Full Length Research Paper

ABSTRACT
Nitrogen is essential due to its function in plant growth and development. It is a component of DNA and proteins which are the building blocks of life. Nitrogen is one of the most limiting factors for crop production. The need to meet the huge nitrogen requirement has necessitated the use of synthetic fertilizer. However, its continued application affects soil health, environment, and agricultural sustainability. Identifying breeding lines and developing new varieties that can efficiently fix atmospheric nitrogen and integrating these varieties into farming systems is one of the best alternatives. Field experiment was conducted at the Savanna Agricultural Research Institute research fields using 20 elite soybean lines under rain fed conditions. The objective was to select soybean (Glycine max) lines for improved nitrogen fixation, water use efficiency and grain yield. Randomized complete block design was used with three replications. Data collected were the amount of nitrogen fixed, nitrogen derived from the atmosphere, and grain yield. Genotypes showed significant variability in the amount of N-fixed, percent nitrogen derived from the atmosphere and grain yield. The mean symbiotic N contribution by the genotypes ranged from 53.6 kg/ha – 370.5 kg/ha. Grain yield was significantly different among the genotypes. Five genotypes showed superior performance for N-fixed and grain yield. There was high heritability, Phenotypic Coefficient of Variation and Genotypic Coefficient of Variation for the selected traits, which is required in breeding program for crop improvement. Genotypes should be re-evaluated in multi-locations to confirm performance and stability for release as varieties.
Key words: Glycine max, genotypes, heritability, genotypic coefficient of variation, phenotypic coefficient of variation.
INTRODUCTION
The cultivated soybean (Glycine max L.) is a worldwide legume crop that has long been regarded as one of the most widely traded commodities. It thrives in a variety of conditions, including tropical, subtropical, and temperate climates (Saryoko et al., 2017). Soybean seeds are high in dietary protein, oil, and calories. Soybean meal is one of the most important ingredients in livestock and aquaculture feeds (Hartman et al., 2011). Its production is crucial in Ghana for alleviating hunger and augmenting the costly source of animal protein. Soybean is a comparatively recent crop in Ghana, and it is mostly grown by smallholder farmers under rain-fed conditions. However, because of its importance in the rural economy of many farm households, its diversified uses and its potential of becoming a cash crop, it has received a lot of attention, particularly in the Northern Region (Adjei-Nsiah et al., 2018). The high demand for soybean requires the cultivation of high-yielding improved varieties. In West Africa's Guinea Savanna, low soil fertility, especially nitrogen (N) and phosphorous (P), the use of unimproved cultivars and low input use are the key reasons of low agricultural output (Ahiabor, 2011; Dakora et al., 1987). To enhance yields, farmers in this region frequently use inorganic fertilizers, which are costly and pollute the air and water.
There is a need for alternative means of ensuring the availability of nitrogen and employing biological nitrogen fixation seems a logical option. Soybean's root nodules are useful in fixing atmospheric nitrogen into the soil by developing a symbiotic relationship with the soil bacteria, collectively known as rhizobia. Nitrogen is a vital limiting ingredient for plant development and production, as well as one of the most significant nutrients for crop growth (Kaur et al., 2017; Mikkelsen and Hartz, 2008). Symbiotic nitrogen fixation by legume crops is therefore beneficial in farming systems (Dakora et al., 1987; Kermah et al., 2018; Yusuf et al., 2009). This symbiosis is facilitated by a complex chemical signaling process between the host legume plant and rhizobia bacteria. Grain legumes can help restore soil fertility by fixing atmospheric nitrogen (N2) while also providing protein-rich grains for family meals and nourishment (Cassman, 1993; Kermah et al., 2018). The selection of plant varieties for superior nitrogen-fixing ability may reduce the reliance on nitrogenous fertilizers and maintain soil nitrogen in the farming system. Depending on a variety of conditions, including available soil N and the existence of sufficient rhizobia numbers with high symbiotic efficiency, the amount of N-fixed in nodulated legumes varies greatly (Abaidoo et al., 2007). The process of biological nitrogen fixation is greatly affected by factors such as soil pH, soil water, available nitrogen content and temperature (Liu et al., 2011). Also, essential nutrients such as potassium and phosphorus when present in very low levels also directly or indirectly affect nodule growth and the metabolic activities of the nitrogenase (Khosro, 2012). Research findings by Lammel et al. (2018)reported that low and high soil pH can indirectly affect crop growth and development by affecting the availability of nutrients needed by crop for its growth and development. Grain legumes have been found to fix between 15 and 201 kg N ha-1 per season in the West African Guinea savanna (Dakora et al., 1987; Oteng-Frimpong and Dakora, 2018; Sanginga et al., 2002). Groundnut (Yusuf et al., 2009), soybean (Sanginga et al., 2002)and cowpea (Dakora et al., 1987)have been shown to contribute up to 48 kg N ha-1, 43 kg N ha-1 and 125 kg N ha-1, respectively to the soil. The yearly N2 fixation by soybean is estimated to be 5.7, 4.6, and 3.4 Tg in the United States, Brazil, and Argentina, respectively (Herridge et al., 2008). Some researches in Ghana have looked at soybean N2 fixation and N contribution to agricultural systems (Dakora et al., 1987; Adjei-Nsiah et al., 2018). However, the 15N natural abundance approach was used to assess a small number of genotypes. The goal of this study was to use the N difference technique to measure symbiotic N2 fixation in 25 soybean genotypes in the Guinea Savanna of Ghana. Evaluating a wide number of soybean genotypes for symbiotic N nutrition could lead to the identification of high N fixing genotypes with higher growth and grain yield for breeding purposes. Such genotypes have the potential to boost soybean production in the Guinea Savanna agro-ecology while also enhancing soil fertility without the use of nitrogen fertilizers.
MATERIALS AND METHODS
Experimental materials
The soybean lines used in this study were developed by the soybean improvement program section of the CSIR-SARI using the locally commercialized varieties developed by the said institution and some lines from the Illinois University/USDA. The list of these genotypes and their parents (Crosses) are presented in Table 1.
Site
The study was carried out at the experimental fields of the CSIR-Savanna Agricultural Research Institute at Nyankpala (9° 23' 54.08" N; 0° 58' 58.57" W, 102 m asl), in the Guinea Savannah zone of Northern Ghana during the 2020 cropping season. The site has a unimodal annual rainfall which ranges between 900 and 1100 mm and usually lasts from May to October. The texture of the soil in this site is sandy loam. The average monthly minimum and maximum temperatures in this site are 23 and 35°C, respectively. The soils are named Nyankpala series and classified under savannah Ochrosols, Plinthic luvisols (Vaccari and Panza, 2001).
Experimental design
The experimental design was Randomized Complete Block with three replicate plots per genotype. Plot size was 2.4 × 5 m with each plot containing 4 rows. The genotypes were sown without rhizobium inoculation. The spacing between rows and within plants was 60 and 5 cm, respectively. The treatment consisted of 20 advanced breeding lines and 5 checks (Parents).
Method used for soil sampling and analysis
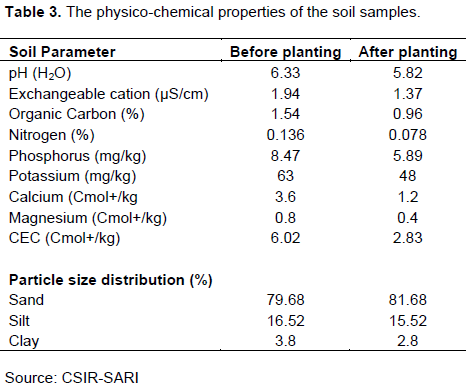
The field had been ploughed and harrowed. Planting was done in the second week of July. Sub-samples were taken in a diagonal manner from 0 to 20 cm depth on the field with a soil auger before planting and during harvesting. The extracted sub-samples were then combined and air-dried to generate the composite sample. The composite soil sample was then transferred to the laboratory and sieved for the physico-chemical properties of the soil in the experimental area using a 1 mm size mesh sieve.
Measurement of N2 fixation
In this study, the N difference technique was employed to quantify the amount of N2 fixed and N contribution by the genotypes. This technique compares the total N of the N2 - fixing species (genotypes) with that of a neighboring non N2 -fixing species with the assumption that the difference between the two is due to N2 fixation (Unkovich et al., 2008). Total N in the shoots of both the soybean and maize was analyzed by using the Kjeldahl procedure.
The following formulas were used to calculate the amount of N-fixed, N derived from the atmosphere, and N fixed (kg/ha).
Amount of N2- fixed = N legume – N maize
Soil N uptake = Total N - N2-fixed
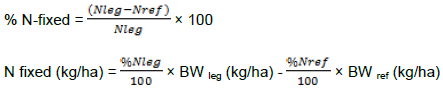
where %Nleg = percent of N-fixed by the legume, %Nref = percent of N-fixed by the non-legume (maize), and BW = biomass weight.
Data collection and statistical analysis
The data collected include days to 50% flowering, nodules per plant and nodule dry weight per plant, shoot biomass, photosynthetically active radiation (PAR), leaf stomata conductance, leaf area index, leaf transpiration rate, photosynthetic rate, height at maturity, pod clearance and number of pods per plant, plants harvested, days to maturity, grain yield per plot and one hundred seed weight.
All data collected were subjected to analysis of variance (ANOVA) using Genstat Statistical Package, and treatment means were compared using Fisher test Least Significance Difference (LSD) at 5% probability level.
Heritability estimate
The variance components were analyzed using the restricted maximum likelihood method in the lme4 function in the R package. Heritability estimates were calculated using the following formula by Burton (1952) and Sharma (1988) as cited by Ene et al. (2016)on the population heritable traits to ascertain the amount of variations in the traits that are due to genetic factors as opposed by environmental factors. These genetic variance components include genotypic variance, phenotypic variance, genotypic coefficient of variation, phenotypic coefficient of variation, genetic advance, genetic advance a percentage of the mean, and K = selection intensity differential.

where Vp = phenotypic variance, Vg = genetic variance, H2 = broad sense heritability, and K = selection intensity differential.
RESULTS
Grain yield
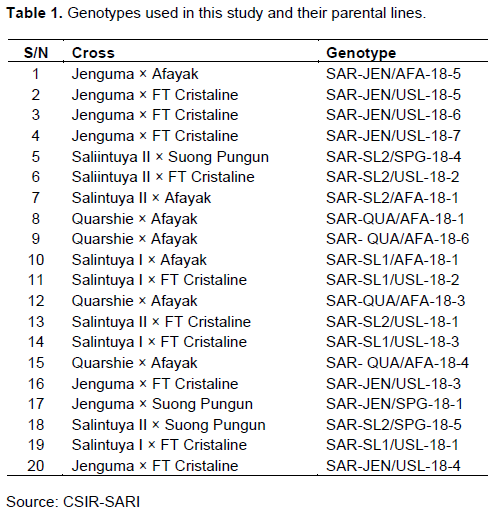
The results of the grain yield are shown in Figure 1. The genotypes showed significant differences (P = 0.03). The highest grain yield (1502 kg/ha) was obtained by genotype SAR-SL2/SPG-18-4, while the lowest grain yield (811 kg/ha) was reported by genotype SAR-JEN/USL-18-6. Apart from twelve of the genotypes that produced grain yield significantly lower than the genotype SAR-SL2/SPG-18-4, all other treatment means were not significantly different from the genotype SAR-SL2/SPG-18-4.
One hundred seed weight
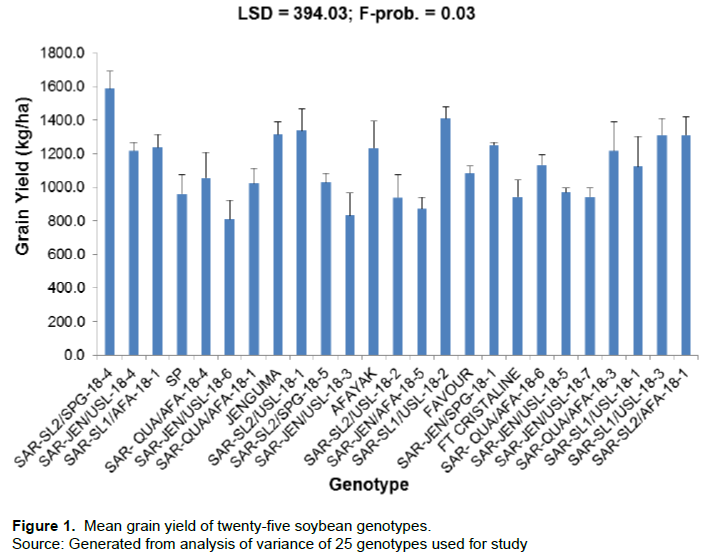
The result for 100 seed weight is as shown in Figure 2. There were significant differences (p = 0.001) among the genotypes. Genotype SAR-SL2/SPG-18-4 recorded the highest 100 seed weight (14 g), while genotype SAR-JEN/AFA-18-5 recorded the lowest seed weight (9.3 g). One hundred seed weight for genotype SAR-SL2/SPG-18-4 was significantly higher than eighteen of the genotypes. However, there was no significant difference between the genotype SAR-SL2/SPG-18-4 and all the other genotypes. Genotype SAR-JEN/AFA-18-5 was significantly lower than only eighteen of the treatments but there was no significant difference between its mean and that for six of the genotypes.
Plant stand at harvest
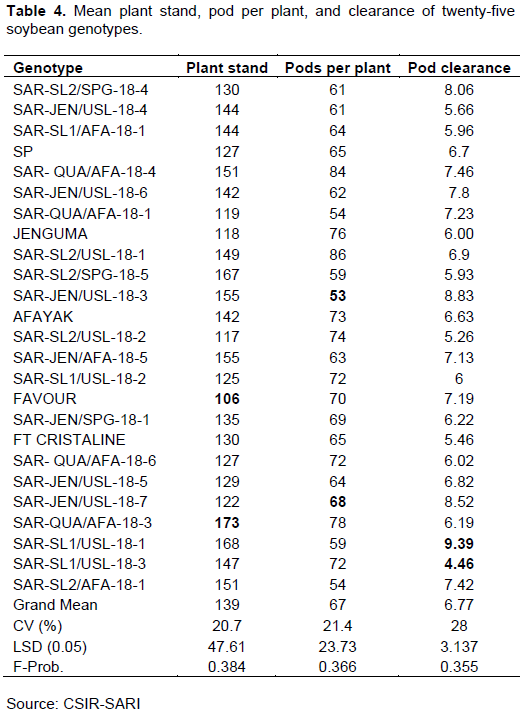
The analysis of variance revealed that there was no significant difference in plant stand among the genotypes recorded by genotype SAR-QUA/AFA-18-3, while the lowest was recorded by variety Favour (Table 4).
Pods per plant and pod clearance
There was no significant difference (P = 0.366) in the mean values of all genotypes for the number of pods per plant. The genotype SAR-SL2/USL-18-1 produced the most pods per plant (86), followed by genotypes SAR-QUA/AFA-18-4 (84) and SAR-QUA/AFA-18-3 (78). The lowest pod per plant was produced by genotype SAR-JEN/USL-18-3 (53) (Table 2). Similar to pod per plant, there were no significant mean differences (P = 0.355) in the pod clearance among all the genotypes. The mean pod clearance was 6.77 cm. Genotype SAR-SL1/USL-18-1 recorded the highest pod clearance (9.39 cm) followed by genotype SAR-JEN/USL-18-3 (8.83 cm). The result is presented in Table 4.

Effective nodules per plant and nodule dry weight
The results showed that genotype differences did not significantly affect the number of nodules produced per plant and the nodule dry weight (Table 5). However, genotype FT Cristaline produced the highest nodule number per plant (56.5), while variety Suong Pungun produced the lowest number of nodules per plant (21.9).
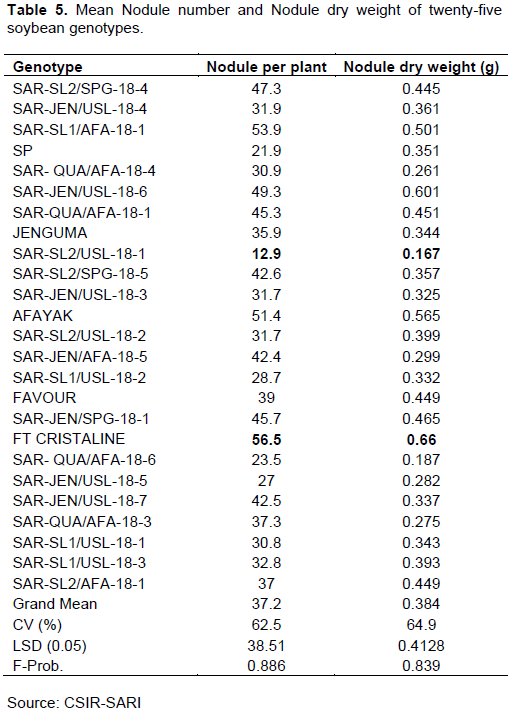
But, the nodule number for the genotype FT Cristaline was not significantly different (P = 0.886) than those for all other genotypes. The highest nodule weight per plant was recorded for genotype FT Cristaline (0.66 g) followed by genotype SAR-JEN/USL-18-6 (0.60), while the lowest was recorded for genotype SAR-SL2/USL-18-1 (0.167).
Again, the nodule weight for the genotype FT Cristaline was not significantly different (P = 0.0839) than those for the other treatments (Table 5).
Biomass weight
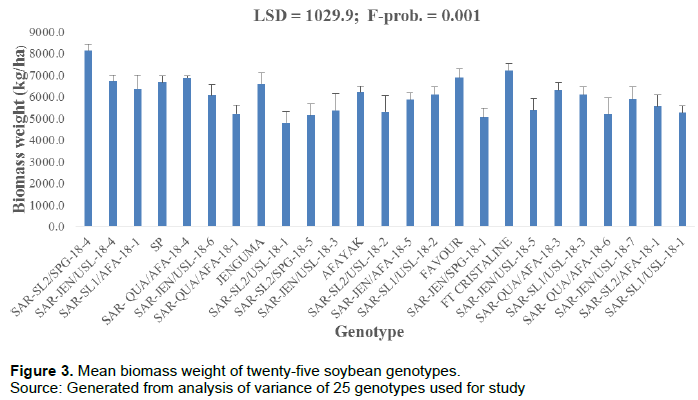
There were significant differences (p = 0.001) among the mean values of the genotypes for biomass weight when the data was subjected to analysis of variance. Genotypes, SAR-SL2/SPG-18-4 and FT Cristaline recorded the highest biomass weight (8133.3 and 7226.7 kg/ha), respectively, while genotype SAR-SL2/USL-18-1 recorded the lowest. Genotype SAR-SL2/SPG-18-4 biomass weight was significantly higher than fifteen of the genotypes but there was no significant difference between the genotype SAR-SL2/SPG-18-4 and the rest of the genotypes (Figure 3).
Amount of N fixed (kg/ha)
There were significant differences (P = 0.001) in the amount of N-fixed among the genotypes (Figure 4). Favour produced the highest amount of N-fixed (370.5 kg N/ha), followed by SAR-SL2/SPG-18-4 (256.8 kg/ha) and FT Cristaline (133.3 kg N /ha), while genotype SAR-SL2/USL-18-1 recorded the lowest amount of N-fixed per hectare (53.6 kg N/ha). The N-fixed by favour was significantly different from all the other genotypes (Figure 4).
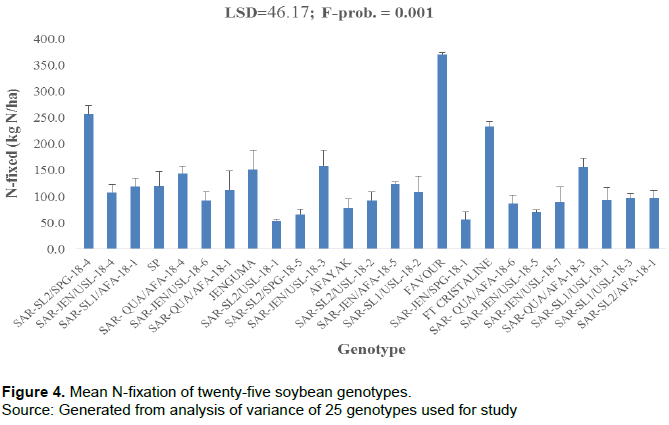
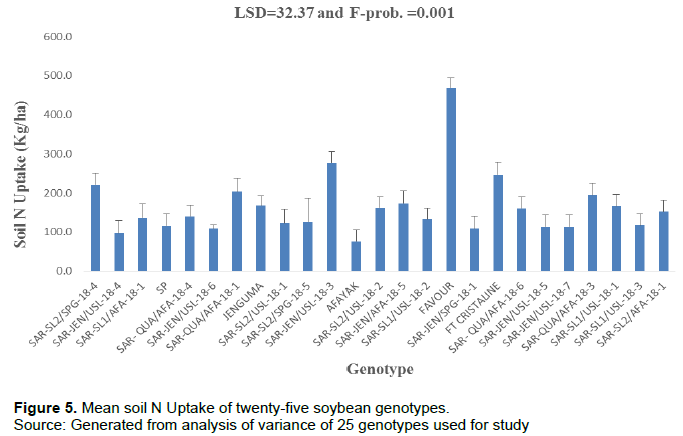
Soil N uptake (kg/ha)
There was a high significant difference (P = 0.001) in the mean values of the genotypes for N uptake. The greatest amount of soil mineral N was taken up by variety Favour (468 kg/ha) and the lowest soil N uptake was recorded by genotype SAR-SL2/USL-18-1 (34 kg/ha). The N uptake of Favour was significantly higher than all the other treatments (Figure 5). The second highest soil mineral nitrogen (N) uptake was recorded by the genotype, SAR-JEN/USL-18-3 (277 kg/ha). However, there was no significant difference between the mean N uptake of the genotype SAR-JEN/USL-18-3 and FT Cristaline.
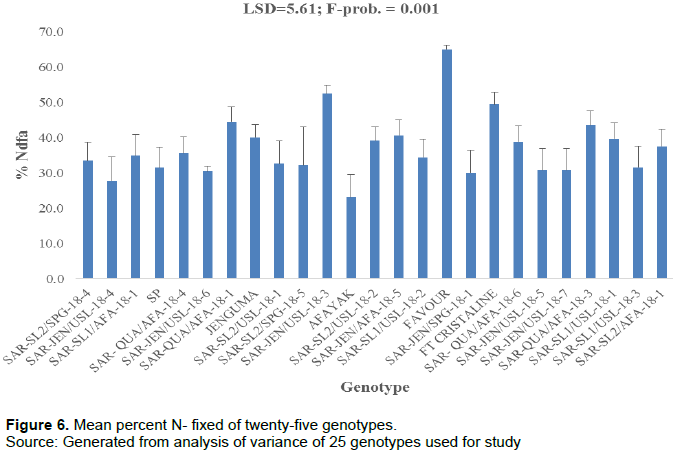
Percent of N derived from the atmosphere (%NDFA)
The ANOVA revealed that there were significant differences (P = 0.001) among the genotypes for their mean percent N derived from the atmosphere (Figure 6). The percent mean NDFA by genotype Favour was significantly different from all the other genotypes. Genotype Afayak recorded the least percent NDFA.
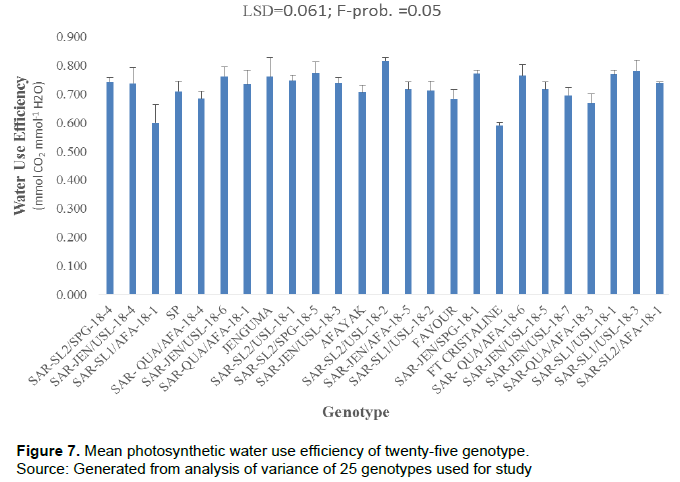
Photosynthetic water use efficiency
There were significant differences (P = 0.05) among the genotypes mean for photosynthetic water-use efficiency at the time of data collection. The genotype SAR-SL2/USL-18-2 showed the highest photosynthetic water use efficiency rate at the time of data collection, while genotype FT Cristaline showed the lowest water use efficiency. However, there was no statistically significant difference between the mean photosynthetic water use efficiency of genotype SAR-SL2/USL-18-2 and six of the genotypes, JENGUMA, SAR-SL1/USL-18-3, SAR-SL1/USL-18-1, SAR-QUA/AFA-18-6, SAR-SL2/SPG-18-5, and SAR-JEN/USL-18-6 (Figure 7).
Estimates of heritability and genetic parameters
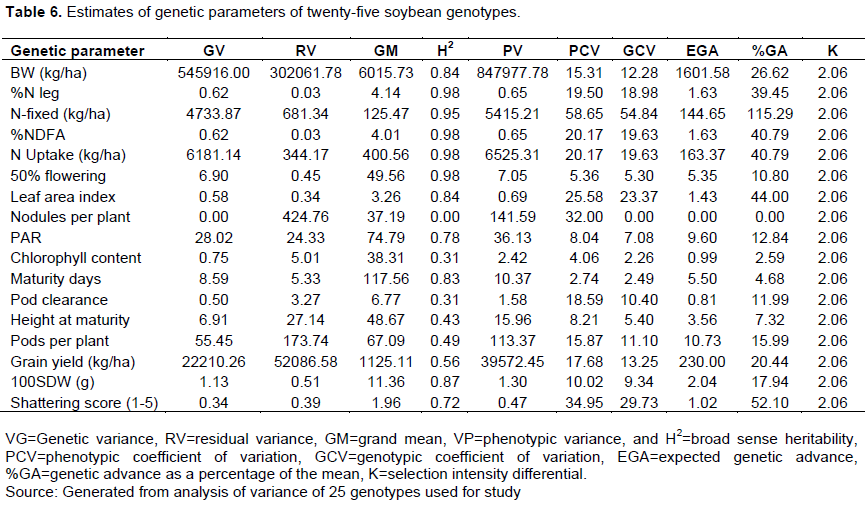
The estimate of heritability of the variance components as presented in Table 6 showed the traits 50% flowering, percent nitrogen, nitrogen derived from the atmosphere, and nitrogen uptake by the genotypes recording the highest heritability of 0.98 or 98% each, followed by the amount of nitrogen fixed (0.95 or 95%), and 100 seed weight (0.87 or 87%). The lowest broad sense heritability was recorded by the trait, nodule number (0.00002 or 0.002%). The genotypic variance in all the traits observed ranged from 0.00002 (Nodule per plant) to 545916.00 (Shoot biomass weight). In this study, it was observed that the phenotypic coefficient of variation values was higher than the genotypic coefficient variation values. However, the differences between these two were not significant. The phenotypic variation values ranged from 0.47 (Shattering score) to 847977.78 (Shoot biomass). The highest genotypic coefficient of variation was observed in trait amount of N-fixed (54.84) while the lowest was recorded by nodule number per plant (0.002). The phenotypic coefficient of variation values ranges from 2.74 to 58.65. The highest expected genetic gain was 1601.58 and the selection intensity differential value was 2.06. The estimated heritability values were very high in most of the parameters with a significant variation between the parameters.
DISCUSSION
The analysis of soil samples showed poor fertility levels of the soils in the Guinea Savanna of West Africa, particularly, low nitrogen (N) and phosphorus (P). Even though there was no Rhizobium inoculation, there was a substantial amount of nodulation exhibited by all the soybean genotypes. This result suggests that there were indigenous bacteria, Bradyrhizobia available in the soil. A report by Delamuta et al. (2013)suggested that Bradyrhizobium species is a native of the tropics and is the main symbiont of cowpea and many other legumes such as groundnut, Bambara groundnut, and Soybean. Although there were no significant differences among the genotypes for both the number of nodules produced and nodule dry weight, genotype FT Cristaline produced the highest nodule number per plant, while genotype SuongPungun produced the lowest number of nodules per plant. Similar to nodule number, the highest nodule weight per plant was recorded for genotype FT Cristaline, while the lowest was recorded for genotype SAR-SL2/USL-18-1 (Table 3). It was interesting to observe that genotype FT Cristaline which produced the highest number of nodules was the same genotype that recorded the highest nodule dry weight. This result was in contrast with what Sarkodie-Addo et al. (2007)reported. They observed significant differences in both the number of nodules produced per plant and nodule dry weight with a negative correlation between the two when medium maturing soybean lines were evaluated for their nitrogen fixation potentials. Also, significant differences in nodule production and nodule dry weight among varieties have been reported in legumes such as cowpea by Egbe and Egbo (2011)and groundnut by Moji et al. (2020).
The results showed significant differences in biomass weight and N fixation (P = 0.001) concluded that genotypes. These findings were similar to what Belane and Dakora (2010)reported. They suggested that a higher amount of nitrogen fixed contributed to increased performance in terms of growth and grain yield when compared with low nitrogen-fixing genotypes. The relatively significant performance exhibited by the genotypes in terms of shoot biomass produced can be attributed to their ability to fix a greater amount of nitrogen. The positive correlation between shoot biomass produced per hectare and the amount of N fixed per hectare in this study confirmed this report.
Grain yield and N fixed results showed significant differences among genotypes. Thus, genotypic effect was responsible for the significant differences observed since all the genotypes were planted under the same environment. Similar research findings have been reported by Sarkodie-Addo et al. (2007)in Soybean, by Oteng-Frimpong and Dakora (2018)in groundnut, and Berchie et al. (2010)in Bambara groundnut. Although genotype Favour was the treatment that recorded the highest amount of N-fixed, its grain yield was significantly lower than the genotype SAR-SL2/SPG-18-4 which produced the largest grain yield. The grain yield of the genotype Favour was even lower than the genotype SAR-SL2/USL-18-1 which fixed the lowest amount of N. The negative correlation observed from this result could be due to the inability of the genotype Favour to translate its greatest amount of N-fixed into a grain yield production. This result was in contrast with what Samago et al. (2018) and Sarkodie-Addo et al. (2007)reported. They observed a positive correlation between N-fixed and grain yield in common bean (Phaseolus vulgaris L.) and soybean (G. max L.) respectively. Samago et al. (2018) observed this when they assessed common bean varieties′ response to Rhizobium inoculation and phosphorus application. Sarkodie-Addo et al. (2007) recorded this when they evaluated soybean medium maturing lines for their nitrogen fixation potentials. Except for a few features, heritability estimates were generally high in most of the traits observed.
For variables such as 50% flowering, percent nitrogen, NDFA, N uptake, N fixed, and 100 seed weight, there was substantial broad-sense heritability observed among the genotypes, implying that the phenotypes strongly reflect the genotypes. The findings of this study matched those of other studies (Datta et al., 2005; Jain et al., 2018). Also, this study observed a high PCV and GCV which are required in a breeding program for crop improvement. Although the PCV was observed to be higher than the GCV, the difference was not significant. This result was in line with what Baraskar et al. (2014)reported. He observed higher values for PCV than the GCV when he carried out a study on genetic variability, heritability and genetic advance in soybean. This suggests the role the environment played in the expression of the characters. The highest PCV and GCV were observed by traits N-fixed (kg/ha) and shattering score. The N-fixed trait was also among the traits with the highest heritability values which probably will make it ideal for selection. GCV and genetic advances, which are important genetic parameters for selection was observed to be high for shattering, which suggest that the trait is core to be considered in a breeding program.
CONCLUSION
Even though there were no significant differences among the genotypes for nodulation, the amount of nitrogen fixed and the amount of grain produced were both notably different. This significant variation could be attributed to genotypic differences. There was a positive correlation between nitrogen fixation and soybean biomass production. Genotypes that fixed the most nitrogen did not produce the highest grain yield. There were significant differences in water use efficiency. Genotype SAR-SL2/USL-18-2 recorded the highest water use efficiency without a simultaneous decrease in photosynthetic rate in biomass production. Genotypes that showed increased performance for water use efficiency with contrast performance in fixing nitrogen can be used as breeding lines in breeding programs that have the objective of enhancing water-use efficiency in high nitrogen-fixing genotypes.
Genotypes that perform creditably well in terms of grain yield could be evaluated in multi-location and if perform the same, should be selected for release as variety. Genotypes with high nitrogen fixation could be used in integrated farming system schemes to improve soil fertility which will indirectly reduce the cost of production and also maintain beneficial eco-systems.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Abaidoo RC, Keyser HH, Singleton PW, Dashiell KE, Sanginga N (2007). Population size, distribution, and symbiotic characteristics of indigenous Bradyrhizobium spp. that nodulate TGx soybean genotypes in Africa. Applied Soil Ecology 35(1):57-67. |
|
|
Adjei-Nsiah S., Alabi BU, Ahiakpa JK, Kanampiu F (2018). Response of grain legumes to phosphorus application in the Guinea savanna agro-ecological zones of Ghana. Agronomy Journal 110(3):1089-1096. |
|
|
Ahiabor B (2011). Innovations as Key to the Green Revolution in Africa. Innovations as Key to the Green Revolution in Africa, September. |
|
|
Baraskar VV, Kachhadia VH, VachhanI JH, Barad HR, Patel MB, Darwankar MS (2014). Genetic variability, heritability and genetic advance in soybean [Glycine max (L.) Merrill]. Electronic Journal of Plant Breeding 5(4):802-806. |
|
|
Belane AK, Dakora FD (2010). Symbiotic N2 fixation in 30 field-grown cowpea (Vigna unguiculata L. Walp.) genotypes in the Upper West Region of Ghana measured using 15N natural abundance. Biology and Fertility of Soils 46(2):191-198. |
|
|
Berchie JN, Sarkodie-Addo J, Adu-Dapaah H, Agyemang A., Addy S, Asare E, Donkor J(2010). Yield evaluation of three early maturing bambara groundnut (Vigna subterranea L. Verdc) Landraces at the CSIR-Crops Research Institute, Fumesua-Kumasi, Ghana. Journal of Agronomy 9(4):175-179. |
|
|
Cassman KG (1993). Nitrogen fixation in tropical cropping systems. In Field Crops Research 34(2):230-232. |
|
|
Dakora FD, Aboyinga RA Mahama AJ (1987). Assessment of N2 fixation in groundnut (Arachis hypogaea L.) and cowpea (Vigna unguiculata L. Walp) and their relative N contribution to a succeeding maize crop in Northern Ghana. Mircen Journal of Applied Microbiology and Biotechnology 3(4):389-399. |
|
|
Datta A, Hossain M, Rahman L (2005).Genetic parameters, inter-relationship and path-coefficient analysis in soybean (Glycine max (L.) Merrill). Journal of the Bangladesh Agricultural University 3(2):225-230. https://doi.org/10.22004/ag.econ.276479 |
|
|
Delamuta JRM, Ribeiro RA, Ormeño-Orrillo E Melo IS, Martínez-Romero E, Hungria M (2013). Polyphasic evidence supporting the reclassification of Bradyrhizobium japonicum group Ia strains as Bradyrhizobium diazoefficiens sp. nov. International Journal of Systematic and Evolutionary Microbiology 63(9):3342-3351. |
|
|
Egbe O, Egbo C (2011). Nodulation , nitrogen fixation and harvest index of extra- short- and short-duration cowpea varieties intercropped with maize at Otobi , Benue state , Nigeria. Journal of Animal and Plant Science 10(3):1315-1324. |
|
|
Ene CO, Ogbonna PE, Agbo CU, Chukwudi UP (2016). Studies of phenotypic and genotypic variation in sixteen cucumber genotypes. Chilean Journal of Agricultural Research 76(3):307-313. |
|
|
Hartman GL, West ED, Herman TK (2011). Crops that feed the World 2. Soybean-worldwide production, use, and constraints caused by pathogens and pests. Food Security 3(1):5-17. |
|
|
Herridge DF. Peoples MB, Boddey RM (2008). Global inputs of biological nitrogen fixation in agricultural systems. Plant and Soil 311(1-2):1-18. |
|
|
Jain RK, Joshi A, Chaudhary HR, Dashora A, Khatik CL (2018). Study on genetic variability, heritability and genetic advance in soybean [Glycine max (L.) Merrill]. Legume Research 41(4):532-536. |
|
|
Kaur G, Serson WR, Orlowski JM, McCoy JM, Golden BR, Bellaloui N (2017). Nitrogen sources and rates affect soybean seed composition in Mississippi. Agronomy 7(4):1-10. |
|
|
Kermah M, Franke AC, Adjei-Nsiah S, Ahiabor BDK, Abaidoo RC, Giller KE (2018). N2-fixation and N contribution by grain legumes under different soil fertility status and cropping systems in the Guinea savanna of northern Ghana. Agriculture, Ecosystems and Environment 261(2017):201-210. |
|
|
Khosro M (2012). Effective factors on biological nitrogen fixation. African Journal of Agricultural Research 7(12):1782-1788. |
|
|
Lammel DR, Barth G, Ovaskainen O, Cruz LM, Zanatta JA, Ryo M, de Souza EM, Pedrosa FO(2018). Direct and indirect effects of a pH gradient bring insights into the mechanisms driving prokaryotic community structures. Microbiome 6(1):7-9. |
|
|
Liu Y, Wu L, Baddeley JA, Watson CA (2011). Models of biological nitrogen fixation of legumes. A review. Agronomy for Sustainable Development 31(1):155-172. |
|
|
Mikkelsen R, Hartz T (2008). Nitrogen Sources for Organic Crop Production. America N, 1-4. %3E |
|
|
Moji KT, Jibrin MJ, Umar FG, Abubakar UD (2020). Genotypic Effect of Groundnut on Nodulation, N Fixation and N Balance in The Two Savannahs of Nigeria. Nigerian Journal of Soil Science |
|
|
Oteng-Frimpong R., Dakora FD (2018). Selecting elite groundnut (Arachis hypogaea L) genotypes for symbiotic N nutrition, water-use efficiency and pod yield at three field sites, using 15 N and 13 C natural abundance. Symbiosis 75(3):229-243. |
|
|
Samago TY, Anniye EW, Dakora FD (2018). Grain yield of common bean (Phaseolus vulgaris L.) varieties is markedly increased by rhizobial inoculation and phosphorus application in Ethiopia. Symbiosis 75(3): 245-255. |
|
|
Sanginga N, Okogun J, Vanlauwe B, Dashiell K (2002). The contribution of nitrogen by promiscuous soybeans to maize based cropping the moist savanna of Nigeria. Plant and Soil 241(11):223-231. |
|
|
Sarkodie-Addo J, Adu-Dapaah H, Ewusi-Mensah N, Asare E(2007). Evaluation of medium-maturing soybean (Glycine max (L) Merrill) lines for their nitrogen fixation potentials. Journal of Science and Technology (Ghana) 26(2). |
|
|
Saryoko A, Homma K, Lubis I, Shiraiwa T (2017). Plant development and yield components under a tropical environment in soybean cultivars with temperate and tropical origins. Plant Production Science 20(4):375-383. |
|
|
Unkovich M., Herridge D, Peoples M, Cadisch G, Boddey B, Giller K, Alves B, Chalk P (2008). Measuring plant-associated nitrogen fixation in agricultural systems. Measuring Plant-Associated Nitrogen Fxation in Agricultural Systems 132-188. |
|
|
Vaccari F, Panza GF (2001). Abdus Salam International Centre for Theoretical Physic pp. 1-24. |
|
|
Yusuf AA, Iwuafor ENO, Abaidoo RC, Olufajo OO, Sanginga N (2009). Grain legume rotation benefits to maize in the northern Guinea savanna of Nigeria: Fixed-nitrogen versus other rotation effects. Nutrient Cycling in Agroecosystems 84(2):129-139. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


