Full Length Research Paper

ABSTRACT
Lablab purpureus is a multipurpose legume mainly grown by subsistence farmers for pulse, forage and vegetable. Despite these diverse uses that can be combined successfully under various agronomic conditions, the bean has not been extensively exploited. Dolichos production in Kenya is constrained by low yielding varieties, pests, poor growing techniques and varieties with non-preferred taste and flavor. A study to characterize six newly bred Lablab genotypes (G2, B1, M5, LG1, W7 and G2), was initiated at the University of Eldoret using morphological and molecular markers. Morphological traits (qualitative and quantitative) were characterized using the descriptors of genus Lablab. Molecular characterization was done at the KEPHIS- Muguga laboratory, using ten SSR markers that are specific for Dolichos. Results from morphological characterization demonstrated a high variation for both qualitative and quantitative traits evaluated. Most of the quantitative traits were highly significant at 99.999% or (P≤ 0.001), except for number of racemes per plant and number of seeds per pod which were significant at 99.99% or (P≤ 0.01) and 99.95% or (P≤ 0.05), respectively. Microsatellite analysis produced six polymorphic primers mapping an average of 3.17 alleles per locus. The combination of morphological and DNA-based markers takes advantage of the best features of both marker types which can be beneficial in selection of best Lablab genotypes and in the process integrates the application of molecular markers to legume breeding.
Key words: Lablab (Lablab purpureus), microsatellites (SSR), genetic diversity.
INTRODUCTION
Dolichos (Lablab purpureus (L.) Sweet) is a grain legume, indigenous to Asia and Africa (Pengelly and Maass, 2001). It is scientifically classified in the family: Fabaceae, the tribe: Phaseolae, Genus: Dolichos and Species: L. purpureus (Rai, 2010). Lablab is a legume well suited to most tropical environments as it is adaptable to a wide range of rainfall, temperature and altitude (Ravinaik et al., 2015).
It is a high grain – yielding, nitrogen fixing, dual purpose legume that improves soil fertility and some more genetically diverse varieties can survive a dry spell. It is more drought tolerant, than common beans (Phaseolus vulgaris), soybean (Glycine max) or cowpea (Vigna unguiculata) and has grain yields that are higher than cowpeas (Adebisi and Bosch, 2004). Therefore, Lablab bean can virtually substitute these common legumes whose growth in drier areas is limited by high temperatures and low rainfall. It can grow in areas where rainfall is less than 500 mm, but loses leaves during prolonged drought periods (Girishi and Gowda, 2009). This drought tolerance feature of L. purpureus is enhanced by its capability to extract soil water from at least 2 m depth even in fine textured soil (Khourya et al., 2014; Burkill, 1995). Local subsistence farmers in Africa traditionally grow it for human consumption, vegetable (flowers and immature pods), green manure, cover crop and concentrate feed (Maass et al., 2010). In 100 g of green pods of Lablab, it contains approximately 6.7 g carbohydrates, 3.8 g proteins, 1.8 g fibre, 210 mg calcium, 68.0 mg phosphorus, and 1.7 mg iron (Kumar et al., 2018). Due to its potential for use as vegetable, pulse, weed control and soil improvement, Lablab has been found to be of great significance (Murphy and Colucci, 2009; Mureithi et al., 2003). Despite Lablab bean’ importance in the food and farming systems, particularly among communities in arid and semi-arid lands, it has remained neglected and underutilized (National Research Council, 2006) and indeed Maass et al. (2010) classified it among the lost crops of Africa.
In Kenya, the grain yield of Lablab on farmers’ fields is low (range between 800 to 900 kg/ha) compared to the yield potential of 2700 to 3000 kg/ha (Kamau et al., 2010; Kamotho et al., 2010). The low yield is attributed to use of unimproved varieties, influx of pests, such as mites and aphids and low use of fertilizers (Kinyua et al., 2008). Despite the several uses of Lablab, its potential in improving agricultural systems of farmers and nutritional status of many households has not been fully exploited. Lablab production in the country is constrained by low yielding varieties, pests, poor growing techniques and varieties with non-preferred taste and flavor, lack of certified seeds and limited information on production practices (Kinyua et al., 2008). Genetic improvement on yield and disease resistance has been successfully applied by researchers working on L. purpureus varieties from Indian ecogeographical zones, who have used genetic markers (Rai et al., 2010; Innes, 2004). The crop improvement program in Kenya aims at employing these improved techniques. The estimation of genetic diversity of a crop species is the basis for its improvement. Although morphological markers enable the detection of genetic variation, they are often influenced by a myriad of external factors in the environment and minimized by a paucity of discernible morphological markers (Kamotho et al., 2016). Significant advancements in molecular biology have shifted the focus of assessment of biodiversity from relying only on morphological markers to using isozymes and DNA markers (Konstantinos, 2008). The use of DNA -based markers in recent years has been used as an important tool for assessing genetic diversity of crops (Gomez et al., 2004), however, morpho-agronomic traits still play a significant role and thus a combined approach is recommended. Therefore, there is a need to incooperate both morphological and molecular tools in characterization of crop species. At the University of Eldoret Biotechnology Department, breeding programs have been developed to breed for the improvement of organoleptic traits (cooking time and taste) and to develop high yielding Lablab varieties. Therefore, the objective of this study was to evaluate the diversity of six improved Dolichos genotypes bred at University of Eldoret based on a combined morpho-agronomic traits and SSR markers.
MATERIALS AND METHODS
Genetic material
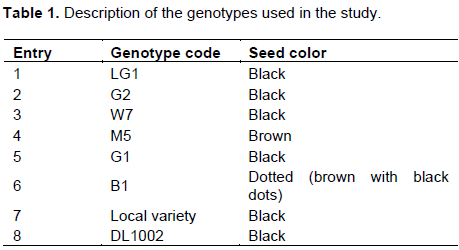
The genotypes used in the current study comprised 6 lines that had been bred at University of Eldoret (W7, M5, B1, G1, G2 and LG1) to improve their yield, cooking time, taste and two commercial checks (DL1002 and a local land race (Local variety) collected from farmers’ field in Meru county, Ruiri village). The genotypes were selected based on adaptability, yield and ability to withstand pests and other diseases. The genotypes are as described in Table 1.
Experimental design, locations and data collection
Morphological characterization
Morpho-agronomic trait evaluation of the genotypes was carried out in three locations representing different agro-climatic zones with different climatic and soil conditions as follows: University of Eldoret farm in Uasin-Gishu county; (0° 34’N, 35° 17’E), at 2134 m above sea level with rhodic ferrosols soil type, with a mean annual temperature of 16.6°C and a mean annual rainfall of 1124 mm), Abma farm in Njoro Nakuru county (0° 20’N, 35° 56’E), at 1920 m above sea level with nitisols soil type, a mean annual temperature of 22°C and a mean annual rainfall of 800 mm) and Mabanga farmer training college in Bungoma county (0° 35’N, 34° 37’E), at 1440 m above sea level with acrisols and nitisols soil type, a mean annual temperature of 26°C and a mean annual rainfall of 700 mm). The experimental design for the trials was Randomized Complete Block Design (RCBD) with three replications. Uniformity in management was ensured particularly within the blocks. Each experimental plot measured 32 m by 13.25 m. Management of each site was as per the recommended standards for each study region. During morphological characterization, qualitative and quantitative traits mainly yield related components, were measured at different growth stages on all the plants. The variables were scored using Gowda (2008) protocol, descriptor list for the genus Lablab. All agronomic practices were carried out including weeding.
The qualitative traits that were measured included: main stem pigmentation, leaf anthocyanin, leaf color, leaf hairiness on inner surface, leaf shape, growth habit, branch orientation, flower color, pod curvature, pod beak, pod color of mature pods, unripe pod attachment, seed color at maturity, and seed shape when dry.
The quantitative traits included: days to 50% flowering (days from emergence to 50% of the plants producing flowers), days to maturity (from emergence to stage when 50% of the plants have mature pods), flower buds/raceme (average number of buds from 10 randomly chosen racemes), racemes/plant (average number of racemes from 10 randomly chosen plants), pods/plant (average number of pods from 10 randomly chosen plants), seeds/pod (average number of seeds from 10 randomly chosen ripe pods), plant height (cm) (measured on 10 random matured plants from the cotyledon scar to tip of the plant), and 1000 seed weight (g) (average weight of 1000 seeds). Data was taken from four inner rows since one outer row from both sides of each plot was removed to take care of border effect. Values used for analysis were measured from 10 plants chosen at random in each plot.
Genetic diversity of the Dolichos genotypes using SSR markers
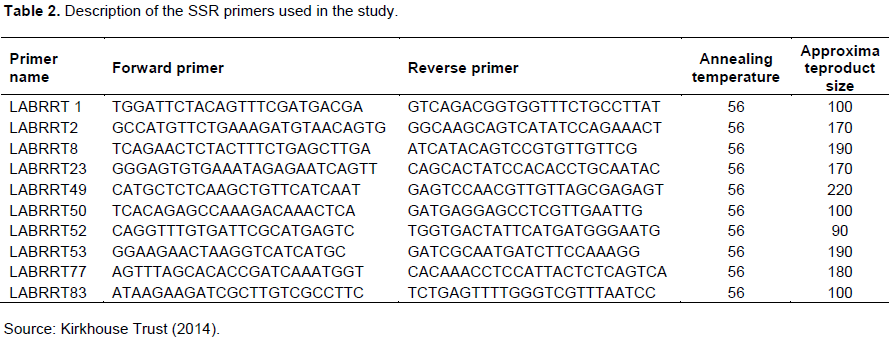
Polymerase chain reaction on the isolated genomic DNA obtained was carried out using ten SSR primer pairs selected from a list of 25 primers developed by Kirk House Trust (Table 2).
Ten plants of each of the six genotypes under study were planted in a greenhouse at the Kenya Plant Health Inspectorate Service (KEPHIS), Muguga laboratories. Young fresh leaves of each genotype were harvested from three-week-old plants and ground into fine powder under liquid nitrogen using a motor and pestle for DNA extraction. The cetyl-tri-methyl-ammonium bromide (CTAB) method (Kimani et al., 2012) was carried out with slight modifications as follows: leaf extracts were transferred to labeled Eppendorf tubes each containing 500 µl of 2 × CTAB mecaptoethanol extraction buffer and the samples placed on ice. The contents were subsequently incubated in a water bath at 65°C for 1 h with invasions after every 15 min. This was followed by addition of 500 µl of chloroform-isoamyl alcohol (24:1) and inversion for 5 min at room temperature to mix. After a centrifuge at 14000 rpm for 10 min, 400 µl of the top clear layer was pipetted into fresh Eppendorf tubes and 250 µl of isopropanol added then incubated at room temperature for 10 min.
A further centrifuge of the mixture at 14000 rpm for 10 min was done to pellet the DNA, the supernatant discarded and 320 µl of 1 × TE was added and the samples placed on ice. A further 40 µl of Magnesium chloride was added and the contents incubated on ice for 10 min followed by a centrifugation to14000 rpm for 10 min and the supernatant discarded. The pellet was then vacuum dried for 5 min before adding 5 µl of R-nae enzyme and placed in a water bath (37°C) for 2 h. 40 µl of sodium acetate was added followed by 250 µl of isopropanol and the contents incubated for 15 min at room temperature.
A 1 ml aliquot of 70% ethanol was then added to the DNA pellet followed by another centrifuge of 14000 rpm for 5 min. The pellet was then vacuum dried for 3 min to remove any liquid and the DNA pellet re-suspended in 50 µl of 1 × TE. It was then left to stand for 10 min at room temperature before storing at 4°C.
The quality and quantity of the DNA was verified by electrophoresis on a 0.8% (w/v) agarose gel, for 30 min at 80 V. Lambda (λ) phage DNA was used as the standard. After electrophoresis, the gel was stained in ethidium bromide (10 mg/ml) for 30 min and later de-stained in distilled water for 20 min before viewing under ultraviolet trans illuminator. The concentrations of the samples were determined by comparing band sizes and intensities of the test
Subsets of 10 SSR primer pairs specific to Dolichos were selected from 25 primers developed by Kirk House foundation. The PCR reactions were performed in a Master cycler (Eppendorf®) using in a final volume of 20 μl Bioneer AccuPower® containing 4 μl pre-mix (1U Top DNA, 250 μM each dNTP, 10 mM Tris-HCl pH 9.0, 30 mM KCl, 1.5 mM MgCl, stabilizer and tracking dye), 0.0025 ng/μl of each forward and reverse primer, 0.5 ng of template DNA, and 6 μl of double distilled water (ddH2O). The PCR cycles consisted of: 92°C for 3 min for initial denaturation, 92°C for 3 min for actual denaturation, annealing at 56°C for 30 s, extension at 72°C for 1 min, followed by 34 cycles of 30 s at 92°C, 1 min at 56°C, 1 min at 72°C, and a final extension step of 5 min at 72°C (Supplementary Table 1).
The DNA fragments were separated on 4% agarose gel run at 100 V for 2 h using 0.5M TBE buffer. The DNA fragments in gel was visualized by staining in 0.5 µg/mg ethidium bromide for 30 min and rinsed in distilled water for 20 min, visualized and photographed on ultraviolet (UV) trans-illuminator at 312 nm. Allele sizes were scored using a 1000 base pair (bp) molecular size ladder.
Statistical analysis
A table of means from quantitative data obtained was subjected to genstat 13th edition and the means of the genotypes separated using least significant difference (LSD) at 5% level of probability of the same software. This was to check on any difference among genotypes. Coefficient of variation (CV) was used to measure statistically the dispersion of data around the mean.
Cluster analysis
The morphological data were scored as binary data and were used to construct a dendogram. A dendogram was constructed for the quantitative data by the Unweighted Pair-Group method (UPGMA) (Nei, 1978) clustering using the sequential agglomerative hierarchical nested (SAHN) program and tree plot of Numerical Taxonomy Multivariate Analysis System package (NTSYS-pc) software, version 2.1.
SSR primers that showed distinct and scorable DNA bands were considered for analysis. The generated DNA bands were scored based on their band marker sizes. The software package Power Marker version 3.25 (Liu and Muse, 2005) was used for the following statistics analysis: sample size, major allele frequencies, number of alleles per polymorphic locus, and Polymorphism Information Content (PIC). The similarity matrix was subjected to UPGMA by selecting the SAHN program and tree plot of NTSYS.
RESULTS
Qualitative traits
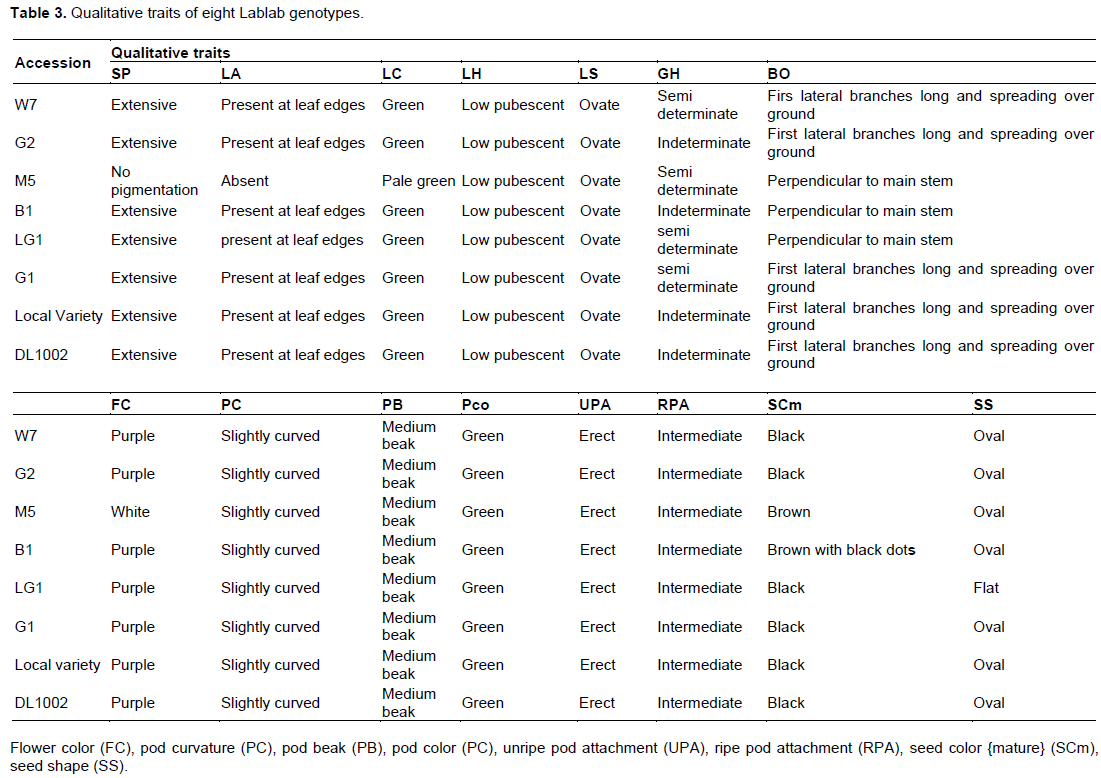
The Lablab genotypes were evaluated for 15 qualitative traits as listed in Table 3. There was no variation in seven of the qualitative traits evaluated in terms of leaf hairiness which was low pubescent, ovate leaf shape, their pod curvature was slightly curved, medium pod beak, green pod color, the attachment of unripe pod was erect and that of ripe pods was intermediate. However, there was variation in a few other traits. Genotypes W7, G1, B1, LG1, M5, Local Variety and DL1002 had extensive stem pigmentation, presence of leaf anthocyanin and green leaf color whereas genotype M5 had no stem pigmentation, no leaf anthocyanin and pale green leaf color. Genotype M5 had white flowers while all the other seven genotypes exhibited purple flower color. On the color of the seed, there was variation in that M5 had brown seeds; B1 had brown seeds with black dots while the other six genotypes had black seeds. There was also a variation in terms of leaf color and leaf anthocyanin in that, M5 had pale green leaves while the other seven genotypes had green leaves, M5 had no leaf anthocyanin which was present in all the other seven genotypes. There was variation in terms of seed shape in that seven of the varieties had oval seed shape expect LG1 which had flat seeds. There was also variation in the growth habit in that, genotype W7, M5, LG1, and G1 were semi determinate whereas G2, B1, Local variety and DL1002 were indeterminate. Since Lablab is largely intercropped with other crops especially maize, most farmers like the determinate genotypes (Shivachi et al., 2012; Gowda, 2008). Lastly, there was variation among the genotypes in terms of branch orientation where genotypes W7, G2, B1, and G1 had their first lateral branches long and spreading over the ground whereas in Genotypes G2, B1, Local variety and DL1002 their first lateral branches were perpendicular to the main stem (Table 3). The Lablab genotypes evaluated, exhibited some degree of phenotypic variation for some traits observed; Plate 1 shows the variation in terms of seed color and flower color.

Quantitative traits
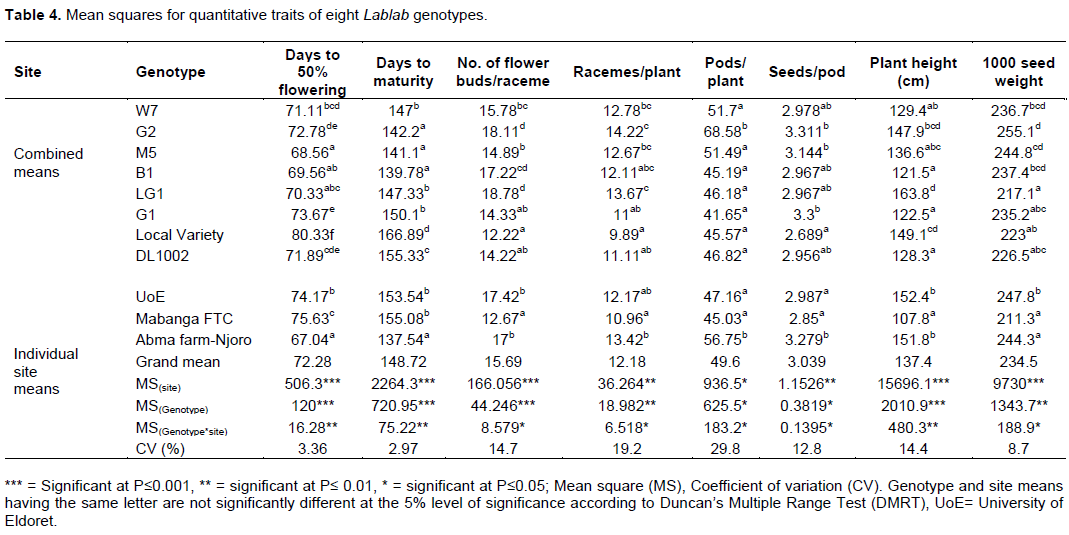
A total of eight quantitative traits were evaluated across three sites as discussed earlier. Sites (locations) were significant at P≤0.001 in; days to 50% flowering, days to maturity, flower buds per raceme, plant height and 1000 seed weight, a significant difference of P≤0.01 for racemes per plant and seeds per pod and a significant difference of P≤ 0.05 for number of pods per plant (Table 4). There was also a high significant difference in the means for the genotypes, P≤ 0.001, in terms of 50% flowering, days to maturity, flower buds per raceme and plant height. The interaction between genotypes and sites were significant at P≤ 0.01 in days to flowering, days to maturity and plant height. Other traits: flower buds per raceme, raceme per plant, pods per plant, seeds per pod and 1000 seed weight had a significance of P≤ 0.05. From the results, days to maturity was highly significant at P≤0.001, the new improved genotypes took shorter time to mature than the two checks. Genotype B1 had the shortest maturity time of 139 days whereas local variety had the longest maturity time of 166 days, a difference of 27 days. The number of flower buds per raceme ranged from 12 in local variety to 18 in genotypes G2 and LG1, plant height ranged from 122 cm in G1 to 163 in LG1, this is because genotype G1 is semi determinate while genotype LG1 is indeterminate. There was no correlation between number of flower buds per raceme and the number of pods per plant e.g. in genotype G2 and LG1 both had an average of 18.11 flower buds per raceme but had different number of pods per plant where genotype G2 had an average of 68.58 and genotype LG1 had an average of 46.18 (Table 4).
Microsatellite (SSR) analysis
Ten primer sets were used to amplify DNA extracts from the 8 genotypes. A total of 6 out of the 10 primers gave polymorphic bands (Table 5), 3 primers failed to amplify any product and 1 primer was monomorphic and therefore were not considered for further analysis. A total of 19 alleles were detected ranging from 2 to 5 per locus with a mean of 2 alleles per locus (Table 5). Genetic diversity was high ranging from 0.219 in LABRRT83 to 0.781 in LABRRT52 with a mean value of 0.542. The polymorphic information content (PIC) values also ranged from 0.195 in LABRRT 83 to 0.746 in LABRRT52 with an average of 0.489. Based on the PIC values, the most polymorphic primers were LABRRT52, LABRRT2 (Plate 2) and LABBRT77, respectively.
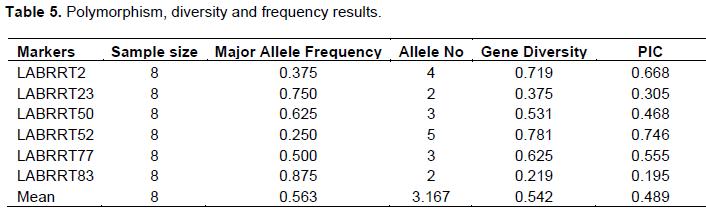

Cluster analysis
The genetic distance based on simple matching cluster analysis constructed from quantitative traits revealed that the closest genotypes were B1 and G1 while the longest distance was observed between G2 and W7&M5 (Figure 1). Using the mean similarity as cutoff, the genotypes were clustered in three categories: the first consisted of genotypes W7, M5, B1, G1 and DL1002; the second LG1 and Local variety, and the third only with G2, which showed high divergence compared to other genotypes, particularly due to higher means of raceme per plant, pods per plant and 1000 seed weight.
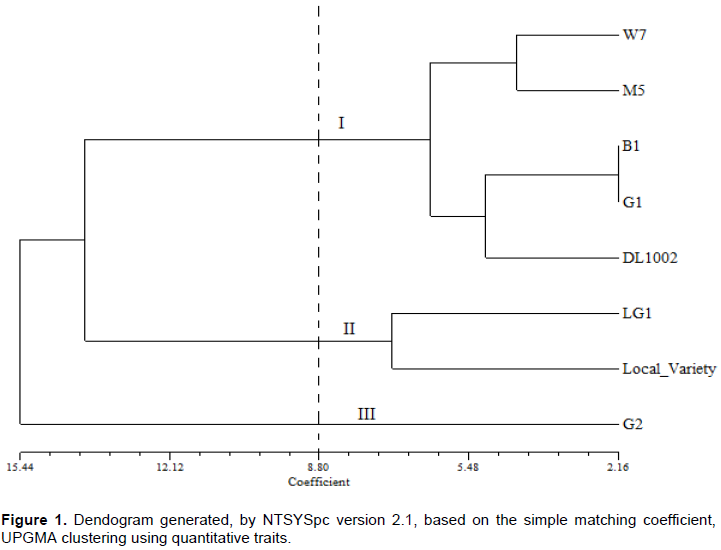
A second dendogram of 8 Dolichos genotypes based on SSR data was constructed (Figure 2). The genotypes were clustered into four categories: (I) consisted of M5, LG1 and W7, (II) B1 and G1, (III) G2, and (IV) Local variety and DL1002. The genetic distance revealed that the closest genotypes were LG1 and W7 with the longest distance being genotype LG1&W7 and DL1002 (Figure 2).
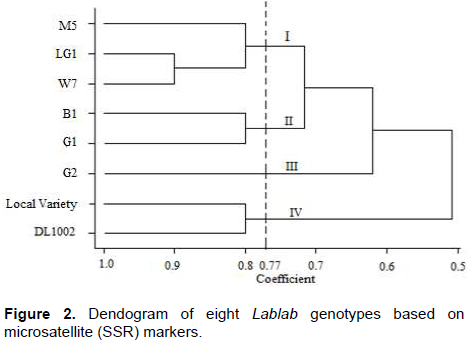
A third dendogram was generated using combined morphological and SSR data (Figure 3). The eight Lablab genotypes were grouped into four categories: (I) B1 and LG1, (II) G2, (III) G1, W7 and M5, and lastly (IV) DL1002 and Local variety. The dendogram also revealed that the closest genotypes were G1 and W7 (Figure 3).
DISCUSSION
Morphological characterization
Morphological traits are useful tools for preliminary evaluation of genotypes since they offer a fast and useful approach for assessing the extent of diversity. The estimation of different morphological traits studied among the improved Lablab genotypes and their checks in the present study revealed the existence of some level of morphological diversity though more variability was noted in quantitative traits that in qualitative trails. This study corroborates similar studies by Kamotho et al. (2017). This implies that the new genotypes constitute a pool of germplasm with adequate genetic variability.
Qualitative traits
The study revealed that there was variation in most of the qualitative traits evaluated though some qualitative traits did not differ significantly across the three sites. These include: stem pigmentation, leaf anthocyanin, leaf color leaf hairiness, branch orientation, flower color, pod curvature, pod color, pod beak, pod attachment, seed color and seed shape. These results also concur with findings by Shivachi et al. (2012). From the results, there could be a correlation between stem pigmentation and leaf anthocyanin, that is, M5 which had no stem pigmentation did not have leaf anthocyanin while the other genotypes had extensive stem pigmentation and subsequently had leaf anthocyanin at the leaf edges, this was also reported by Shivachi et al. (2012), Kar and Gogoi (2006), and Adebisi and Bosch (2004). All the genotypes that were purple flowered were also black seeds while white flowered genotype (M5), was brown seeded (Table 5). These results confirm those of Ewansiha and Chiezey (2007). All the genotypes showed great resistance to pests and diseases except local variety which showed mild resistance. This observation could be explained by the fact that the six genotypes (M5, G1, G2, LG1, B1 and W7) had been bred for pest and disease resistance. These variations observed in qualitative traits can be used in the selection of genotypes for further breeding programs.
Quantitative traits
Most of the quantitative traits evaluated were highly significant at (P≤ 0.001), except for number of racemes per plant and number of seeds per pod which were significant at P≤ 0.01 and number of pods per plant which was significant at P≤0.05. There was high diversification with notable variations in, days to 50% flowering, maturity period, number of flower buds per raceme, racemes per plant, number of seeds per pods, plant height, and yield among other agronomic traits. This result also conforms to findings from Kamotho et al. (2017) and Gowda (2008). Days to 50% flowering varied from 68 to 80 days with genotype M5 with the least days and Local variety with the most days, respectively. This result conforms to results gotten by Arup and Subrata (2010) and Ayisi et al. (2006) who reported 57-115 and 63-75 days, respectively. Consequently, there was a notable variation in terms of days to maturity ranging from 142 to 166 days, with genotype M5 taking least days and Local variety taking the most days to mature, respectively. Nonetheless, the maturity days lie within the recommended period of between 90 and 210 days (Adebisi and Bosch, 2004). Number of days to maturity is a key factor that determines farmers’ adoption where most farmers prefer early maturing genotypes as opposed to late maturing once. There was also a slight variation in terms of number of seeds per pod with a range of 2 to 3 which represented a significance of 99.95%. Several investigators have reported a seed range of 2 of 6 seeds per pod of the same crop (Rai et al., 2010; Kar and Gogoi, 2006; Nandi et al., 2000). Significant difference was also noted in plant height with a range of 121 to 163 cm with genotype B1 being the shortest and genotype LG1 being the longest, respectively. In Kenya, most farmers’ grow Lablab as an intercrop with maize (Okumu and Wanjekeche, 2005) thus the short and determinate genotypes are popular. There was also notable significant difference in terms of 1000 seed weight with a range of 217 to 255 g where genotype LG1 had the lowest weight and genotype G2 had the highest weight. There could be a relationship between seed weight and shape of the seed. This is because, genotype G2 which had the highest weight had rounded seeds that were well filled compared to genotype LG1 which had the least weight and had flat seeds that were not well filled. From the cluster analysis (Figure 1) based on simple matching coefficient constructed from quantitative traits revealed that the closest genotypes were B1 and G1. Using the mean similarity as cutoff, the genotypes were clustered into three groups: (I) W7, M5, B1, G1, and DL1002; (II) LG1 and Local variety and (III) G2. Cultivated Lablab is morphologically diverse with large variations (Kamotho et al., 2017). Such diversity enhances the direct selection of these traits in developing combinations for breeding programs.
Genetic diversity based on SSR markers
Lack of amplification of an allele in certain genotypes might be due to the result of divergence in the sequences flanking the microsatellite or simply the production of an undetectable amount of PCR product, creating a null allele (Smulders et al., 1997; Lavi et al., 1994). The number of alleles produced by different primers from the results ranged from 2 to 5 with an average of 3.2 alleles per primer. This result concurs with findings by Shivachi et al. (2012) and Wang et al. (2007) who reported mean amplifications of 3.5 and 3.6 alleles per primer. In similar studies, on common beans involving microsatellites, Arunga et al. (2015), Jose et al. (2009) and Maras et al. (2008) reported mean amplifications of 2.17, 7.8 and 7.1 alleles per primer, respectively. Polymorphism information content is a closely related diversity measure (Botstein et al., 1980). The more the number of alleles uncovered by a marker, the higher its PIC. Therefore, the polymorphic information content (PIC) value was calculated to characterize the capacity of each primer to detect polymorphic loci which ranged from 0.195 to 0.746 in the current study. This concurred with results by Arunga et al. (2015), Somta et al. (2009) and Cabral et al. (2011) who recorded PIC values of between 0.17 and 0.41, 0.049 and 0.883 and 0.11 and 0.5, respectively. The result showed that most of the primers were found to be highly informative and can be used to study phylogenetic relationship and genetic diversity in future. Thus, the three markers: LABRRT52, LABRRT2 and LABRRT77 with 0.746, 0.668 and 0.555 PIC values should be recommended for use in future Dolichos Lablab diversity studies. The allele frequency of all the primers was generally below 0.95 indicating that they were all polymorphic in character (Asare et al., 2011). This was slightly lower than findings from Datta et al. (2010) who reported an allelic frequency of 1.3. Gene diversity was high ranging from 0.219 to 0.781 with a mean value of 0.542, comparing favorably with the results obtained by Asare et al. (2011) although the mean value was slightly high (0.58). The results therefore indicate genetic diversities in the genotypes under this study and thus should be exploited further in future breeding program.
Comparing genetic distance of the Lablab genotypes based on morphological and SSR markers
The comparison of morphological versus SSR and combined dendograms results suggest that most of the genotypes for both dendograms had similar grouping patterns and that some morphological clusters were confirmed by SSR analysis. These comparison of morphological and SSR maker clustering pattern gives information on the amount of variability found between the genotypes and hence helps in identifying the best method of assessing diversity. Compared to phenotypic traits, molecular markers have the advantage of not being influenced by the environment, specific, reliable and wider range of genome sampling but have the disadvantage of accessing the genome as a whole and not only the regions responsible for the expression of traits of interest.
CONCLUSION AND RECOMMENDATION
Both morphological and SSR markers were able to group the Lablab accessions into distinct groups, though their genetic variability is relatively low thus it needs to be expanded through rigorous breeding programs. Since morphological markers, especially quantitative traits can easily be influenced by external factors like environmental conditions, it is always important to back up such findings with molecular markers which are increasingly becoming available for use. Since SSR markers specific to Lablab have been developed, it is therefore necessary to characterize all the Lablab accessions in the country at molecular level which will form a good basis for further development of breeding and selection programs.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGMENTS
This work was funded by a grant from Kirk House Trust through University of Eldoret. The authors acknowledge the assistance from KEPHIS Muguga for allowing us to use their facility for Microsatellite analysis. They also appreciate the management of Mabanga Farmer Training Centre and Abma Farm Njoro for allowing them to establish our trials on their farms. Lastly, they are grateful to the entire technical staff of Biotechnology Department, University of Eldoret for their support and technical assistance.
REFERENCES
|
Adebisi A, Bosch CH (2004). Lablab purpureus (L) sweet in: Grubben, GJH Denton, OA. (Editors). TROT A2:=Vegetablesles/Legumes. Wageningen, Netherlands pp. 343-348. |
|
|
Arunga EE, Kinyua MG, Ochuodho J, Chepkoech E (2015). Genetic diversity of determinate French beans grown in Kenya based on morpho-agronomic and simple sequence repeat variation. Journal of Plant Breeding and Crop Science 7(8):240-250. |
|
|
Arup C, Subrata D (2010). Characterization and identification of selection indices of pole type dolichos bean. Vegetable crops research bulletin 73:33-45. |
|
|
Asare PA, Galyuon IKA, Sarfo JK, Tetteh PJ (2011). Morphological and molecular based diversity studies of some cassava (Manihot esculenta crantz) germplasm in Ghana. African Journal of Biotechnology 10(63):13900-13908. |
|
|
Ayisi KK, Bopape MP, Pengelly BC (2006). Assesment of variation in growth and yield of diverse lablab (Lablab purpureus) germplasm in Limpopo Province, South Africa. CAB Abstract. |
|
|
Burkill H (1995). The useful plants of West Tropical Africa. Royal Botanical Gardens, kew 3:857. |
|
|
Botstein D, White RL, Skolnick M, Davis RW (1980). Construction of a genetic linkage map in man using restriction fragment length polymorphisms. American Journal of Human Genetics 32:314-331. |
|
|
Cabral PDS, Soares TCB, Lima ABP, Miranda FD, Souza FB, Gonclave LSA (2011). Genetic diversity in local and commercial dry bean (Phaseolus vulgaris?) accessions based on microsatellite markers. |
|
|
Datta S, Mahfooz S, Singh P, Choudhary AK, Singh F, Kumar S (2010). Cross genera amplification of informative microsatellite markers from common bean and lentil for the assessment of genetic diversity in pegeon pea. Physiology and Molecular Biology of Plants 16:123-134. |
|
|
Ewansiha SU, Chiezey U (2007). Morpho-phonological variation in Lablab purpureus. Tropical grasslands 41: 277-284. |
|
|
Girish G, Gowda MB (2009). Inheritance of qualitative characters in Dolichos bean Lablab purpureus L. Sweet Environmental Ecology 27(2):571-580. |
|
|
Gomez JO, Blair MW, Frankow-Lindberg BE, Gullberg U (2004). Molecular and phenotypic diversity of common bean landraces from Nicaragua. Crop Sciience 44(4):1412-1418. |
|
|
Gowda BM (2008). Genetic enhancement of Dolichos Bean Through Intergration of conventional breeding and molecular approaches-Final report. University of Agricultural science, Bangalore. |
|
|
Innes J (2004). Molecular marker techniques for crop improvement. Course Manual pp. 4-63. |
|
|
Jose CF, Mohammed MMS, Thomas G, Varghese G, Selvarj N, Dorai M (2009). Genetic diversity and conservation of common bean (Phaseolus vulgaris L. Fabeaceae) land races in Nilgras. Current science 97(2). |
|
|
Kamau EM, Kinyua MG, Gohole L, Kiplagat O (2010). Screening of local lablab (Lablab purpureus) accessions for resistance to cowpea aphid (Aphis craccivora Koch). KARI-Kenya. |
|
|
Kamotho GN, Muasya RM, Kinyua MG (2017). Assessment of phenotypic diversity of Kenyan dolichos bean (Lablab purpureus L. sweet) germplasm based on morphological markers. International Journal of Agriculture, Environment and Bioresearch 2(6). |
|
|
Kamotho GN, Kinyua MG, Muasya RM., Gichuki ST, Wanjala BW, Kimani E and Kamau EM (2016). Assessment of genetic diversity of Kenyan dolichos bean (Lablab purpureus l. sweet) using simple sequence repeat (SSR) markers. International Journal of Agriculture, Environment and Bioresearch 1(1). |
|
|
Kamotho GN, Kinyua MG, Muasya RM, Kimani EN, Orwa, DO (2010). A baseline survey on production, Utilization and Marketing Constraints of Lablab bean: Impact on Lablab Bean Improvement in Kenya. International Journal of Professional Pactice 1:21-29. |
|
|
Kar A, Gogoi HK (2006). Study of bean variability in Arunachal Pradesh. Bulletin of Arunachal Forestry Research 22(1&2):40-44. |
|
|
Khourya CK, Bjorkman AD, Dempewolf H, Ramirez-Villegasa J, Guarino L, Jarvisa A, Rieseberg LH, Struik PC (2014). A look at 10 orphan crops and their benefits. Increasing Homogeniety in Global Food Supplies and the Implications for Food Security.Proceedings of the national Academy of Sciences. |
|
|
Kimani EN, Wachira FN, Kinyua MG (2012). Molecular diversity of Kenyan Lablab bean (Lablab purpureus (L.) Sweet) accessions using amplified fragment length polymorphism markers. American Journal of Plant Science 3:313-321. |
|
|
Kinyua MG, Orwa D, Kimani E, Kamotho G (2008). Survey of Dolichos bean (Lablab purpureus) Production systems, Utilization, Marketing and the collection and Characterization of gemplasm in Kenya. Proceeding of the International Dolichos meeting, Arusha, Tanzania. |
|
|
Konstantinos T, Koutira O, Papadopouios II, Tokatlidis IS, Tamoutsidis EG, Vasiliki P M, Sotirious MK (2008). Genetic diversity in bean populations based on Random Amplified Polymorphic DNA markers. Biotechnology 7(1):109. |
|
|
Kumar H, Ghawade SM, Shivaputra, Meghwal ML (2018). Effect of Gamma Radiations on Growth, Yield and Quality Traits of Dolichos Bean (Lablab purpureus L.). International Journal of Current Microbiology and Applied Sciences ISSN: 2319-7692. Special Issue-6:853-859. |
|
|
Lavi UM, Akkaya A, Bhagwat E, Lahav-Cregan PB (1994). Methodology of generation and characteristics of simple sequence repeat DNA markers in avocado (Persea americana M.). Euphytica 80:171-177. |
|
|
Liu K and Muse SV (2005). PowerMarker: An integrated analysis environment for genetic marker analysis. Bioinformatics 21(9):2128-2129. |
|
|
Maass BL, Knox MR, Venkatesha SC, Angessa TT, Ramme S, Pengelly BC (2010). Lablab purpureus-A Crop Lost for Africa? Tropical Plant Biology 3(3):123-135. |
|
|
Maras M, Jelka SV, Javornik B, Meglic V (2008). The efficiency of AFLP and SSR markers in genetic diversity estimation and gene pool classification of common bean (Phaseolus vulgaris). ACTA Agriculturae Solvenica 91. |
|
|
Mureithi JG, Gachene CKK, Wamuongo JLW (2003). Legume cover crops research in Kenya: Experiences of the legume research network project. KARI Technical Note 12. |
|
|
Murphy AM, Colucci EP (2009). A tropical forage solution to poor quality ruminant diets: A review of Lablab purpureus. Livestock Research for Rural Development (11)2. |
|
|
Nandi A, Tripathy P, Lenka D (2000). Genetic divergence in hyacinth bean (Dolichos lablab). Indian In: Journal of Agricultural Sciences 70(7):450-451. |
|
|
National Research Council (2006). Lablab.Lost crops of Africa: Volume II: Vegetables. Washington DC: The National Academies Press. |
|
|
Nei (1978). Molecular evolutionary genetics. Evolutionary Biology pp. 159-164. |
|
|
Okumu M, Wanjekeche E (2005). Grow dolichos (njahe): to improve your soil and get more money. KARI-Kitale. |
|
|
Pengelly B, Maass L (2001). Lablab purpureus (L.) Sweet-Diversity, potential use and determination of a core collection of this multipurporse tropical legume. Genetic Resources and Crop Evolution 3(48):261-271. |
|
|
Rai N, Ashish K, Singh K, Singh M, Datta D, Rai M (2010). Genetic relationship among Lablab (Lablab purpureus) genotype cultivars from different races based on quantitative traits and Random Amplified Polymorphic DNA (RAPD) marker. African Journal of Biotechnology 9(2):137-144. |
|
|
Ravinaik K, Hanchnamani CN, Patil MG, Imamsaheb SJ (2015). Evaluation of dolichos genotypes (Dolichos lablab L.) under North eastern dry zone of Karnataka. The Asian Journal of Horticulture 10(1):49-52. |
|
|
Shivachi A, Kinyua MG, Kiplagat KO (2012). MIcrosettlite analysis of selected Lablab purpureus genotypes in Kenya. Rwanda journal ISSN 2305-2678 28(E). |
|
|
Smulders MJM, Bredemeijer G, Rus-kortekass W, Arens P, Vosman B (1997). Use of short microsatellites from database sequences to generate polymorphisms among Lycopersicon esculentum cultivars and accessions of other Lycopersicon species. Theoretical and Applied Genetics 97:264-272. |
|
|
Somta P, Sommanas W, Srinives P (2009). Molecular diversity assessment of AVRDC- The world vegetable centre elite Parental Mung Beans. Breeding Science 59:149-157. |
|
|
Wang M, Morris JB, Barkley NA, Dean RE, Jenkins TM, Pederson GA (2007). Evaluation of genetic diversity of the USDA Lablab purpureus germplasm collection using simple sequence repeats markers. Journal of Horticultural Science and Biotechnology 82(4):571-578. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


