Full Length Research Paper

ABSTRACT
Inbred line development requires information on the nature and magnitude of genetic variability and transmissibility of desired trait in source populations. In this study, the level of genetic variability, heritability and genetic advance of thirteen agronomic and fresh yield traits among twelve shrunken-2 super-sweet corn populations were evaluated. Field experiments were conducted for two years in Ibadan, Nigeria using a randomised complete block design with three replicates. Estimates of genetic variability components, broad-sense heritability and genetic advance were computed for each trait. All the traits exhibited significant genotypic differences. Genotypic variance was significant for number of marketable cobs, yield of cobs, number of cobs, number of kernel rows, husk cover, ear height and days to anthesis, while environmental variance was significant for all the traits. Phenotypic coefficients of variation were higher than the corresponding genotypic coefficients of variation for all traits. Broad-sense heritability ranged from 22.2% for anthesis-silking interval to 85.1% for husk cover. The genetic advance was high (32.7%) for husk cover, medium (12.0%) for yield of cobs and low for other traits. Genetic variability was present among the shrunken-2 super-sweet corn populations. Opportunities abound for further improvement of the populations and extraction of lines for hybrid seed production.
Key words: Broad-sense heritability, genotypic coefficient of variation, husk cover score, sweet corn, yield of marketable cobs.
INTRODUCTION
Maize is the most widely cultivated staple food crop in sub-Saharan Africa, where it contributes up to 70% of the daily caloric intake (Martin et al., 2000; FAO, 2007). It is a highly diverse grain crop with multiple applications and consists of different types, generally classified by kernel endosperm characteristics. The most common types of maize are flint, dent, flour, pop, pod, waxy and sweet. The physical appearance of each kernel type is determined by the endosperm pattern, quantity and quality. Cultivated maize varieties in West and Central Africa are mostly of the flint and dent types which have largely been improved by the use of both temperate and tropical exotic germplasm (Kim et al., 1987; Kim and Ajala, 1996).
Sweet corn (Zea mays L. var saccharata) is a highly perishable vegetable maize crop, harvested exclusively for fresh human consumption when the grains are at the milk stage with approximately 75% moisture content. It is a type of maize with kernels that are sweet as a result of high sugar content which when consumed in the immature stage, has high levels of total sugars than normal field maize, rich in fibre, minerals and certain vitamins (Tracy, 1997; Lertrat and Pulam, 2007). It has also been reported to possess significant antioxidant properties which reduce the possibilities of developing cancer and inflammations in humans (Dewanto et al., 2002). Because of these qualities, sweet corn could play a significant role in the nutrition of the peoples of West and Central Africa (Adetimirin, 2008) where fresh green maize is an important component of human diet. The difference between sweet corn and field maize arises from a mutation that influences carbohydrate biosynthesis in the endosperm. In the genome of sweet corn, at least one of the eight mutant monogenic and recessive genes preventing the conversion of sugars to starch is present (Tracy et al., 2006; Qi et al., 2009; Santos et al., 2014) resulting in the characteristic sugary taste. Of these mutants, the four most useful are shrunken-2 (sh2), brittle-2 (bt2), sugary-1 (su1), and sugary enhancer (se). While sh2 and bt2, classified as class 1 mutants (super-sweet corn), are involved in large reductions in starch and large increase in sugar, su1 and se referred to as class 2 mutants (sweet corn) occur later in the starch biosynthesis pathway and influence the types and proportion of polysaccharides stored in the endosperm (Boyer and Shannon, 1984; Tracy, 1997). The grain of matured normal field maize is known to contain about 3% sugar, while sweet and super-sweet corns contain 9 to 14% and 15 to 30% sugar, respectively (Creech, 1965; Teixeira et al., 2013). The sh2 (Yousef and Juvik, 2002) and bt2 (Brewbaker, 1977) types of sweet corn have the greatest commercial value.
Nigeria produces about 40% of the maize production in West and Central Africa (FAO, 2016). Maize is grown throughout Nigeria from the high rainfall forest of the southeast to the low rainfall Sudan Savanna of the north. Widening food preferences have resulted in increased importation of canned sweet corn to Nigeria. Great potential therefore exists for the production and commercialisation of super-sweet corn. However, sweet corn cultivars are virtually nonexistent in Nigeria. To bridge the gap created by this challenge, a broad-base temperate super-sweet sh2 maize population was introduced into the country and adapted to the prevailing tropical environmental conditions by four cycles of mass selection (Adetimirin, 2008). This adapted population was meant to serve both as an open pollinated variety, as well as, basis for inbred line development.
Super-sweet corn has a narrow genetic base (Tracy, 2001; Teixeira et al., 2013; Mahato et al., 2018) and possesses some undesirable characteristics, such as poor field emergence and seedling vigour and susceptibility to diseases and pests. In addition, tropical climatic conditions are characterized by high disease incidence, pest attack, short day length, and high temperatures. One way to improve the performance of this tropicalised population and broaden it genetic base is by crossing the sh2 maize with the more adapted field maize germplasm, which could be sources of favourable alleles for enhanced agronomic performance, as well as, increased disease and pest resistance. Previous studies (Tracy, 2001; Cartea et al., 1996; Malvar et al., 1997, 2001; Butrón et al., 2008; Entringer et al., 2017) have shown the utility of field corn in the improvement of agronomic performance of sweet corn. These studies have also indicated that field maize genotypes could differ in their ability to improve the agronomic performance and quality of sweet corn.
In order to develop hybrids, the source populations from which inbred lines are extracted for hybrid seed production should have superior qualities (Hallauer, 1990). One of such source populations are the open-pollinated varieties (Sleper and Poehlman, 2006; Hallauer et al., 2010). In this study, the estimates of variance components, heritability and genetic advance for measured traits in sh2 super-sweet corn populations derived from crosses between a tropicalised sh2 super-sweet corn population and tropical field maize genotypes were reported.
MATERIALS AND METHODS
Experimental site
The study was conducted at the experimental field of the Department of Agronomy, Faculty of Agriculture, along Parry road, University of Ibadan (7°26' N, 3°54' E), Ibadan, Nigeria.
Experimental design and layout
Twelve sh2 populations, eleven of which were derived from crosses between tropical field maize genotypes and a tropicalised sh2 donor population were evaluated over two years in a randomised complete block design with four replicates. The soil at the experimental site is sandy-loam with 20.3 g kg-1 organic carbon, 0.90 g kg-1 total nitrogen, 17.90 mg kg-1 available P (Bray-1), 0.28 cmol kg-1 K and a pH (H2O) of 6.2. The populations in each block were planted in four 5.0 m long rows spaced at 0.75 m apart. Seeds were sown at 0.50 m apart within the row. Four seeds were sown per hill and later thinned to two to give a plant population of approximately 53,333 plants per hectare. At two weeks after planting (WAP),
All agronomic and fresh ear yield data excluding flowering data collected on the two middle rows of each plot. Data on days to anthesis (DA) and days to silking (DS) were recorded as number of days from planting to when 50% of the plants in a plot shed pollen or have emerged silks, respectively. Anthesis-silking interval (ASI) was calculated as the difference between DS and DA. Plant height (PH) and ear height (EH) measured in meters were taken at fresh ear yield harvest on all plants in the two middle rows of a plot, as the average distance from the soil level to the collar of the uppermost leaf and collar of the leaf bearing the uppermost ear, respectively. Husk cover (HC) was scored on a scale of 1 to 9 (1 = husk tightly covers ear tip and extends beyond it; 9 = poor husk cover with ear tip clearly exposed). Harvesting of fresh ears was done three weeks after silking. Fresh ear yield data included: number of cobs (NC; fresh ears with husk removed) harvested per plot expressed per hectare; yield of cobs (YC; fresh ears with husks removed) recorded as total weight of cobs harvested per plot expressed in tonnes per hectare; number of marketable cobs (NMC; cobs with approximately 250 filled edible kernels) per plot expressed per hectare; yield of marketable cobs (YMC) recorded as total weight of marketable cobs per plot expressed in tonnes per hectare; number of kernel rows (KR) recorded as the average number of kernel rows of 10 top cobs; cob length (CL) measured in cm as the average length of 10 top cobs; cob diameter (CD) measured in mm using an electronic 6 in. digital calliper (Pittsburgh®, Item #47257), as the average diameter of 10 top cobs taken at the middle portion of the cob.
Data analyses
The PROC MIXED procedure in SAS (SAS version 9.1.3, SAS Institute, 2003) was used to compute variance components for all traits. In the model, populations and the interaction of populations by environment were considered random, while environments and replications within environments were fixed. Each year was considered a separate environment.
Genotypic (GCV) and phenotypic (PCV) coefficients of variation were computed for all traits according to Singh and Chaudhary (2004) using the equations:
GCV (%) = {(√ σ2g) /x} × 100
PCV (%) = {(√ σ2p) / x} × 100
where σ2g = genotypic variance, σ2p = phenotypic variance and x = grand mean for the trait.
The GCV and PCV were considered low when less than 10%, moderate when 10 to 20% and high when greater than 20% as explained by Deshmukh et al. (1986).
Broad-sense heritability and their standard errors were estimated using variance ratios as explained by Hallauer et al. (2010) using the following equations:
H2 = {σ2g/ (σ2g + σ2ge/e + σ2/re)} × 100
SE (H2) = SEσ2g/ (σ2g + σ2ge/e + σ2/re)
where σ2g = genotypic variance, σ2 = environmental variance, σ2ge = variance due to genotype by environment interaction, σ2p = phenotypic variance = σ2g + σ2ge + σ2, r = number of replicates, e = number of environments and SE = standard error.
Heritability estimates were categorized into low (less than 40%), medium (40-59%), moderately high (60-79%) and very high (80% and above) as described by Singh (2001).
Genetic advance (GA) was estimated using the formula by Singh and Chaudhary (2004):
GA = i σp h2
where i = 1.40 (selection intensity at 20.0%), σp = phenotypic standard deviation, h2 = broad-sense heritability.
Genetic advance expressed as percentage of the mean was estimated as described by Souza et al. (2009) as follows:
GA (%) = GA/x × 100
where x = grand mean of all hybrids for the trait.
The GA was considered low (less than 10%), moderate (10-20%) and high (above 20%) according to Johnson et al. (1955).
RESULTS AND DISCUSSION
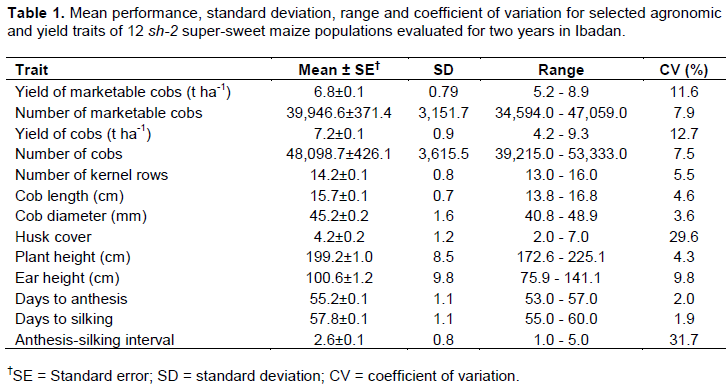
Mean performance of populations
Selection of genetically superior genotypes as well as inbred lines for hybrid seed production requires sufficient genetic variability and high heritability in the base population. The populations evaluated in this study exhibited a wide range in values for all the traits assessed (Table 1). This indicates that a sufficient range of variability in all the traits exists among the populations. The presence of variability could be a consequence of the differences in the ability of the field maize genotypes involved in the development of derived sh2 populations to improve the donor sh2 population (Butrón et al., 2008; Entringer et al., 2017) as well as a reflection of the influence of environment on the expression of the traits.
Estimates of variance components, heritability and genetic advance
Estimates of genetic variance (σ2g) were significant only for YC, NC KR, HC, EH and DA, while estimates of environmental variance (σ2e) were significant for all variables (Table 2). The estimates of genetic variances were higher than the corresponding environmental variances for all traits except PH and ASI. The EH had the highest significant genetic and environmental variances. The observed differences among the genotypes for most of the traits were therefore more due to genetic than environmental causes. The PCV estimates in this study were higher than the corresponding GCV for all traits, although the differences between values were low except for ASI. The PCV and GCV were high for HC, whereas only PCV was high for ASI. The GCV and PCV estimate for YC, as well as GCV for ASI and PCV for YMC were moderate. All other traits manifested low GCV and PCV values. The range in values of GCV and PCV were 1.43 (DS) to 26.50 (HC) and 1.75 (DS) to 30.05 (HC), respectively (Table 2). The slightly higher PCV than GCV for all traits in this study indicates that the expressions of the traits were influenced, though to a limited extent, by the environment and there is the possibility of improvement using phenotypic selection. Similar results indicating higher PCV than GCV for all traits were reported by Saleh et al. (2002), Alan et al. (2013) and Niji et al. (2018) in sweet corn as well as Maphumulo et al. (2015), Sesay et al. (2016) and Jilo et al. (2018) in field maize. The PCV for ASI was more than twice the GCV, an indication of limited chance for selection for the trait in the populations studied. Jilo et al. (2018) reported similar results for ASI. The high GCV exhibited by HC shows that the trait is less affected by environmental fluctuations, which guarantees selection progress for the trait. High GCV estimates are indicative of low amenability of the trait to environmental changes (Hefny, 2011). Emphasis on HC is therefore important in the development of cultivars from the present genetic materials. In sweet corns, good husk covering is one of the most important quality characteristics (Tracy, 1997) as complete husk covering shields of the ears from earworm attack (Lynch et al., 1999; Gardner et al., 2000). Consistent with the results of the present study, Alan et al. (2013) and Niji et al. (2018) reported low PCV and GCV estimates for DA, DS, PH, CL and CD in sweet corn.
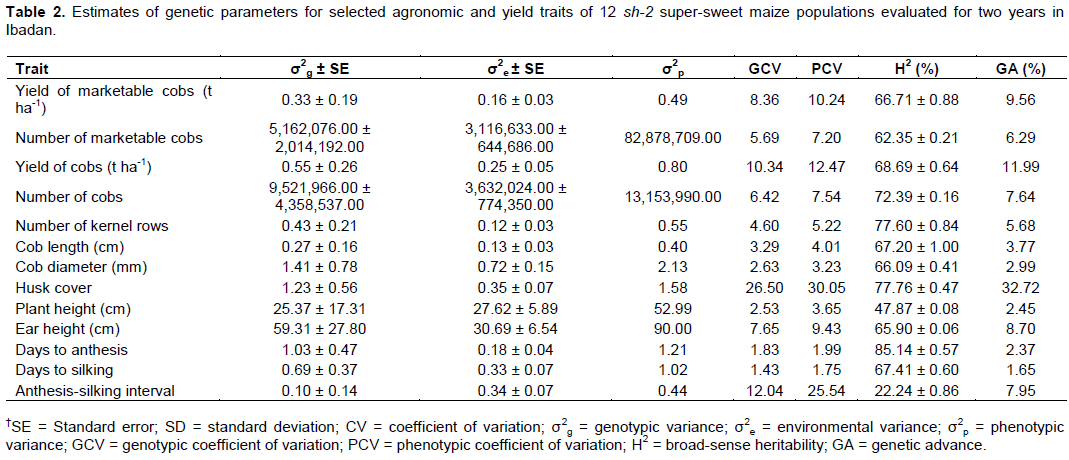
Estimates of broad-sense heritability ranged from 22.2% for ASI to 85.1% for DA. The heritability estimates were low for ASI, medium for PH, moderately high for YMC, NMC, YC, NC, KR, CL, CD, HC, EH and DS, and very high for DA (Table 2). Genetic advance expressed as a percentage of the mean ranged from 1.7 (DS) to 32.7 (HC). The YC and HC exhibited medium and high GA, respectively, while the estimates for all other variables were low (Table 2). In this study, heritability estimates were moderately high to very high for the traits considered, except PH and ASI. However, the GA expressed as a percentage of the mean accompanying these estimates were low, except for YC with medium GA and HC with high GA. This suggests that genetic control of the traits was predominantly non-additive. Johnson et al. (1955) and Jilo et al. (2018) had previously suggested the simultaneous consideration of heritability estimates and GA because high heritability may not always be associated with high GA. These traits may respond to phenotypic selection and could be improved through heterosis breeding (Bello et al., 2012; Nzuve et al. 2014). The moderately high heritability estimate for HC in this study coupled with high GA indicated that husk covering is a quality trait exhibiting additive gene action and selection for the trait would be effective. However, HC manifested high PCV suggesting a strong influence of environment on its expression.
CONCLUSIONS
Super-sweet corn has a narrow genetic base and poorly adapted to tropical environmental conditions. The performance of super-sweet corns under tropical conditions could be enhanced by crossing super-sweet corn cultivars with adapted tropical field maize germplasm. However, field maize genotypes could differ in their ability to improve the agronomic performance and quality of super-sweet corn. In this study, 11 sh2 super-sweet corn populations (developed by backcrossing between a sh2 donor population and 11 adapted field maize genotypes) along with the donor parent sh2 population were assessed to determine the nature and extent of genetic variability and transmissibility of desired trait within them. This study revealed the existence of a wide range of genetic variability in the sh2 populations for the traits studied, which could be exploited for specific breeding objectives. The range of variability for most of the traits indicated that the field corn genotypes varied in their ability for sweet corn production. The higher estimates of genetic variances than the corresponding environmental variances for most of the traits indicated that the observed differences among the genotypes for most of the traits were more due to genetic than environmental causes. The conversion of the field corn genotypes to sh2 super-sweet corn offer great promise for the development of super-sweet corn adapted to the growing conditions of Nigeria. Opportunities abound for further improvement of the populations and extraction of lines for hybrid seed production.
ACKNOWLEDGEMENTS
The donor sh2 population used in this study was generously provided by Professor V. O. Adetimirin of the Department of Agronomy, University of Ibadan. The normal endosperm tropical field maize genotypes were sourced from the Maize Improvement Programme of the International Institute of Tropical Agriculture (IITA), Ibadan, Nigeria. This research did not benefit from any specific grant whatsoever.
REFERENCES
|
Adetimirin VO (2008). Stand establishment and early field vigour variation in a tropicalised shrunken-2 maize population. Field Crops Research 108:143-149. |
|
|
Alan O, Kinaci G, Kinaci E, Kutlu I, Budak Basciftci Z, Sonmez K, Evrenosoglu Y (2013). Genetic variability and association analysis of some quantitative characters in sweet corn. Notulae Botanicae Horti Agrobotanici 41(2):404-413. |
|
|
Bello OB, Ige SA, Azeez MA, Afolabi MS, Abdulmaliq SY, Mahamood J (2012). Heritability and genetic advance for grain yield and its component characters in maize (Zea mays L.). International Journal of Plant Research 2(5):138-145. |
|
|
Boyer CD, Shannon JC (1984). The use of endosperm genes for sweet corn improvement. Plant Breeding Reviews 1:139-161. |
|
|
Brewbaker JL (1977). 'Hawaiian superweet #9' corn. HortScience 12:355-356. |
|
|
Butrón A, Álvarez A, Revilla P, Malvar RA, Rodriguez VM, Galarreta JIR, Ordás A (2008). Agronomic performance of sweetcorn populations derived from crosses between sweetcorn and field corn. Journal of Agricultural Research 6:378-384 |
|
|
Cartea ME, Malvar RA, Revilla P, Ordás A (1996). Improvement of early vigor and adaptation of sweet corn to European Atlantic coast with open-pollinated field corn populations. Maydica 41:119-125. |
|
|
Creech RG (1965). Genetic control of carbohydrate synthesis in maize. Genetics 52:1175-1186. |
|
|
Deshmukh SNN, Basu MS, Reddy PS (1986). Genetic variability, character association and path coefficients of quantitative traits in Virginia bunch varieties of groundnut. Indian Journal of Agricultural Science 56:816-821. |
|
|
Dewanto V, Wu X, Liu RH (2002). Processed sweet corn has higher antioxidant activity. Journal of Agriculture and Food Chemistry 50(14):4959-4964. |
|
|
Entringer GC, Vettorazzi JCF, Crevelari JA, Durães NNL, Catarina RS, Pereira MG (2017). Super-sweet corn breeding by backcross: A new choice for the Brazilian market. Brazilian Journal of Agriculture 92(1):12-26. |
|
|
Food and Agriculture Organization (FAO) (2007). |
|
|
Food and Agriculture Organization (FAO) (2016). Food and Agriculture Organization of the United Nations statistical database, |
|
|
Gardner J, Hoffmann M, Smith ME (2000). Resistance to European corn borer in processing sweetcorn. HortScience 35:871-874. |
|
|
Hallauer AR (1990). Methods used in developing maize inbreds. Maydica 35:1-16. |
|
|
Hallauer AR, Carena MJ, Miranda Filho JB (2010). Quantitative ge¬netics in maize breeding. Springer Science+Business Media LCC New York, USA. doi 10.1007/978-1-4419-0766-0 |
|
|
Hefny M (2011). Genetic parameters and path analysis of yield and its components in corn inbred lines (Zea mays L.) at different sowing dates. Asian Journal of Crop Science 3(3):106-117. |
|
|
Jilo T, Tulu L, Birhan T, Beksisa L (2018). Genetic variability, heritability and genetic advance of maize (Zea mays L.) inbred lines for yield and yield related traits in southwestern Ethiopia. Journal of Plant Breeding and Crop Science 10(10):281-289. |
|
|
Johnson HW, Robinson HF, Comstock RE (1955). Estimation of genetic and environmental variability in soybeans. Agronomy Journal 47:314-318. |
|
|
Kim SK, Ajala SO (1996). Combining ability of tropical maize germplasm in West Africa. II. Tropical vs. Temperate Tropical origins. Maydica 41:135-141. |
|
|
Kim SK, Efron Y, Khadr F, Mareck J, Fajemisin J (1987). Registration of 16 maize streak virus resistant tropical maize parental inbred lines. Crop Science 27:824-825. |
|
|
Lertrat K, Pulam T (2007). Breeding for increased sweetness in sweet corn. International Journal of Plant Breeding 1(1):27-30. |
|
|
Lynch RE, Wiseman BR, Sumner HR, Plaisted D, Warnick D (1999). Management of corn earworm and fall armyworm (Lepdoptera: Noctuidae) injury on a sweet corn hybrid expressing a cryIA(b)gene. Journal of Economic Entomology. 92:1217-1222. |
|
|
Mahato A, Shahi JP, Singh PK, Kumar M (2018). Genetic diversity of sweet corn inbreds using agro-morphological traits and microsatellite markers. 3 Biotech 8:332 |
|
|
Malvar RA, Cartea ME, Revilla P, Butrón A, Ordás A (2001). Checking performance of field corn inbreds as donors of favourable alleles to improve early vigor and adaptation of sweet corn hybrids to European conditions. Maydica 46:187-193. |
|
|
Malvar RA, Revilla P, Cartea ME, Ordás A (1997). Field corn inbreds to improve sweet corn hybrids for early vigor and adaptation to European conditions. Maydica 42:247-255. |
|
|
Maphumulo SG, Derera J, Qwabe F, Fato P, Gasura E, Mafongoya P (2015). Heritability and genetic gain for grain yield and path coefficient analysis of some agronomic traits in early-maturing maize hybrids. Euphytica 206:225-244. |
|
|
Martin RV, Washington R, Downing TE (2000). Seasonal maize forecasting for South Africa and Zimbabwe derived from an agroclimatological model. Journal of Applied Meteorology 39:473-479. |
|
|
Niji MS, Ravikesavan R, Ganesan KN, Chitdeshwari T (2018). Genetic variability, heritability and character association studied in sweet corn (Zea mays L. saccharata). Electronic Journal of Plant Breeding 9(3):1038-1044. |
|
|
Nzuve F, Githiri S, Mukunya DM, Gethi J (2014). Genetic variability and correlation studies of grain yield and related agronomic traits in maize. Journal of Agricultural Science 6(9):166-176. |
|
|
Qi X, Zhao Y, Jiang L, Cui Y, Wang Y, Liu B (2009). QTL analysis of kernel soluble sugar content in super-sweet corn. African Journal of Biotechnology 8:6913-6917. |
|
|
Saleh G, Abdullah D, Anuar AR (2002). Performance, heterosis and heritability in selected tropical maize single, double and three-way cross hybrids. The Journal of Agricultural Science 138(1):21-28. |
|
|
Santos PHAD, Pereira MG, Trindade RS, Silva da Cunha K, Entringer GC, Vettorazzi JCF (2014). Agronomic performance of super-sweet corn genotypes in the north of Rio de Janeiro. Crop Breeding and Applied Biotechnology 14:8-14. |
|
|
SAS Institute Inc. (2003). The SAS system for Windows, release version 9.1.3. Cary, NC: SAS Institute. |
|
|
Sesay S, Ojo D, Ariyo OJ, Meseka S (2016). Genetic variability, heritability and genetic advance studies in top-cross and three-way cross maize (Zea mays L.) hybrids. Maydica 61(2). |
|
|
Singh BD (2001). Plant Breeding: Principles and Methods. Kalyani Publishers, New Delhi-Ludhiana, India 506 p. |
|
|
Singh RK, Chaudhary BD (2004). Biometrical methods in quantitative genetic analysis. Kalyani Publishers, New Delhi. |
|
|
Sleeper DA, Poehlman JM (2006). Breeding field crops, 5th edition. Blackwell Publishers Iowa, Ames. 424 p |
|
|
Souza ARR, Miranda GV, Pereira MG, de Souza LV (2009). Predicting the genetic gain in the Brazilian white maize landrace. Ciênc Rural 39:19-24. |
|
|
Teixeira FF, Miranda RA, Paes MCD, Souza SM, Gama EEG (2013). Melhoramento do milho doce. Sete Lagoas: Embrapa Milho e Sorgo. 32 p. |
|
|
Tracy WF (1997). History, genetics, and breeding of super-sweet (shrunken2) sweet corn. Plant Breeding Reviews14:189-236. |
|
|
Tracy WF (2001). Sweet corn. Speciality corns. Hallauer, A.R. Ed. 2nd ed. CRC Press, Boca Raton, Florida pp. 155-197. |
|
|
Tracy WF, Whitt SR, Buckler ES (2006). Recurrent mutation and genome evolution: example of Sugary1 and the origin of sweet maize. Crop Science 46:1-7. |
|
|
Yousef GG, Juvik JA (2002). Enhancement of seedling emergence in sweet corn by marker-assisted backcrossing of beneficial QTL. Crop Science 42(1):96-104. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


