Full Length Research Paper

ABSTRACT
Understanding the genetic variability, heritability and genetic advance of traits in any plant population is an important pre-requisite for breeding program. The experiment was conducted to assess the magnitude of genetic variability, heritability and genetic advance of 24 maize inbred lines for 16 quantitative traits. The field experiment was conducted during 2016 cropping season at Jimma Agricultural Research Center (JARC). Alpha lattice (0, 1) design with three replications and nine blocks was used. Analysis of variance showed high significance (P<0.01) differences among genotypes for all traits studied except tassel size. The genotypic coefficient of variation (GCV) for all traits studied was smaller than the phenotypic coefficient of variation (PCV), indicating the significant role of environment in the expression of traits studied. The estimates of PCV and GCV was high for grain yield, thousand kernel weight, ear height, ear diameter, anthesis and silking interval and plant aspect. Heritability estimates ranged from 9.15 for tassel size to 96.02 for thousand kernel weight. Estimates of genetic advance as percent of mean at 5% selection intensity ranged from 2.76% for days to maturity to 50.69% for grain yield. High heritability along with high genetic advance was obtained for plant height, ear length and 1000-kernel weight, indicating the predominance of additive gene effects in controlling the traits and effective selection on the basis of these traits would be absolutely useful for the improvement of inbred lines. Therefore, it could be recommended that due emphasis should be given for these traits for the improvement of maize inbred lines.
Key words: Heritability, genetic variability, genetic advance, inbred lines.
INTRODUCTION
Maize (Zea mays L, 2n=2x=20), a member of the grass family Gramineae (Poaceae), is one of the oldest cultivated crops. Maize is predominately cross pollinated by wind, but self-pollination is also possible (Sleper and Poehlman, 2006). Maize is the most important crop worldwide and basic trade product recurring ingredient for millions of people in Sub-Saharan Africa (Nzuve et al., 2013). Maize has also become the most most important staple food in rural Ethiopia (Abate et al., 2015). In addition, maize is also gaining popularity in Ethiopia for its demand for stover as animal fodder and a source of fuel for rural families. In Ethiopia, maize as food constitutes about 20% consumption and constitutes about 13% as feed in total cereals (FAO, 2013). It contributes about 29% of the calorie intake from total cereal consumption, followed by wheat and teff which is contributing about 21 and 17%, respectively (FAO, 2013).
Maize is the global leading cereal in terms of annual production which is about 1040.21 million tons, followed by wheat with production of 748.24 million tons and third in area under cultivation among all cereal crops. In Ethiopia, maize ranks second after teff in area coverage and first in total production (Abate et al., 2015). In Ethiopia, maize grows under a wide range of environmental conditions from 500 to 2400 m.a.s.l. The mid- altitude, sub-humid agro-ecology is, however, the most important maize producing environment in the country (Kebede et al., 1993). Though Ethiopia compares favorably with the main maize producing country in Sub Saharan Africa, the country is yet to achieve its potential in terms of production because of the old varieties dominating the seed system in the country, many biotic and abiotic factors, lack of genetically diverse source materials and little success in developing high yielding hybrids and synthetic varieties for different agro ecologies of the country (Abate et al., 2015). In particular, most of the varieties grown in southwestern highland of the Ethiopia are low yielding local cultivars with very tall plant ear, resulting in root and stalk lodging.
Information on the nature and magnitude of variability and heritability in a population is one of the prerequisites for successful breeding program in selecting genotypes with desirable characters (Dudly and Moll, 1969). Genetic improvement in traits of economic importance along with maintaining sufficient amount of variability is always the desired objective in maize breeding programs. In order to improve the genetic diversity of local germplasm, it is important to know the extent of already existing genetic variations in the material. The productivity and quality of maize are assured through initially evaluating, identifying and properly selecting of promising parental lines from available maize inbred lines.
In Ethiopia, maize improvement started half a century ago (Mosisa et al., 2002). During the late 1960s and early 1970s, several promising genotypes of east African origin were introduced and evaluated at different locations. Different studies were conducted to elucidate the nature and magnitude of genetic variability among maize inbred lines which identified traits like ear length, ear diameter, kernel number per row, ears per plant, 100 seed weight and rows per ear as potential selection criteria in a breeding program. For instance, Aman et al., (2015) identified traits such as kernels row per ear, grain yield, kernels number per row, 1000-kernel weight, ear diameter, ear length, days to anthesis, days to silking,
plant height, ear height, days to maturity and plant aspect as important traits in maize improvement. The national maize breeding program of Ethiopia also developed several maize inbred lines for use. However, little effort has been made; particularly in mid altitude area of the country to determine the variability of maize inbred lines considering their different morphological traits. Thus, the study was undertaken to determine the nature and magnitude of genetic variability among maize inbred lines.
MATERIALS AND METHODS
Description of the study site
The study was conducted at the Jimma Agricultural Research Center (JARC) during the main cropping season of 2016. The center is located in Ethiopia, Oromia National Regional State in Jimma zone, 343 km to the southwest of the capital city of the country, Addis Ababa. It is located around 07°46'N latitude and 36°47'E longitude coordinate and at an elevation of 1753 m.a.s.l. It represents the mid-altitude agro ecological zones which receive an annual rainfall of 1607.99 mm, with minimum and maximum mean temperatures of 16.60 and 31.48°C, respectively (JARC Agro-metrology department, 2016). The major soil type of the area is Chromic Nitosol and Combisol of upland and fluvisol of bottom land with a pH of about 5.2 (EIAR, JARC profile).
Experimental materials
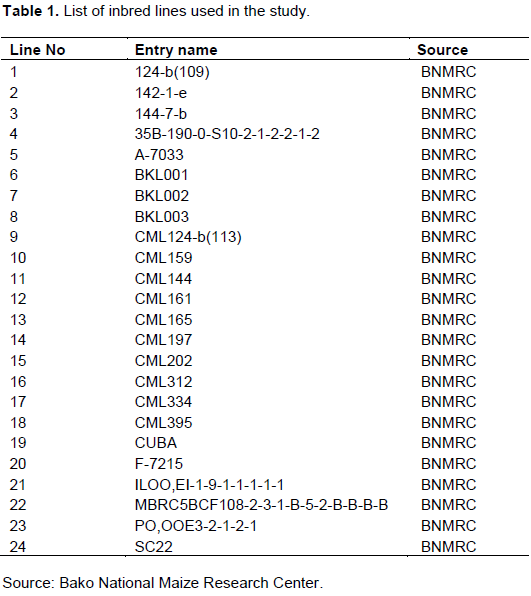
The experimental materials were obtained from Bako National Maize Research Center (BNMRC). Twenty-four commercial inbred lines were used. Description of the experimental materials used in this study is indicated in Table 1.
Experimental design and field management
The treatments were evaluated in alpha lattice design (Patterson and Williams, 1976) in three replications. Each treatment was planted in four rows of 5.1 m length with spacing of 0.75 m between rows and 0.30 m between plants within the rows. Two seeds were planted per hill and then thinned to one plant per hill to achieve standard plant density of 44,444 plants per hectare. The middle two rows were used for observations and data recording. Field management and other agronomic management practices were done following research recommendations for the area.
Data collected
Data were recorded on individual plant and plot bases for sixteen quantitative traits at the appropriate growth stage of the crop depending on the standard evaluation system of maize plant (IBPGR, 1991).
Data recorded on a plant basis
Plant height (cm)
The height of the ten randomly selected plants was measured from base of the plant to the base of the first branch of the tassel and the average of the ten plants recorded as plant height for the plot.
Ear height (cm)
The height of the ten randomly selected plants was measured from base of the plant to the node bearing the upper most ear of the same plant used to measure plant height and the average was recorded as ear height for the plot.
Leaf length (cm)
Leaf length was measured from five randomly selected leaves per plant in the leaf subtending the upper ear from the ligule to apex for all ten randomly selected plants after anthesis and the average was recorded for the plot.
Leaf width (cm)
Leaf width was measured for five randomly selected leaves per plant in the leaf subtending the upper ear at the widest point along its length for all ten randomly selected plants after anthesis and the average was for the plot.
Leaf area (cm2)
Is the average area of five randomly selected leaves per plant in the plot as the product of its length and width taken from each of 10 sampled plants, then multiplied by the correction factor k (k=0.75),
where LA= LW x LL x K.
Ear length (cm)
The length of ears harvested from the ten randomly selected plants was measured using ruler and the average was recorded for the plot. The same ears used to record ear diameter were used to record ear length.
Ear diameter (cm)
The diameter of ears harvested from the ten randomly selected plants was measured using Digital caliper and the average was recorded for the plot.
Number of kernel rows per ear
The total number of rows was counted in ten randomly taken ears and the average value was recorded as number of rows per ear.
Data recorded on plot basis
Days to 50% anthesis
Days to anthesis was recorded as the number of days from planting to the day when 50% of the plant in a plot started pollen shading.
Days to 50% silking
Days to silking was recorded as the number of days from planting when 50% of plants in the plot had their silks emerged 2-3 cm above the sheath.
Anthesis and Silking interval
Anthesis-silking interval was determined as the difference between the numbers of days to anthesis and silking.
Tassel size
Tassel size was recorded after milk stage using 3-7 scale; where 3, 5 and 7 indicate small, medium and large, respectively.
Plant aspect
Plant aspect was scored using 1-5 scale, where 1-indicates good (considering ear size, uniformity, disease infestation, husk cover, and so on) while 5 indicates poor genotype having undesirable ear characters.
Days to maturity
Days to maturity was recorded as the number of days from planting to the day on which 50% of the plants in the plot formed a black layer on sampled grains.
Thousand kernel weight (g)
After shelling, random sample of kernels from the bulk of each experimental unit was counted using a photoelectric seed counter and weighed in grams on sensitive balance after the moisture has been adjusted to 12.5%.
Grain yield (t/ha)
Field weight of all harvested ears was weighted and converted to grain yield using shelling percent of 80 percent. Grain yield was then determined in tons per hectare after adjusting moisture content of 12.5 percent using the following formula.
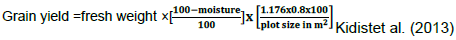
0.8 is shelling ratio, while 1.176 is constant.
Data analysis
The data collected for each character were subjected to analysis of variance (ANOVA) using statistical software, SAS version 9.3.
Estimation of variance components
Variance components were estimated to identify genetic variability among inbred lines. Error (σ2e), genotypic (σ2g) and phenotypes (σ2p) variances were calculated from expected mean squares of analysis of variance by adopting the formula suggested by Hallauer and Miranda (1988).
Error variance
σ2e = MSe
Where: MSe= mean square of error
Genotypic variance
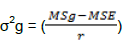
Where: MSg= mean square of genotype, MSe= mean square of error and r=number of replications
Phenotypic variance
σ2p= σ2e+ σ2g
Where: σ2e= error variance and
σ2g=genotypic Variance
Genotypic coefficient of variation (GCV)
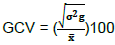
Where: σ2g= genotypic variance and 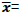 mean of the trait
mean of the trait
Phenotypic coefficient of variation (PCV)
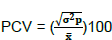
Where: σ2p= phenotype variance and mean of the trait
PCV and GCV values were categorized as low, moderate, and high values as indicated by Sivasubramanian and Menon (1973) as follows:
0-10% = low, 10-20% = Moderate and >20% = high
Heritability broad sense 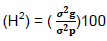
Where: σ2g= genotypic variance and σ2p= phenotypic variance.
Then it was categorized as low, moderate and high as indicated by Robinson et al. (1955).
0-30% =low, 30-60% =moderate and >60 =high
Genetic advance (GA) and genetic advance as per cent of the mean (GAM) were estimated as devised by Johnson et al., (1955), that is,
GA = kσpH2
Where: σp= the phenotypic standard deviation of the character, H2= broad sense heritability estimate and k = selection differential where k=2.063 at 5 % selection intensity.
Genetic advance as per cent of the mean was calculated to compare the extent of predicted advances of different traits under selection.
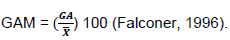
GA = genetic advance, and mean for the trait
The GA as percent of the mean was categorized as low, moderate and high according to Johnson et al. (1955) as follows.
0-10% =low, 10-20% =moderate and >20% =high
RESULTS AND DISCUSSION
Analysis of variances
There were highly significant (P£0.01) differences among the inbred lines for all studied traits, except for tassel size (Table 2). The present result is also in agreement with the findings of Taye (2014) and Mustafa et al. (2015). Thus, the genetic variability recorded in this study could be exploited by plant breeders to develop improved hybrid varieties.
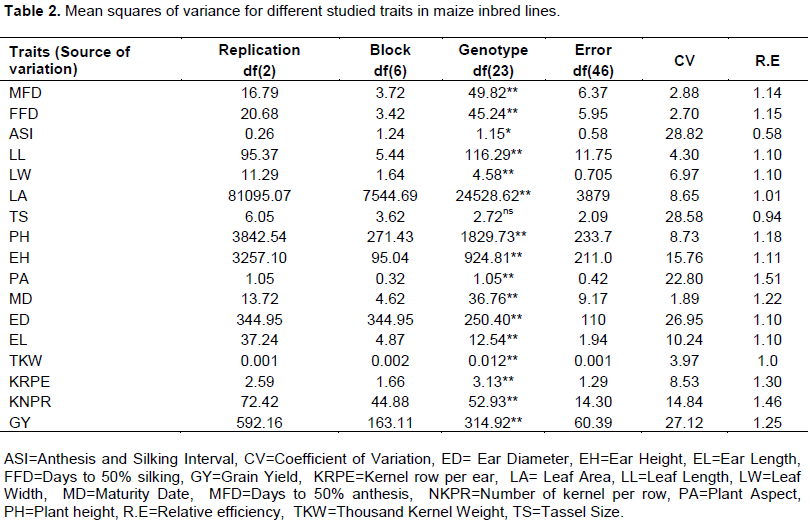
Mean and range values
The range and mean values for the 16 traits are presented in Table 3. The results indicated significant differences among the inbred lines for growth, yield and yield related traits. The mean grain yield ranged from 5.69 to 52.21 ton per hectare. Among the studied inbred lines 45.83% of inbred lines gave above the grand mean. Number of kernels per row ranged from 16.07 to 34.1307 with a mean value of 25.48 numbers. The range observed for kernel row per ears was 10.93 to 15.33 with overall mean of 13.31 rows. The maximum and minimum values of 1000-kernel weight were 0.43 and 0.20 t/ha respectively, with a mean value of 0.32t/ha.
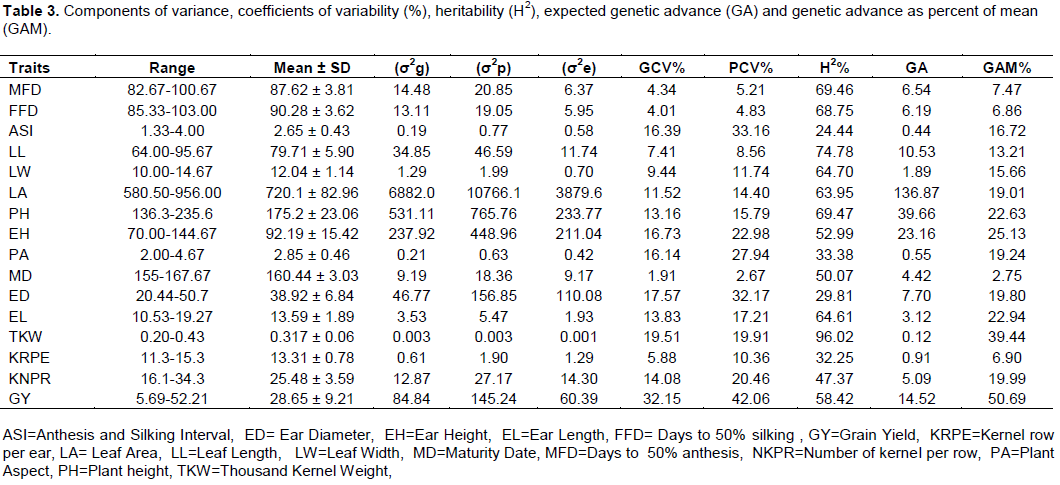
The mean plant and ear heights of the genotypes ranged from 136.33 to 235.67 cm and 70.00-144.67 cm respectively. The range observed for days to 50% silking was 82.67 to 100.67 days, with overall mean of 90.28 days. Days to 50% anthesis varied from 82.67 to 100.67, with overall mean of 87.62 days. Days to maturity varied from 155 to 167.67 days, with a mean of 160.44 days. Among total studied inbred lines 58.33% of inbred lines were indicated to exhibit early maturing day.
The inbred lines CML359, CML144 and PO, OOE3-2-1-2-1 were found to be superior in terms of grain yield per hectare, as well as in other important yield components. It is, therefore, suggested that these lines could be used for further improvement of maize inbred lines for yield targeted breeding strategy. Particularly, the wide variability observed for grain yield as a quantitatively inherent character among the genotypes means that, there is ample opportunity for selection in the genotypes for improvement of this important economic character. The range and mean values of the studied traits suggested the existence of sufficient variability among the studied inbred lines for the majority of considered characters and their considerable potential for improvement. Tulu et al. (2014) reported a wide range of variability of traits such as grain yield per hectare, row number per ears, number of kernels per row, ear length, ear diameter and 1000-kernel weight which is confirmed by the present study. Iqbal et al. (2015) also observed high range values for plant height and ear height in maize inbred lines they studied.
Genotypic and phenotypic coefficients of variation
Estimated variance components of PCV and GCV of the studied traits are presented in Table 3. High GCV was observed for grain yield (32.15). On the contrary, moderate values were observed for thousand kernel weight (19.51), ear height (16.73), ear diameter (17.57), anthesis and silking interval (16.39), plant aspect (16.14), number of kernels per row (14.08), ear length (13.83), plant height (13.16) and leaf area (11.54). The rest of the traits such as, leaf width (9.44), leaf length (7.41), kernel row per ear (5.88), days to 50% anthesis (4.34), days to 50% silking (4.01) and days to maturity (1.91) depicted low genotypic coefficient of variation.
High PCV was observed for grain yield (42.06), anthesis and silking interval (33.16), ear diameter (32.17), plant aspect (27.94) and ear height (22.98). On the contrary, moderate values were observed for number of kernels per row (20.46), thousand kernel weight (19.91), ear length (17.21), plant height (15.79), leaf area (14.40) and leaf width (11.74). The remaining traits such as, number of kernel rows per ears (10.36), leaf length (8.56), days to 50% anthesis (5.21), days to 50% silking (4.83) and days to maturity (2.67) showed a low phenotypic coefficient of variation.
A high range of PCV and GCV was noted for grain yield per hectare, anthesis and silking interval, ear diameter, ear height and plant aspect suggesting that these traits are under the influence of genetic control. Similarly, Kumar et al. (2014) and Nzuve et al. (2014) also reported the highest GCV and PCV for plant height, ear height, 1000 grain weight and kernel number per row. Hence, these traits can be relied upon; and simple and effective selection can be practiced for further improvement. The inbred lines showed adequate variability with regard to these traits, thus; genetic improvement could be achieved through selection for these traits (Vashistha et al., 2013; Mustafa et al., 2015). Low PCV and GCV were observed for leaf width, kernel row per ears, leaf length, days to 50% anthesis, days to 50% silking, and days to maturity and this could probably be attributed to the phenotypic plasticity and also presence of both positive and negative alleles in the maize genotypes leading to low genotypic variation. Similar results were reported by Manju et al. (2002) and Shakoor et al. (2007).
Phenotypic coefficient of variation was found to be higher than the genotypic coefficient of variation for all traits. Similarly, Yusuf (2010) also reported higher phenotypic coefficients of variations than the genotype coefficient of variations for all studied traits. However in this study, the two values differ slightly, indicating less influence of environmental factor on gene expression for the traits. Moreover, the difference between PCV and GCV was low for traits like thousand kernel weight, days to maturity, days to 50% silking, days to 50% anthesis, leaf length, leaf width, plant height, leaf area, leaf length and kernel rows per ears. This implies, less environmental influence on these traits, which ensured practically higher chance for selection. Ear height, number of kernels per row and grain yield showed moderate values between PCV and GCV, which guarantees average chance for selection. Anthesis and silking interval, ear diameter and plant aspect relatively achieved high difference among phenotypic and genotypic coefficient of variation and hence these traits provide practically, less chance for selection, due to higher influence of environmental factors.
Heritability and genetic advance
Broad sense heritability (H2), an estimate of the total contribution of genetic variance to the total phenotypic variance, ranged from 24.44 for anthesis-silking interval to 96.02 for 1000-kernel weight (Table 3). Higher heritability estimates were scored for 1000-kernel weight (96.02), leaf length (74.79), plant height (69.47), days to 50% anthesis (69.46), days to 50% silking (68.75), leaf width (64.70), ear length (64.62) and leaf area (63.95). Moderate heritability estimates were observed for grain yield per hectare (58.42), ear height (52.99), days to maturity (50.07), number of kernels per row (47.38), plant aspect (33.38) and kernel row per ears (32.26). In contrast, ear diameter (29.82), anthesis and silking interval (24.45) had low heritability estimates.
The high heritability estimates suggest selection of such character could be fairly easy. Therefore, 1000-kernel weight, leaf length, plant height, days to 50% anthesis, days to 50% silking, leaf width and leaf area could easily be passed from one generation to the next then enhancing the efficiency of selection in maize improvement program. This indicated that the traits are under genetic control and the environmental factors did not greatly affect their phenotypic variation. Thus, conventional breeding for these traits could lead to maize improvement (Lule et al., 2012).
Moderate heritability estimates were observed for grain yield per hectare, ear height, days to maturity, number of kernels per row, plant aspect and kernel row per ear indicating these traits may respond positively to phenotypic selection. This is reliable with the result of Al-Tabbal and Al-Fraihat (2012) and Nzuve et al. (2014). The traits exhibited moderate heritability estimates could be improved through heterosis breeding or hybridization (Bello et al., 2012). Ear diameter, anthesis and silking interval indicated low heritability estimates. This implies selection is considerable difficult for such traits due to the masking effect of the environment on the phenotypic traits. The genetic advance as percent of mean (GAM) at 5% selection intensity ranged from 2.76% for days to maturity to 50.69% for grain yield per hectare (Table 3). There was high genetic advance expressed as a percent of mean for some traits like: grain yield (50.69%), 1000-kernel weight (39.44%), ear height (25.13%), ear length (22.94%) and plant height (22.64%). On the other hand, traits such as number of kernels per row (20.00%), ear diameter (19.79%), plant aspect (19.24%), leaf area (19.01%), anthesis-silking interval (16.72%), leaf width (15.67%) and leaf length (13.21%) had moderate genetic advance as percent of mean and the traits like days to 50% anthesis (7.47 %), number of kernel rows per ears (6.90%), days to 50% silking (6.86%) and days to 90% maturity (2.76%) had low genetic advance as percent of mean.
Genetic advance (GA) as a percentage of the mean was higher for traits such as grain yield per hectare, 1000-seed weight and ear height showing that, these traits are under the control of additive gene action. This is supported by the findings of Atnafu and Rao (2014) who reported high genetic advance for plant height, kernel rows per ears, 1000 kernel weight, ear height, and grain yield per hectare. The traits like days to maturity and days to 50% silking indicated low values of genetic advance as per cent of mean and which correspondingly indicated low value of genetic variation for the traits as indicated by low GCV and PCV values. This implies the importance of genetic variability in improvement through selection. This result is also confirmed by results of Fekadu (2014). Maruthi and Rani (2015) observed high genetic advance as per cent of mean for ear height, plant height, number of kernels per ears, ear length, ear diameter and 1000 -grain weight, which is generally in agreement with the result of the present study.
The estimate of GAM for grain yield was 50.69%. The current yield for inbred line CML359 was 5521 kg/ha. Therefore, whenever the best 5% high yielding inbred lines is selected as a parent, the mean grain yield could be improved by 2798.61 kg/ha. As a result, mean genotypic value of the new population of grain yield will be improved from 5521 to 8319.61 kg/ha per one selection cycle for the line CML359. In the same way, the estimate of genetic advance as percent of mean for 1000-kernel weight was 39.44% and CML359 lines indicated 0.41 kg/ha of 1000-kernel weight. Therefore, after one selection cycle performed for best performing inbred lines at 5% selection intensity, it will be advanced from 0.41 to 0.57 kg/ha. The present study revealed high heritability estimates coupled with the high expected genetic advance as per cent of mean for 1000-kernel weight, ear length and plant height and also moderate heritability estimates with higher genetic advance for grain yield and ear height. This indicated these traits could be improved more easily than the other traits through simple selection. Therefore, even if heritability estimates provide the basis for selection on phenotypic performance, the heritability estimates and genetic advance should be always considered simultaneously, as high heritability is not always associated with high genetic advance.
CONCLUSION
In this study, considerable amount of genetic variability among the studied inbred lines was observed. The maximum and minimum grain yield per hectare ranged from 5.22t to 0.57t for CML359 and CML159 inbred lines, respectively. PCV was found to be higher than the GCV for all traits. The highest phenotypic and genotypic coefficients of variation were observed for grain yield, anthesis-silking interval, ear diameter, plant aspect and ear height. Leaf length, days to 50% anthesis, days to 50% silking, number of kernel rows per ear and days to maturity had low phenotypic and genotypic coefficient of variation.
High broad sense heritability was recorded for 1000-kernel weight, leaf length, plant height, days to 50% anthesis, days to 50% silking, leaf width, ear length and leaf area which indicated that the variation observed was under genetic control and less influenced by environment. High genetic advance as a percent of mean was observed for grain yield per hectare, 1000-kernel weight, ear height, ears length and plant height. Inbred lines showed high heritability with high genetic advance for thousand kernel weight, ear length and plant height which implies that these traits are under additive gene action. These estimates suggested that selection on the basis of these traits is helpful for breeding program otherwise no genetic gain can be achieved.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENT
The authors specially thank Jimma Agricultural Research Center for their support in field experimentation and data recording.
REFERENCES
|
Abate T, Bekele S, Abebe M, Dagne W, Yilma K, Kindie T, Menale K, Gezahegn B, Berhanu T, Tolera K (2015). Factors that transformed maize productivity in Ethiopia. Food Security 7(5):965-981. |
|
|
Aman J, Bantte K, Alamerew S, Tolera B (2015). Evaluation of Quality Protein Maize (Zea mays L) Hybrids at Jimma, Western-Ethiopia. Journal of Forensic Anthropology 1:101. |
|
|
Atnafu B, Rao TN (2014). Estimates of Heritability, genetic advance and correlation study for yield and its attributes in maize (Zea mays L.). Journal of Plant Science 2(1):1-4. |
|
|
Al-Tabbal JA, Al-Fraihat AH (2012). Heritability studies of yield and yield associated traits in wheat genotypes. Journal of Agricultural Science 4(4):11. |
|
|
Bello OB, Ige SA, Azeez MA, Afolabi MS, Abdulmaliq SY, Mahamood J (2012). Heritability and Genetic Advance for Kernel Yield and its Component Traits in Maize (Zea Mays L.). International Journal of Plant Research 2(5):138-145. |
|
|
Dudly JW, Moll RH (1969). Interpretation and use of estimates of heritability and genetic variance in plant breeding. Crop Science 9: 257-267. |
|
|
Falconer DS, Mackay FC (1996). Introduction to Quantitative Genetics. Longman, New York P 464. |
|
|
Food and Agriculture Organization (FAO) (2013). FAOSTAT Database on Agriculture. Food and Agriculture Organization of United Nations. |
|
|
Fekadu K (2014). Genetic variability for yield and yield related traits in some maize (Zea mays L.) inbred lines in Central Highland of Ethiopia. Ms.c thesis submitted to the collage of Natural and Computational Sciences, Department of Biology, School of Graduate Studies Haramaya University. |
|
|
Hallauer AR, Miranda JB (1988). Quantitative genetics in maize breeding. 2nd ed. Iowa state Universality Press. |
|
|
International board for Plant Genetic Resources (IBPGR) (1991). Descriptors for maize. International maize and wheat Improvement centre, Mexico City/International board for Plant Genetic Resources, Rome. pp. 29-55. |
|
|
Iqbal J, Shinwari ZK, Rabbani MA (2015). Maize (Zea mays L.) Germplasm AAGRO Morphological Characterization Based on Descriptive, Cluster and Principal Component Analysis. Pakistan Journal of Botany 47:255-264. |
|
|
JARC Agro-metrology department (2016). Mean of ten years (2007-2016) Metrological data of Jimma agricultural research center unpublished. |
|
|
Johnson HW, Robinson H, Comstock RE (1955). Estimates of genetic and environmental variability in soybeans. Agronomy Journal 47:314-318. |
|
|
Kebede M, Gezahegne B, Benti T, Mossisa W, Yigzaw D, Assefa A (1993). Maize production trends and research in Ethiopia. Proceedings of the First National Maize Workshop of Ethiopia. Addis Ababa, Ethiopia pp. 142-154. |
|
|
Kidist A, Nigussie D, Habtamu Z (2013). Growth, Productivity and Nitrogen Use Efficiency of Maize (Zea Mays L.) as Influenced by Rate and Time of nitrogen Fertilizer Application in Haramaya District, Eastern Ethiopia. Ms.c thesis. |
|
|
Kumar GP, Reddy VN, Kumar SS, Rao PV (2014). Genetic Variability, heritability and genetic advance studies in newly developed maize genotypes (Zea mays L.). International Journal of Pure and Applied Bioscience 2(1):272-275. |
|
|
Lule D, Tesfaye K, Fetene M, De Villiers S (2012). Inheritance and association of quantitative traits in finger millet (Eleusine coracana Subsp. Coracana) landraces collected from eastern and south eastern Africa. International Journal of Genetics 2(2):12-21. |
|
|
Manju PR, Sreelathakumary I (2002). Genetic variability, heritability and genetic advance in hot chilli (capsicum Chinese jacq.). Journal of Tropical Agriculture. 40:4-6. |
|
|
Maruthi RT, Rani KJ (2015). Genetic variability, heritability and genetic advance estimates in maze (Zea mays L.) inbred lines. Journal of Applied and Natural Science 7(1):149-154. |
|
|
Mosisa W, Hadji T, Wonde A, Legese WA, Diallo Twumasi A, A. Guta A (2002). Developing low-N tolerant maize varieties for mid-altitude subhumid agro-ecology of Ethiopia. Integrated Approaches to Higher Maize Productivity in the New Millennium (No. CIS-4176. CIMMYT.). |
|
|
Mustafa HSB, Farooq J, Bibi T, Mahmood T (2015). Cluster and principle component analyses of maize accessions under normal and water stress conditions. Journal of Agricultural Sciences, Belgrade 60(1):33-48. |
|
|
Nzuve F, Githri S, Mukunya DM, Gethi J (2014). Genetic variability and Correlation studies of grain yield and related agronomic traits in maize. Journal of Agricultural Science 6(9):166. |
|
|
Nzuve F, Githiri S, Mukunya DM, Gethi J (2013). Analysis of genotype x environment interaction for grain yield in maize hybrids. Journal of Agricultural Science 5(11):75. |
|
|
Patterson H, Williams E (1976). A new class of resolvable incomplete block designs. Biometrika 63(1):83-92. |
|
|
Shakoor MS, Akbar M, Hussain A (2007). Correlation and path coefficient studies of some morpho-physiological traits in maize double crosses. Pakistan Journal Agricultural Science 4(4):213-216. View |
|
|
Sivasubramanian S, Menon M (1973). Heterosis and inbreeding depression in rice. Madras Agriculture Journal 60:1139. |
|
|
Sleper DA, Poehlman JM (2006). Breeding field crops (No. Ed. 5). Blackwell publishing. |
|
|
Taye A (2014). Genetic variability of yield and yield related traits in some maize inbred lines (Zea mays L.) developed for mid-altitude agro-ecology of Ethiopia. MSc thesis submitted to the collage of Natural and Computational Sciences, Department of Biology, School of Graduate Studies, Haramaya University. |
|
|
Tulu B (2014). Correlation and path coefficients analysis studies among yield and yield related traits of quality protein maize (QPM) inbred lines. International Journal of Plant Breeding Crop Science 1(2):6-17. |
|
|
Vashistha A, Dixit NN, Dipika, Sharma SK, Marker S (2013). Studies on heritability and genetic advance estimates in maize genotypes. Bioscience Discovery 4(2):165-168. |
|
|
Yusuf M (2010). Genetic variability and correlation in single cross hybrids of quality protein maize (Z. mays L.). African Journal of food, Agriculture, nutrition and development 10(2):2166-2175. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


