying levels of applied nitrogen: relationships with grain yield. Tropical Science 47(4):149-158. |
|
|
Kim SK, Efron Y, Khadr F, Mareck J, Fajemisin J (1987). Registration of 16 maize streak virus resistant tropical maize parental inbred lines. Crop Science 27:824-825. |
|
|
Lee SS, Yun SH, Kim JH (1999). Sugars, soluble solids, and flavour of sweet, super-sweet, and waxy corns during grain filling. Korean Journal of Crop Science 44(3):267-272. |
|
|
Lertrat K, Pulam T (2007). Breeding for increased sweetness in sweet corn. International Journal of Plant Breeding 1(1):27-30. |
|
|
Lizaso JI, Boote KJ, Cherr CM, Scholberg JMS, Casanova JJ, Judge J, Jones JW, Hoogenboom G (2007). Developing a Sweet Corn Simulation Model to Predict Fresh Market Yield and Quality of Ears. Journal of American Society of Horticultural Science 132(3):415-422. |
|
|
Malvar RA, Cartea ME, Revilla P, Butrón A, Ordás A (2001). Checking performance of field corn inbreds as donors of favourable alleles to improve early vigor and adaptation of sweet corn hybrids to European conditions. Maydica 46:187-193. |
|
|
Malvar RA, Revilla P, Cartea ME, Ordás A (1997). Field corn inbreds to improve sweet corn hybrids for early vigor and adaptation to European conditions. Maydica 42:247-255. |
|
|
Ogunbodede BA (1999). Green maize production in Nigeria in the new millennium- Prospects and Problems. Genetics and food security in Nigeria in the twenty-first century. Olaoye, G. and Ladipo, D.O. Eds. Genetics Society of Nigeria special publication pp. 33-37. |
|
|
Osayintola OJ, Mareck JH, Akingbala JO (1992). Effect of time of harvest and variety on the quality of boiled green field maize (Zea mays L.). Tropical Science 32:369-376. |
|
|
Qi X, Zhao Y, Jiang L, Cui Y, Wang Y, Liu B (2009). QTL analysis of kernel soluble sugar content in super-sweet corn. African Journal of Biotechnology 8:6913-6917. |
|
|
Santos PHAD, Pereira MG, Trindade RS, Silva da Cunha K, Entringer GC, Vettorazzi JCF (2014). Agronomic performance of super-sweet corn genotypes in the north of Rio de Janeiro Crop Breeding and Applied Biotechnology 14:8-14. |
|
|
SAS Institute Inc. (2003). SAS/STAT user's guide, version 9.1.3. SAS Institute Inc., Cary, NC, USA. |
|
|
Teixeira FF, Miranda RA, Paes MCD, Souza SM, Gama EEG (2013). Melhoramento do milho doce. Sete Lagoas: Embrapa Milho e Sorgo. 32 p |
|
|
Tracy WF (1997). History, genetics, and breeding of super-sweet (shrunken2) sweet corn. Plant Breeding Reviews14:189-236. |
|
|
Tracy WF (2001). Sweet corn. Speciality corns. Hallauer, A.R. Ed. 2nd ed. CRC Press, Boca Raton, Florida. pp. 155-197. |
|
|
Tracy WF, Whitt SR, Buckler ES (2006). Recurrent mutation and genome evolution: example of Sugary1 and the origin of sweet maize. Crop Science 46:1-7. |
|
|
U.S. Department of Agriculture (USDA) (2017). Sweet corn data. |
|
|
Yousef GG, Juvik JA (2002). Enhancement of seedling emergence in sweet corn by marker-assisted backcrossing of beneficial QTL. Crop Science 42(1):96-104. |
|
|
Zan GH, Brewbaker JL (1999). Seed quality of isogenic endosperm mutants in sweet corn. Maydica 44:271-277. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


