Full Length Research Paper

ABSTRACT
F1 progenies derived from a set of diallel crosses involving native and 5 elite cucumber varieties were evaluated in a randomized complete block design (RCBD) with three replications to study combining ability for some important agronomic traits. The analysis of variance revealed that the mean squares due to genotypes were significant for the traits studied, indicating wide diversity among the parents. Mean squares for general combining ability (GCA) and specific combining ability (SCA) were significant, implying both additive and non-additive genetic system controlling the studied traits. Significant GCA values were recorded by the native variety (Odukpani) for vegetative traits, fruit girth and average fruit weight. Low GCA values were recorded for days to flowering by the elite varieties, implying they are negative combiners for early flower influx. GCA for number of pistillate flowers and number of fruits per plant were positive and significant for the elite varieties, indicating they could be exploited in cucumber breeding programmes for improved female flower production. The high mean values for vegetative traits and days to flowering of the progenies arising from crosses with Odukpani as the maternal parent appear to suggest cytoplasmic influence in the inheritance of the traits.
Key words: Combining ability, Cucumis sativus, cucumber, gene action.
INTRODUCTION
Cucumber (Cucumis sativus L.) is a widely cultivated plant and one of the most important and popular home garden vegetable species grown worldwide (Sun et al., 2006; Yang et al., 2014). The crop is noted for its high water content that confers it with diuretic properties. It also has a cleansing action as it removes accumulated pockets of old waste materials and chemical toxins from the body. Some cucumber varieties have relatively high levels of ß –carotene and evidence suggests that diets rich in ß –carotene and other carotenoids (such as lycopene) can prevent the onset of some chronic diseases such as anemia and prostate cancers (Cuevas et al., 2010).
Breeding for new and high yielding cultivars that are adapted to Nigeria’s ecologies is necessary because of the paucity of planting materials. This can be achieved through hybridization studies and exploitation of combining ability for important agronomic traits such as early flowering and fruit yield (Ene et al., 2018).
In many F1 hybrids breeding, analysis of combining ability has been used in practical plant improvement programs to determine the relative importance of general combining ability (GCA) and specific combining ability (SCA) with a view to harnessing superior parents for crossing in hybridization programs (Yoshioka et al., 2010; Adel and Ali, 2013, Golabadi et al., 2015). General combining ability is the manifestation of the additive gene action for the selection of parents and SCA represents the non-additive gene action (Singh et al., 2011). Superior traits identified in parents are not a guarantee that the traits can be transferred to the progenies (Ene et al., 2018); hence it is important to isolate potential combining lines through desirable cross combinations to obtain recombinants.
Improving fruit yield requires adequate information regarding the nature of GCA and SCA available in a wide array of genetic materials to be used in hybridization programmes. Several authors have used different genetic analyses to study combining ability in cucumber for fruit yield and yield components (Lopez-Sese and Staub, 2002; Xing Fang et al., 2004; Munshi et al., 2006; Mule et al., 2011; Sarkar and Sirohi, 2011; Singh et al., 2011; Olfati et al., 2012; Ene et al., 2018) and gynoecious sex expression (Wahid et al., 2003; Staub et al., 2005; Dey et al., 2010; Singh et al., 2011; Pati et al., 2015). Combining ability studies have been conducted by Adel and Ali (2013) Golabadi et al. (2015) and Ene et al. (2018), but these authors used elite parental lines to make crosses. Elite varieties of cucumber developed in advanced countries have been reported to have important qualitative and quantitative traits such as early flowering, improved gynoecious sex expression, fruit quality and yield traits (Sadiq et al., 2019; Chinatu et al., 2017; Pal et al., 2017). But farmers encounter some challenges in the use of the elite varieties as planting materials because of poor adaptation. They are vulnerable to both biotic and abiotic stresses which often culminate into poor fruit yield (Strefeler and Wehner, 1986).
Selective mating designs using adapted native and elite varieties as parents would allow for inter-mating of the selects in different cycles. It would also exploit both additive and non-additive gene effects, which could be useful for the genetic improvement of several yield traits (Singh and Pawar, 2005). Therefore, the objectives of the present study were to obtain information about the performance of cucumber genotypes and estimate the combining ability of the parents and their F1 progenies in a diallel cross between native and elite cucumber varieties.
MATERIALS AND METHODS
Experimental site
The studies were carried out in the early and late cropping seasons of 2016 at the Agricultural Services and Training Centre (ASTC), Vom-Jos, Plateau State, Nigeria. The area lies at latitude 9o55’N and longitude 8o35’E in the Southern Guinea Savanna ecology of Nigeria. The climate is characterized by two distinct wet and dry seasons. The wet season starts in April and ends in October while the dry season starts in November and ends in March. The mean annual rainfall is about 1,326mm and a relative humidity of between 22 and 87%. The mean monthly maximum and minimum temperatures are 34.5and 14.2°C, respectively; and an altitude of 1,195 m above sea level (Zitta and Madaki, 2020).
Genetic materials
The materials for the research comprised six parental lines made up of a popular local variety obtained from Odukpani in Cross River State which was obtained on the 25th February, 2015; and five elite varieties (Graffiton, Poinsett, Ashley, Marketmore and Monarch) obtained from Jos, Plateau State, Nigeria on the 10th March, 2015 (Table 1) and their F1 progenies. Field evaluations of the parents were earlier done at the Teaching and Research Farm of College of Agriculture, Garkawa between April- July, 2015 and August – November, 2015. The parents were crossed in a full diallel fashion between April and July (early cropping season) of 2016 to obtain the F1 seeds. To ensure reasonable emergence and proper protection of the seedlings, the F1 progenies and parents were planted in the nursery in a screen house at the ASTC, Vom, Jos. Both were transplanted 2 weeks after sowing (WAS) to the field in August (late cropping season), 2016 in a randomized complete block design (RCBD) with three replications. Planting was done using an intra- and inter- row spacings of 1m x 1m. The plot size was 2m x 4m (8m2) giving a plant population of 16 per plot. The total land area for the research work was 360m2. Weed control was done by manual weeding using small hoe. To achieve optimum fruit yield, NPK 15:15:15 fertilizer was applied at the rate of 120Kg/ha in two equal split doses at the time of seed sowing and before flowering as recommended by Iwalewa and Amujoyegbe (2019).
Five plants were randomly sampled for data collection on number of leaves/plant (NL/P), number of branches/plant (NB/P), vine length (VL), days to first flowering (DFF), days to 50% flowering (D50%F), number of staminate flowers plant-1(NSF/P), number of pistillate flowers/plant (NPF/P), days to first fruiting (DFFr), days to 50% fruiting (D50%Fr), average fruit weight (AFW) and number of fruits plant-1 (NFr/P).
Statistical analysis
Data from the parents (averaged over two cropping seasons) and F1 progenies were analyzed using Genstat 10.3 DE statistical package. Significant differences (p<0.05) among the genotypes were further performed using diallel analysis techniques (Hayman, 1954). The estimates of GCA and SCA effects of parents and hybrids were obtained as:
Where: Xij = the mean phenotypic value; µ = the general mean; GCA (gi and gj) = general combining ability effects of the ith and jth parents, respectively; SCA = (Sij) = specific combining ability effects of the cross i x j; Yij = the mean of i x j genotype; Yi = total array involving the jth parent as male in a diallel cross; Yii = the mean of the ith parent in the array; Yjj = the mean of the jth parent in the array; n = number of parents.
Levels of significance of the GCA and SCA were determined using the CD:
Error variances of the traits were determined as shown below:
SE value of GCA (gi - gj) = [2/(n + 2)]1/2;SE value of SCA
(sij) = [n(n-1)/(n=1)(n+2)]1/2
CD = critical difference; σ2e = error variance; t = t tabulated at 5% probability; n = number of parents; SE = standard error; GCA = general combining ability; SCA = specific combining ability; gi – gj = GCA of the ith and jth parents; sij = SCA of the cross i x j.
RESULTS
Mean comparison of agronomic traits for parents and F1 generations
The performances of the parents and crosses for the agronomic traits studied are presented in Table 2. The mean values for number of leaves per plant (NL/P), number of branches per plant (NB/P) and vine length (VL) for the parents ranged from 107.6 to 280.3, 9.5 to 23.5 and 101.3 to 296.9cm, respectively. The mean values of the F1 hybrids for NL/P, NB/P and VL ranged from 130.4-290.5, 9.4 to 25.8 and 117.8 to 301.6, in that order. When the indigenous accession, Odukpani, was used as the maternal parent the values for the vegetative traits of the hybrids were significantly higher and above grand mean (NL/P=200.3; NB/P=15.6; VL=200.8cm). The Odukpani parent was delayed for more than two weeks before attaining days to first flowering (DFF) and days to 50% flowering (D50F%). The performances of the F1 progenies for DFF and D50%F varied from 29.3 to 48.1 days and 32.7 to 54.7 days, respectively. The offspring arising from crosses with Odukpani as the maternal parent took longer days to flower.
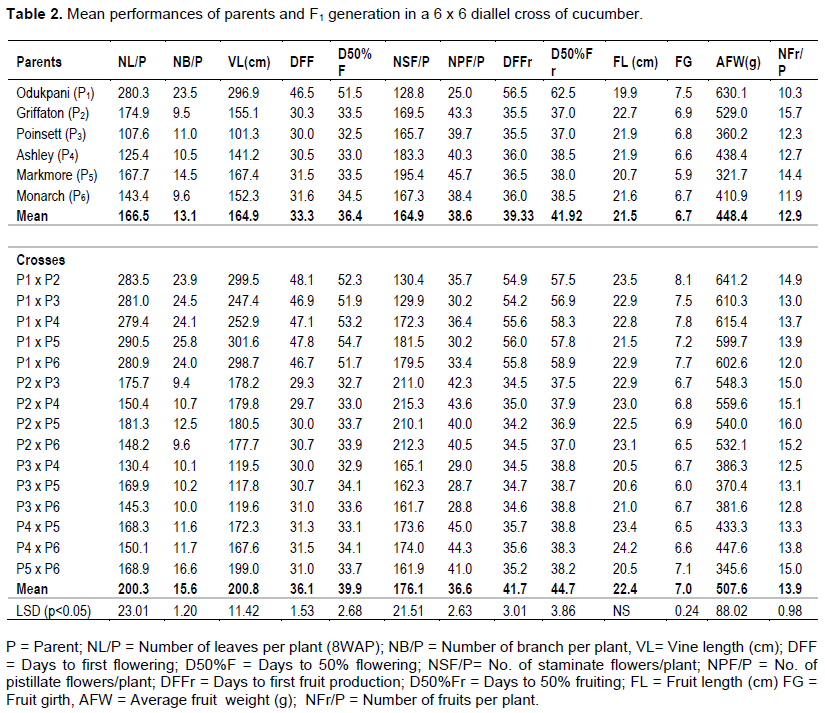
The mean number of staminate flowers plant-1 (NSF/P) varied from 82.8 (Odukpani) to 195.4 (Marketmore). The number of pistillate flowers ranged from 25.0 (Odukpani) to 45.7 (Marketmore). Seven F1 progenies (Griffaton x Poinsett, Griffaton x Ashley, Griffaton x Marketmore, Griffaton x Monarch, Ashley x Marketmore, Ashley x Monarch and Marketmore x Monarch) were prolific in the production of pistillate flowers (Table 2).
The mean number of days to first fruiting (DFFr) and days to fifty percent fruiting (D50%Fr) was significantly (p<0.05) delayed in the Odukpani parent (DFFr=56.5; D50%Fr=62.5ays). Consistent with days to flowering, days to fruiting took the same trend as offspring arising from crosses involving Odukpani as the maternal parents took longer days to fruit. The mean values for fruit girth (FG) differed significantly and varied from 7.5cm (Odukpani) to 5.9cm (Markmore). The highest mean values for FL (24.2 cm) and FG (8.1 cm) were recorded for Ashley x Monarch and Odukpani x Griffaton crosses, respectively. The average fruit weight and number of fruits plant-1 of the cucumber varieties evaluated varied from 321.7g (Markmore) to 630.1g (Odukpani) and 10.3 (Odukpani) to 15.7 (Griffaton), in that order. Nine Crosses (Odukpani x Griffaton, Odukpani x Poinsett, Odukpani x Ashley, Odukpani x Marketmore, Odukpani x Monarch, Griffaton x Poinsett, Griffaton x Ashley, Griffaton x Marketmore and Griffaton x Monarch) were outstanding; while seven progenies (Odukpani x Griffaton, Odukpani x Marketmore, Griffaton x Poinsett, Griffaton x Ashley, Griffaton x Marketmore, Griffaton x Monarch and Marketmore x Monarch) were promising with respect to number of fruits per plant (Table 2).
Analysis of variance for agronomic traits
Vegetative traits
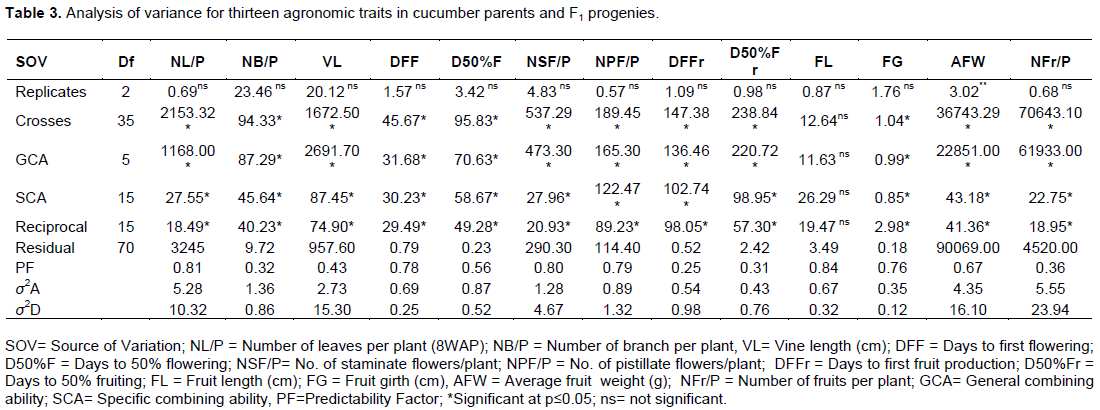
The analysis of variance results revealed that general combining ability (GCA) and specific combining ability (SCA) effects were significant for mean number of leaves plant-1 (NL/P), number of primary branches plant-1 (NB/P) and vine length (VL). Higher values for dominant effects (σ2D) were obtained for NL/P and VL; while additive effect (σ2A) was high for NB/P (Table 3).
Flowering traits
The GCA and SCA effects were significant for mean days to first flowering (DFF), days to fifty percent flowering (D50%F), number of staminate flowers plant-1 (NSF/P) and number of pistillate flowers plant-1 (NPF/P). Higher values of σ2D were recorded for NSF/P and NPF/P. Conversely, the σ2A value was found to be superior to the σ2D for DFF and D50%F (Table 3).
Fruit size and yield
While GCA and SCA for fruit length were not significant (p<0.05); they were significant for fruit girth. The variance component analysis and predictability factor showed that additive component had greater effects on the genetic control of fruit girth. Significant effects of GCA and SCA were also recorded for fruit weight and number of fruits per plant. Higher magnitude of the dominant effects, compared with the additive effects were obtained for these traits (Table 3).
GCA and SCA effects for parents and F1 progenies
Estimates of GCA effects for the parents varied from -3.31 (Ashley) to 4.27 (Odukpani) for number of leaves per plant (NL/P), -0.22 (Monarch) to 4.70 (Odukpani) for number of branches per plant (NB/P) and -0.04 (Poinsett) to 5.01 (Odukpani) for vine length (VL). The native variety (Odukpani) recorded positive and significantly high GCA value compared to the values recorded for the elite varieties for the vegetative traits assessed (Table 4). Specific combining ability for the progenies identified seven hybrids (Odukpani x Griffaton, Odukpani x Marketmore, Odukpani x Monarch, Griffaton x Poinsett, Griffaton x Marketmore, Griffaton x Monarch and Marketmore x Monarch ) and eight (Odukpani x Griffaton, Odukpani x Marketmore, Odukpani x Monarch, Griffaton x Poinsett, Griffaton x Marketmore, Poinsett x Ashley, Poinsett x Marketmore and Marketmore x Monarch) crosses as positive combiners for number of leaves per plant and number of branches per plant, respectively. Similarly, ten hybrids (Odukpani x Griffaton, Odukpani x Ashley, Odukpani x Marketmore, Odukpani x Monarch, Griffaton x Poinsett, Griffaton x Ashley, Griffaton x Marketmore, Poinsett x Ashley, Poinsett x Marketmore and Marketmore x Monarch) were positive combiners for vine length (Table 4). The Odukpani variety also recorded positive and significant GCA values for days to first flowering (DFF) (4.12*) and days to 50% flowering (D50%F) (5.32*). The values for DFF and D50%F for three elite varieties, Griffaton, Marketmore and Monarch were significant but negative (-4.78* to -0.67*). General combining ability effects for number of staminate flowers per plant (NSF/P) and number of pistillate flowers per plant (NPF/P) ranged from -1.89* to 4.23* and -1.65* to 4.49*, respectively. The Odukpani accession was a negative combiner for the flowering traits assessed. Significant SCA effects for the progenies ranged from -10.21* to 0.50* (DFF), -2.39* to 3.33* (D50%F), -6.23* to 4.77* (NSF/P) and -1.03* to 5.67* (NPF/P). Ten F1 progenies recorded positive and significant SCA values for production of pistillate flowers. The progenies comprised Odukpani x Griffaton, Odukpani x Ashley, Odukpani x Marketmore, Griffaton x Poinsett, Griffaton x Ashley, Griffaton x Marketmore, Poinsett x Ashley, Poinsett x Marketmore, Ashley x Marketmore and Ashley x Monarch. Estimates of GCA for days to first fruiting (DFFr) and days to 50% fruiting (D50%Fr) varied from -3.46* (Graffiton) to 4.24* (Odukpani) and from -3.94* (Griffaton) to 4.37* (Odukpani), respectively. The crosses with negative and significant SCA effects for both days to first and 50% fruiting included Odukpani x Griffaton, Odukpani x Marketmore, Griffaton x Poinsett, Griffaton x Ashley, Griffaton x Marketmore, Griffaton x Monarch, Poinsett x Marketmore, Poinsett x Monarch and Marketmore x Monarch (Table 4).
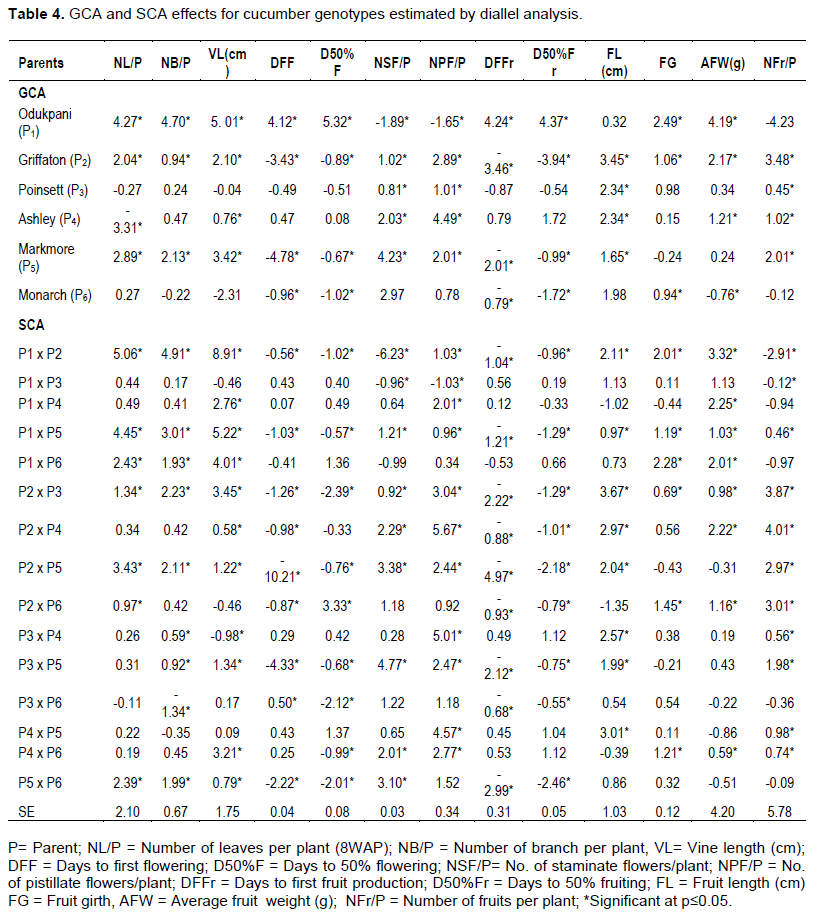
The Odukpani accession was observed to be a positive combiner for fruit girth (FG) as it recorded significantly higher GCA (2.49*) than the elite varieties. SCA effects for fruit length were significant and positive for nine crosses (Odukpani x Griffaton, Odukpani x Marketmore, Griffaton x Poinsett, Griffaton x Ashley, Griffaton x Marketmore, Griffaton x Monarch, Poinsett x Ashley, Poinsett x Marketmore and Ashley x Marketmore) and also significantly positive for fruit girth for six crosses (Odukpani x Griffaton, Odukpani x Marketmore, Odukpani x Monarch, Griffaton x Poinsett, Griffaton x Monarch and Ashley x Monarch). For the parental lines, GCA effects for average fruit weight (AFW) varied from -0.76* to 4.19* and for mean number of fruits per plant (NFr/P) it ranged from -4.23 to 3.38. Significant and positive SCA effects were observed for eight (Odukpani x Griffaton, Odukpani x Ashley, Odukpani x Marketmore, Odukpani x Monarch, Griffaton x Poinsett, Griffaton x Ashley, Griffaton x Monarch and Ashley x Monarch) and nine (Odukpani x Marketmore, Griffaton x Poinsett, Griffaton x Ashley, Griffaton x Marketmore, Griffaton x Monarch, Poinsett x Ashley, Poinsett x Marketmore, Ashley x Marketmore and Ashley x Monarch) crosses for AFW and NFr/P, respectively (Table 4).
DISCUSSION
The significant differences in vegetative, flowering and fruit traits implied that the traits had significant contributions towards diversity among the genotypes evaluated. The greater diversity in the present materials is due to the yield characters which will offer a promising base for improvement of the fruit yield through selection from the parents and progenies. The superior mean values for the agronomic traits of the progenies arising from crosses with Odukpani as the maternal parent suggests some form of cytoplasmic effects in the inheritance of these traits. Maternal effects have been implicated in the inheritance of cucumber leaf and fruits traits (Golabadi et al., 2015) and in days to flowering in cowpea (Manggoel and Uguru, 2012). According to Etterson and Galloway (2002) responses to selection may be affected by maternal effect because for many characters, the genotype of the mother via the cytoplasm account for a considerable portion of the genetically based variation in the progeny phenotype. Therefore, selection based on direct genetic effect may not be adequate, as it may lead to omission of potentially important source of genetic variance contributed by the cytoplasm of the maternal strain (Wolf, 2002).
The superior mean values for female flower production by the crosses: Graffiton x Poinsett, Graffiton x Ashley, Graffiton x Markmore, Graffiton x Monarch, Ashley x Markmore, Ashley x Monarch and Markmore x Monarch, suggest high reproductive efficiency. Therefore, the progenies that recorded high number of pistillate flowers could be selected as candidates in breeding programs whose main interest is in increased number of gynoecious flowers. Similarly, progenies from crosses between Odukpani and Griffaton were identified suitable combination for improving fruit weight and number of fruits as the two parents produced outstanding progenies for these traits.
The result of the variance components and predictability factor, in this study, indicated the superiority of dominant gene effects over additive gene effects for the genetic control of number of leaves per plant and vine length. Higher values for additive gene effects obtained for number of leaves per plant implied the superiority of additive over dominant gene control for this trait. Similar reports by Dogra and Kanwar (2011) showed the superiority of dominant gene effects over additive effects in cucumbers. The findings from this study is however inconsistent with that reported by Golabadi et al. (2015).
The authors reported relative superiority of additive gene effects over dominant gene effects for the genetic control of vegetative traits in cucumber. The discrepancy might be due to differences in the parents used and the environmental conditions of the two studies. The higher value of σ2D and the high deviation of the predictability factor from unity implied that selection methods would be preferred for enhancement in the genetic control of number of leaves and vine length. On the other hand, the superior additive effects of number of branches implied the predominance of gene action in the genetic control of the trait. Since selection for vegetative traits would directly influence photosynthesis, information on the genetic control of these traits would be important in enhancing optimum fruit production in cucumber (Golabadi et al., 2015).
Flowering is an important physiological attribute for crop survival and continuity. Flowering is of great importance in annual crops, including cucumber, as it is a component of the adaptation of a variety to a particular environment and it also determines yield (Manggoel and Uguru, 2012; Ishiyaku et al., 2005). The significant GCA and SCA for the flowering traits assessed in this study imply the importance of both additive and dominant genetic controls of these traits in cucumber. The higher value of σ2D compared with that of σ2A for number of staminate flowers per plant and number of pistillate flowers per plant indicate the predominance of gene action in the genetic control of these traits. The superior σ2A value for days to first and 50% flowering implied the predominance of additive effects. This indicates that selection methods would be preferred for the enhancement of these traits. If the genetic basis of early and continuous flowering is understood, it can be exploited in the development of cucumber varieties that can flower and fruit continually thereby ensuring all year round availability of cucumbers to the teaming population in sub Saharan Africa.
Fruit length (FL) and fruit girth (FG) are the parameters that made up fruit size. Fruit size is an important determinant of yield, quality and marketability. The significant GCA and SCA effects for FG showed that both additive and dominant gene actions have significant effects on the genetic control of this trait. This report is consistent to reports of significant GCA and SCA effects for FG by earlier authors (Singh et al., 2011; Mule et al.; 2011; Golabadi et al. 2015). This result is also in agreement with an earlier report by Hormuzdi and More (1989), but inconsistent with those reported by Dogra and Kanwar (2011) and Sarkar and Sirohi (2011). The discrepancy might be due to differences in the parents used and the environmental conditions of the two studies for making diallel crosses.
Fruit yield has been reported as the most important breeding trait in cucumber (Singh et al., 2011; Olfati et al. 2012). Average fruit weight (AFW) and number of fruit plant-1 (NFr/P) are important yield component traits that affects cucumber yield. The significant GCA and SCA effects for AFW and NFr/P obtained in this study is in agreement with the findings of Golabadi et al. (2015) and Jagesh et al. (2013). The higher magnitude of σ2D, compared with σ2A, indicate the dominant genetic control of fruit yield. The predominance of additive gene effects in the genetic control of fruit yield in cucumber has been reported (Olfati et al., 2012). The relatively small deviation of predictability factor from unity observed in this study indicates the importance of additive gene action for the genetic control of number of fruits per plant suggesting that cyclic selection should be effective for improving number of fruits per plant. Lopez-Sese and Staub (2002) reported that fruit number was under the genetic control of both additive and dominant gene actions. Sarkar and Sirohi (2011) also reported the over-dominant gene action for the genetic control of fruit number per plant and suggested heterosis breeding for achieving higher yields in cucumber.
Combining ability is a useful prediction of genetic capability of parental lines and crosses. Greater GCA values for traits indicate the capacity of the parent to produce superior progenies for the traits. The high GCA value for vegetative traits recorded by the indigenous accession (Odukpani) was considered to be an appropriate parent for hybridization as a positive combiner for these traits.
This indigenous genetic material could, therefore, be selected in hybridization programs for improving the vegetative traits that would, in turn, enhance photosynthetic capacity. Prolific vegetative growth is an important variable for most physiological and agronomic studies as it helps to enhance photosynthetic efficiency and yield (Blanco and Folegatti, 2005).
Earliness to flowering and fruiting are important agronomic traits that can be exploited to ensure continuous and all year round production. The elite varieties with low GCA values for days to flowering were considered to be appropriate parents for hybridization as negative combiners to reduce the length of days to flowering. In earliness to flower appearance, it showed that the genotypes are good combiners for early flower influx. The significantly higher GCA values recorded for NSF/P and NPF/P by the elite varieties implied that these parents could be selected in hybridization programs for improving flower production and reproductive efficiency.
Fruit length (FL) and fruit girth (FG) are marketable characters that determine fruit size. Similarly, average fruit weight (AFW) and number of fruits per plant (NFr/P) are indicators for yield improvement. The high value of GCA effects recorded by Odukpani (FG and AFW) and Griffaton (FL and NFr/P) implied they are good combiners and they are candidates that could be selected in hybridization programs for improving these traits. Mule et al. (2011) reported the best general combiners for fruit length and fruit diameter in a line tester analysis.
CONCLUSION
The native Odukpani varieties with high GCA value for vegetative traits, fruit girth and fruit weight is a promising parent for hybridization as the best combiner for these traits. The low GCA values for days to flowering recorded by the elite varieties implied they are negative combiners for early flowering. The significantly positive GCA for number of pistillate flowers and number of fruits plant-1 observed for the elite varieties indicated they are best combiners and could be exploited in cucumber breeding programmes for superior female flower production and fruit yield. The superior mean values for number of pistillate flowers by the crosses Graffiton x Poinsett, Graffiton x Ashley, Graffiton x Markmore, Graffiton x Monarch, Ashley x Markmore, Ashley x Monarch and Markmore x Monarch implied high reproductive efficiency. Therefore, these progenies could be selected as candidates in breeding programs aimed at increased number of gynoecious flowers. The significantly high mean values for days to flowering and some yield traits in progenies arising from crosses with Odukpani as the maternal parent suggests cytoplasmic influence in the inheritance of these traits.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interest.
REFERENCES
|
Adel MM, Ali EA (2013). Gene Action and Combining Ability in a Six Parent Diallel Cross of Wheat. Asian Journal of Crop Science 5(1):14-23. |
|
|
Blanco FF, Folegatti MV (2005). Estimation of leaf area for greenhouse cucumber by linear measurements under salinity and grafting. Scientia agricola (Piracicaba, Braz.) 62(4):305-309. |
|
|
Chinatu LN, Onwuchekwa-Henry CB, Okoronkwo CM (2017). Assessment of Yield and Yield Components of Cucumber (Cucumis sativus L.) in Southeastern Nigeria. International Journal of Agriculture and Earth Science 3(1):35-44. |
|
|
Cuevas HE, Song H, Staub JE, Simon PW (2010). Inheritance of Beta-Carotene-Associated Flesh Color in Cucumber (Cucumis Sativus L.) Fruit. Euphytica 171(3):301-311. |
|
|
Dey S, Behera T, Munshi A, Pal A (2010). Gynoecious inbred with better combining ability improves yield and earliness in bitter gourd (Momordica charantia L.). Euphytica 173:37-47. |
|
|
Dogra BS, Kanwar MS (2011). Exploitation of combining ability in cucumber (Cucumis sativus L.). Agricultural Science Research Journal 2:55-59. |
|
|
Ene CO, Ogbonna PE, Agbo CU, Chukwudi,UP (2018). Heterosis and combining ability in cucumber (Cucumis sativus L.). Information Processing in Agriculture. https:// doi.org/10.1016/j.inpa.2018.07.008 |
|
|
Etterson D, Galloway LF (2002). The influence of light on paternal plants in Campanula americana (Campanulaceae): Pollen characteristics and offspring traits. American Journal of Botany 89(12):1899-1906. |
|
|
Golabadi M, Golkar P, Eghtedary A (2015). Combining ability analysis |
|
|
of fruit yield and morphological traits in greenhouse cucumber (Cucumis sativus L.). Canadian Journal of Plant Science 95(2):377-385. |
|
|
Hayman BI (1954). The Theory and analysis of diallel crosses. Genetics 39:789-809. |
|
|
Hormuzdi SG, More TA (1989). Studies on combining ability in cucumber (Cucumis sativus L.). Indian Journal of Genetics and Plant Breeding 49(2):161-165. |
|
|
Ishiyaku MF, Singh BB, Craufurd PQ (2005). Inheritance of time to flowering in Cowpea [Vigna unguiculata (L.) Walp.]. Euphytica 142(3):291-300. |
|
|
Iwalewa EA, Amujoyegbe BJ (2019). Influence of fertilizer types and rates on the growth and fruit yield of three cucumber (Cucumis sativus L.) varieties. Ife Journal of Agriculture 31(3):36-49. |
|
|
Jagesh K, Munshi AD, Kumar R., Sureja AK, Sharma RK (2013). Combining ability and its relationship with gene action in slicing cucumber. Indian Journal of Horticulture 70(1):135-138. |
|
|
Lopez-Sese AI, Staub J (2002). Combining ability analysis of yield components in cucumber. Journal of the American Society for Horticultural Science 127(6):931-937. |
|
|
Manggoel W, Uguru, MI (2012). Evidence of Maternal Effect on the Inheritance of Flowering Time in Cowpea (Vigna unguiculata (L.) Walp.) International Journal of Plant Breeding and Genetics 6(1):1-16. |
|
|
Mule PN, Khandelwel V, Patil AB, Chaudhary BR (2011). Combining ability studies in cucumber (Cucumis sativus L.). Vegetable. Science 38:203-205. |
|
|
Munshi AD, Kumar R, Panda B (2006). Combining ability in cucumber (Cucumis sativus L.). Indian Journal of. Agricultural. Science 76:750-752. |
|
|
Olfati JA, Samizadeh H, Rabiei B, Peyvast GH (2012). Grif?ng's methods comparison for general and speci?c combining ability in cucumber. Scientific World Journal 2012:1-4. |
|
|
Pal S, Rahj H, Yadav SN (2017). Evaluation of cucumber genotypes for yield and quality traits. Journal of Hill Agriculture 8(2):144-150. |
|
|
Pati K, DasMunshi A, Behera TK (2015). Inheritance of gynoecism in cucumber (Cucumis sativus L.) using genotype gbs-1 as gynoecious parent. Genetika 47(1):349-356. |
|
|
Sadiq GA, Omerkhil N, Zada KA, Safdary AJ (2019). Evaluation of growth and yield performance of five cucumbers (Cucumis sativus L.) genotypes: Case study Kunduz province, Afghanistan. International Journal of Advanced Education and Research 4(6):22-28 |
|
|
Sarkar M, Sirohi PS (2011). Diallel analysis of quantitative characters in cucumber (Cucumis sativus L.). Vegetable Science 38(1):73-75. |
|
|
Singh S, Pawar IS (2005). Theory and application of biometrical genetics. CBS Press, Frederiksberg, Denmark. |
|
|
Singh R, Singh AK, Kumar S, Singh BK, Singh SP (2011). Studies on combining ability in Cucumber (Cucumis Sativus L.). Vegetable Science 38(1):49-52. |
|
|
Staub JE, Fan Z, Sun Z, Lower RL (2005). Identification and use of yield and fruit quality genes in cucumber. Pickle Packers International Meeting Proceedings P 3. |
|
|
Strefeler MS, Wehner TC (1986). Estimates of Heritabilities and Genetic Variances of Three Yield and Five Quality Traits in Three Fresh-market Cucumber Populations. Journal of the American Society for Horticultural Science 111(4): 599-605. |
|
|
Sun Z, Lower RL, Staub JE (2006). Variance component analysis of parthenocarpy in elite U.S. processing type cucumber (Cucumis sativus L.) lines. Euphytica 148(3):333-341. |
|
|
Wahid MM, Medany MA, Abou-Hadid AF (2003). Diallel analysis for yield and vegetative characteristics in cucumber (Cucumis sativus L.) under low temperature conditions. ISHS Acta Horticulturae, p. 598. |
|
|
Wolf JB, Vaughn TT, Pletscther LS, Cheverud JM (2002).Contribution of Maternal effect QTL to genetic architecture of early growth in mice. Heredity 89(4):300-310. |
|
|
Xing Fang G, Ping ZS, Qing XC (2004). Analysis of combining ability of early yield and total yield character of cucumber cultivated in open ?eld in spring. China Vegetable 6:13-15. |
|
|
Yang L, Koo D, Li D, Jiang J, Luan F, Renner SS, Henaff E, Sanseverino W, Garcia-Mas J, Casacuberta J, Senalik DA, Simon PW, Chen J, Weng Y (2014). Mechanisms of decreasing dysploidy in Cucumis [abstract]. Plant and Animal Genome Conference. Paper No. W233. |
|
|
Yoshioka Y, Sugiyama M, Sakata Y (2010). Combining ability analysis of fruit texture in cucumber by mechanical measurement. Breeding Science 60(1):65-70. |
|
|
Zitta WS, Madaki DH (2020). Climate Change, Rainfall Trends and variability in Jos Plateau. Journal of Applied Science 20(2):76-82. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


