Full Length Research Paper

ABSTRACT
Manihot esculenta spp flabellifolia is a potential progenitor for cassava root protein content and yield improvement. Storage roots of cassava landraces are low in protein content due to the fact that past breeding objectives concentrated mainly on yield and resistance to diseases. The improvement of cassava through its wild progenitor is of importance for the full utilization of the potential of the wild progenitor. An interspecific F1 was crossed to a cultivated variety (MTAI - 8) to generate a backcross population. Root protein, yield and other quality traits were evaluated. High root protein content of 9.61%; fresh root yield of 60.00 ton ha-1; dry root yield of 34.75 ton ha-1; and dry matter content of 59.45% was found in this population. High broad-sense heritability was obtained for all the traits evaluated which is a good indicator that genetic improvement can be achieved in this population. This first backcross population had protein values higher than the earlier documented values in the landraces.
Key word: Cassava, Manihot esculenta ssp flabellifolia, interspecific, protein and yield.
INTRODUCTION
MATERIALS AND METHODS

RESULTS
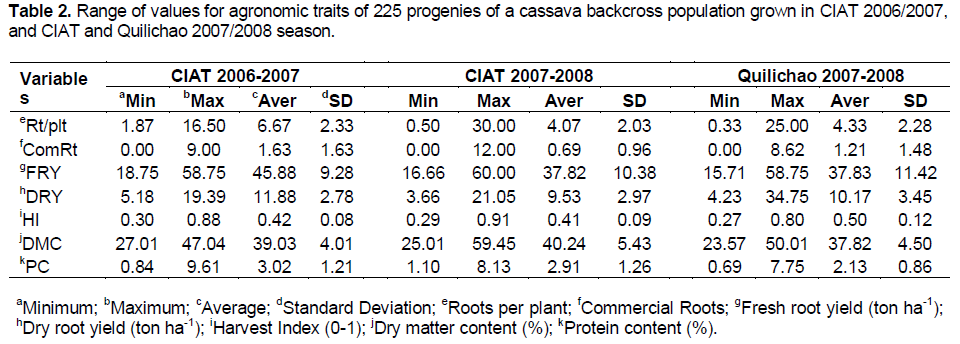

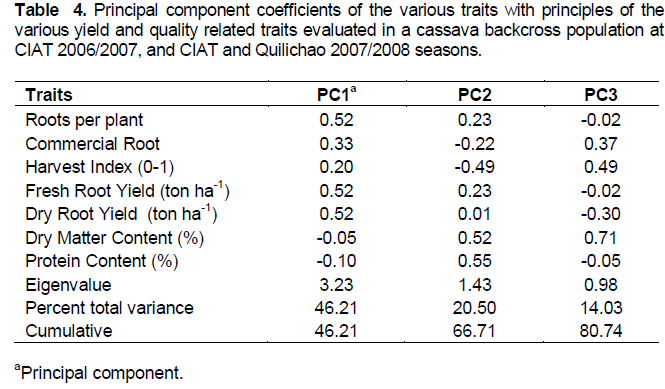

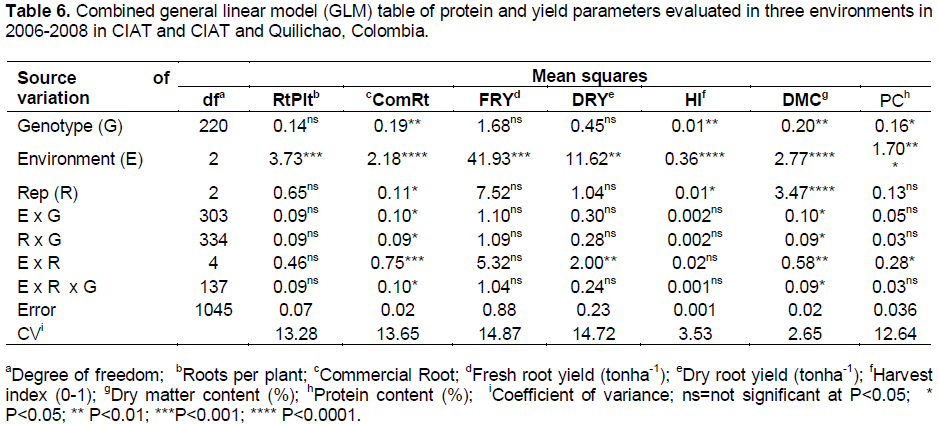
DISCUSSION
CONFLICT OF INTEREST
ACKNOWLEDGEMENTS
REFERENCES
| Agrobase (2005). Agrobase generation II user's manual. Agronomix Software, Manitoba. | ||||
| Akinbo O, Labuschagne M, Fregene M (2010). Embryo rescue as a method to develop and multiply a backcross population of cassava (M. esculenta Crantz) from an interspecific cross of Manihot esculenta ssp flabellifolia. Afr. J. Biotechnol. 9:7058-7062. | ||||
|
Aina OO, Dixon AGO, Akinrinde EA (2007). Additive main effects and multiplicative interaction (AMMI) analysis for yield of cassava in Nigeria. J. Biol. Sci. 7:796-800. Crossref |
||||
| Anonymous (1968). Tabla de Composicion de Pastes e otros Alimentos de Centro America. Publicacion No. E-40. Instituto Nacional de Centro America e Panama. | ||||
| Asiedu R, Bai KV, Terauchi R, Dixon AGO, Hahn SK (1992). Status of wide crosses in Cassava and Yam. In: Thotttapily, G. (ed.). Biotechnology; enhancing research on tropical crops in Africa: Proceedings of an international conference held at the Int. Instit. Trop. Agric, pp. 26-30 November 1990, IITA, Ibadan, Nigeria. | ||||
| Balyejusa-Kizito EB, Rönnberg-Wästiljung A-C, Egwang T, Gullberg U, Fregene M, Westerbergh A (2007). Quantitative trait loci controlling cyanogenic glucoside and dry matter content in cassava (Manihot esculenta Crantz) roots. Hereditas 144:129-136. | ||||
|
Carabali A, Belloti AC, Montaya-Lerma J, Fregene M (2010). Manihot flabillifolia Pohl, wild source of resistance to the whitefly Aleurotrachelus socialis Bondar (Hemiptera Aleyrodidae). Crop Prot. 29:34-38. Crossref |
||||
|
Ceballos H, Sánchez T, Chávez AL, Iglesias C, Debouck D, Mafla G, Tohme J (2006). Variation in crude protein content in cassava (Manihot esculenta Crantz) roots. J. Food Composit. Anal. 19:589-593. Crossref |
||||
| Ceballos H, Iglesias CA, Pérez JC, Dixon AGO (2004). Cassava breeding: opportunities and challenges. Plant Mole. Biol. 56:503-516. | ||||
|
Chávez AL, Sánchez T, Jaramillo G, Bedoya JM, Echeverry J, Bolanos EA, Ceballos H, Iglesias CA (2005). Variation of quality traits in cassava evaluated in landraces and improved clones. Euphytica143:125-133. Crossref |
||||
| CIAT (2004). Improved cassava for the developing world. Annual Report. 2003. | ||||
| CIAT (2003). Improved cassava for the developing world. Annual Report, 2002. | ||||
| CIAT (2002). Improved cassava for the developing world. Annual Report, 2001. | ||||
|
Crossa J (1990). Statistical analysis of multi-location trials. Advan. Agron. 44:55-85. Crossref |
||||
| Dixon AGO, Asiedu R, Bokanga M (1994a). Breeding of cassava for low cyanogenic potential: Problems, progress and perspective. Acta Horticulture 375:153-161. | ||||
| Dudley JW (1974). Seventy generations of selection for oil and protein in maize. Crops Science Society of America, Mandison, WI, USA. | ||||
|
Egesi CN, Ilona P, Ogbe FO, Akoroda M, Dixon A (2007). Genetic variation and genotype x environment interaction for yield and other agronomic traits in cassava in Nigeria. Agron. J. 99:1137-1142. Crossref |
||||
|
Fermont AM, van Asten PJA, Tittonell P, van Wijk MT, Giller KE (2009). Closing the cassava yield gap: An analysis from smallholder farms in East Africa. Field Crops Res. 112:24–36. Crossref |
||||
| Fregene MA, Blair MW, Beebe SE, Ceballos H (2007). Marker-assisted selection in common beans and cassava. In: Marker-Assisted Selection (MAS) in Crops, Livestock, Forestry and Fish: Current Status and the Way Forward. FAO Publishing, P. 471. | ||||
| Fregene M, Morante N, Sánchez T, Marin J, Ospina C, Barrera E, Gutierrez J, Guerrero J, Bellotti A, Santos L, Alzate A, Moreno S, Ceballos H (2006). Molecular markers for introgression of useful traits from wild Manihot relatives of cassava, marker-assisted selection (MAS) of disease and root quality traits. J. Root Crops 32:1-31. | ||||
| Griffing B (1956). Concept of general and specific combining ability in relation to diallel crossing systems. Aust. J. Biol. Sci. 9:463–493. | ||||
| Gomez KA, Gomez AA (1984). Statistical Procedures for Agricultural Research. Wiley-Interscience Publication, New York, pp. 304-306. | ||||
|
Hajjar R, Hodgkin T (2007). The use of wild relatives in crop improvement: A survey of developments over the last 20 years. Euphytica 156:1-13. Crossref |
||||
| Hock-Hin Y, Van-Den T (1996). Protein contents, amino acid compositions and nitrogen-to protein conversion factors for cassava roots. J. Sci. Food. Agric. 70:51-54. | ||||
| Iezzoni FA, Pritts MP (1991). Application of principal component analysis to horticulture research. Hort. Sci. 26:19-22. | ||||
|
Iglesias CA, Calle F, Hershey C, Jaramillo G, Mesa E (1994). Sensitivity of cassava (Manihot esculenta Crantz) clones to environmental changes. Field Crops Res. 36:213-220. Crossref |
||||
|
Jaramillo G, Morante N, Pérez JC, Calle F, Ceballos H, Aria B, Bellotti AC (2005). Diallel analysis in cassava adapted to the midaltitude valleys environment. Crop Sci. 45:1058-1063. Crossref |
||||
| Kawano K (1998). Socio-economic contribution of cassava varietal improvement to the small farmer communities in Asia. In: Howeler R. (ed.). Cassava Breeding, Agronomy and farmer participatory research in Asia. CIAT, Bangkok, Thailand. | ||||
|
Kawano K, Narintaraporn K, Narintaraporn P, Sarakarn S, Limsila A, Limsila J, Suparhan D, Sarawat V, Watananonta W (1998). Yield improvement in a multistage breeding program for cassava. Crop Sci. 38:325-332. Crossref |
||||
|
Krom M (1980). Spectrophotometric determination of ammonia: A study of modified Bethelot reaction using salicylate and cichloroisicyanurate. The Analyist 105:305-316. Crossref |
||||
| Lebot V (2009). Tropical root and tuber crops. soils, Plant Growth. Crop Prod. pp. 1-9. | ||||
| Magoon ML, Maini SB, Krishnan R (1973). Breeding for tuber quality in cassava. Trop. Root Tuber Crops Newslett. 5:27-29. | ||||
| Morantes N, Sanchez T, Fregene M (2002). Mining the Primary and Secondary Gene Pool: Protein and Dry Matter Yield Genes from Wild Manihot Species. CIAT annual report SB-2, #page=159. | ||||
| Nassar NMA, Dorea G (1982). Protein content in some cassava cultivars and its hybrid with wild Manihot species. Turrialba 32:429-432. | ||||
| Nassar NMA, Grattapaglia D (1986). Variabilidade de clones de mandioca em relação a fertilidade e aspectos morfológicos. Turrialba 36:555-559. | ||||
| Nassar NMA, Rafael da Silva J, Vieira C (1986). Hibridação interespecifica entre mandioca e espécies de Manihot. Cienc Cult 38:1050-1055. | ||||
|
Ngeve JM, Nukenine EN. Dixon AGO (2003). Reaction of cassava genotypes to the cassava mosaic disease in three distinct Agro-ecologies in Nigeria. J. Phytopathol. 151:101-107. Crossref |
||||
| Ngoan TN, Quyem TN, Kim H, Kawano K (1995). Cassava cultivars and breeding research in Vietnam. In: Cassava Breeding, Agronomy Research and Technology Transfer in Asia. Howeler R. (ed). CIAT, Bangkok, Thailand, pp. 253-261. | ||||
|
Njoku DN, Vernon G, Egesi CN, Asante I, Offei SK, Okogbenin E, Kulakow P, Eke-okoro ON, Ceballos H (2011). Breeding for Enhanced β-Carotene Content in Cassava: Constraints and Accomplishments. J. Crop Improv. 25:560-571. Crossref |
||||
|
Novozamsky I, Houba VJG, van Eck R, van Vark W (1983). A novel digestion technique for multi-element analysis. Commun. Soil Sci. Plant Anal. 14:239-249. Crossref |
||||
| Nuwamanya E, Baguma Y, Emmambux N, Taylor J, Patrick R (2010). Physicochemical and functional characteristics of cassava starch in Ugandan varieties and their Progenies. J. Plant Breed. Crop Sci. 2:001-011. | ||||
|
Ojulong H, Labuschangne MT, Herselman L, Fregene M (2008). Introgression of genes for dry matter content from wild cassava species. Euphytica. 164:163-172. Crossref |
||||
|
Okechukwu RU, Dixon AGO (2009). Performance of improved cassava genotypes for early bulking, disease resistance and culinary qualities in an inland valley. Agron. J. 101:1258-1265. Crossref |
||||
|
Okogbenin E, Porto MCM, Egesi C, Mba C, Espinosa E, Santos LG, Ospina C, Marín J, Barrera E, Gutiérrez J, Ekanayake I, Iglesias C, Fregene MA (2007). Marker-Assisted Introgression of resistance to cassava mosaic disease into Latin American germplasm for the genetic improvement of cassava in Africa. Crop Sci. 47:1895–1903. Crossref |
||||
| Okogbenin E (2004). Qualitative trait loci mapping of root quality traits, morphological characters, and early bulking in cassava (Manihot esculenta Crantz). PhD Thesis Department of Agronomy, University of Ibadan, P. 208. | ||||
| Okogbenin E, Fregene M (2002). Genetic analysis and QTL mapping of early root bulking in an F1 population of non-inbred parents in cassava (Manihot esculenta Crantz). Theoret. Appl. Genet. 106:58-66. | ||||
| Okogbenin E, Porto MCM, Dixon AGO (1998). Influence of planting season on incidence and severity of African cassava mosaic disease in the sub-humid zone of Nigeria. In: Akoroda M.O. and I.J. Ekanayake (eds.). Root crops and poverty alleviation. International Institute of Tropical Agriculture, Ibadan, Nigeria, pp. 388-392. | ||||
| Olasanmi B (2010). Marker-assisted selection for improvement of trait associated with high and early root productivity in cassava (Manihot esculenta Crantz). PhD thesis, Department of Agronomy, University of Ibadan, Ibadan, Nigeria. P. 168. | ||||
| Pérez JC, Ceballos H, Lenis JI, Ortega E, Morante N (2002). Heritability of agronomically relevant traits in cassava. In: Taylor N.J., F. Ogbe, Fauquet C.M. (eds) Cassava, an ancient crop for modern times food, health, culture. A paper presented at 5th international scientific meeting of the cassava biotechnology Network, Danforth Plant Science Centre St. Louis, MO, USA, p. 218. | ||||
|
Rolland-Sabaté A, Sánchez T, Buléon A, Colon P, Jaillais B, Ceballos H, Dufour D 2012. Structural characterization of novel cassava starches with low and high-amylase contents in comparison with other commercial sources. Food Hydrocoll. 27:161-174. Crossref |
||||
| SAS Institute Inc. (2002). SAS/STAT software system for Windows: Changes and Enhancement for Release 9.1. Cary, NC: SAS Institute Inc. P. 158. | ||||
|
Sánchez T, Salcedo E, Ceballos H., Dufour D, Mafla G, Morante N, Calle F, Pérez JC, Debouck D, Jaramillo G, Moreno IX (2009). Screening of Starch Quality Traits in Cassava (Manihot esculenta Crantz). Starch 61:12–19. Crossref |
||||
|
Sayre R, Beeching JR, Cahoon EB, Egesi C, Fauquet C, Fellman J, Fregene M, Gruissem W, Mallowa S, Manary M, Maziya-Dixon B, Mbanaso A, Schachtman DP, Siritunga, D, Taylor N, Vanderschuren H, Zhang P (2011). The BioCassava Plus Program: Biofortification of Cassava for Sub-Saharan Africa. Annu. Rev. Plant Biol. 62:251–72. Crossref |
||||
|
Searle PL (1984). The Berthelot or indophenol reaction and its use in the analysis chemistry of nitrogen. The Analyst 109:549-565. Crossref |
||||
| Skalar Inc. USA (1995). The SANplus segmented flow analyzer. Soil and Plant Analysis. Skalar Analytical B.V., De Breda, The Netherlands, pp. 70-72. | ||||
| Steel RGD, Torrie JH (1960). Principles and Procedures of Statistics. McGraw-Hill, New York, pp. 39-40. | ||||
| Tanksley SD, McCouch SR (1997). Seed banks and molecular maps: Unlocking genetic potential from the wild. Science 277:1063-1066. | ||||
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


