Full Length Research Paper

ABSTRACT
The scarlet eggplant (Solanum aethiopicum L.) is an African traditional vegetable grown for its edibles leaves and fruits, which can contribute to food security in Republic of Benin. However, very little information is available on the genetic variability of this vegetable for varietal improvement purposes. Sixty accessions of scarlet eggplants collected throughout 52 villages in Benin republic were characterized using 34 (10 qualitative and 24 quantitative) variables. The experiment was laid out using complete randomized block design with three replications on the experimental site of Massi. The Principal Component Analysis (PCA) analysis revealed 18 discriminating quantitative variables. A strong correlation was observed between plant height, number of fruits per plant, fruit diameter, fruit weight and number of seeds per fruit. A cluster phenogram divided the 60 accessions into three groups corresponding to Kumba (Cluster 1), Gilo (Cluster 2), and Shum groups (Cluster 3). These groups differed by the number of seeds per fruit, weight of fruits, diameter of fruits, number of fruits per plant, plants height at flowering stage, plants length, and plant width. Accessions Samibi, Ikangougou, Kannan and Yètchanmiyé with the highest fruit weight per plant can be used as parents in the framework of a scarlet eggplant breeding program in Republic of Benin.
Key words: Benin, accessions, traits diversity, breeding program.
INTRODUCTION
The scarlet eggplant or garden eggplant (Solanum aethiopicum L.) is one of the most cultivated and consumed fruit vegetables in tropical Africa for its very diverse nutritional and medicinal values ??(Kouassi et al., 2014; Sabatino et al., 2019). Indeed, the various phytochemical compounds (alkaloids, saponins, flavonoids, tannins, ascorbic acid, and steroids) in the fruit and leaves of this traditional vegetable suggest its nutritionally and therapeutically value (Chinedu et al., 2011; Tunwagun et al., 2020). In Republic of Benin, scarlet eggplant is cultivated in all agro?ecological zones and its fruits and leaves are consumed as vegetable, raw or cooked in sauce (Achigan?Dako et al., 2010). Scarlet eggplant is known for its great morphological variability. Three cultivated groups (Gilo, Shum, and Kumba) having distinct morphological traits were found in some West-African countries such as Nigeria (Adeniji et al., 2019), Burkina Faso (Bationo-Kando et al., 2015; Sawadogo et al., 2016), Côte d'Ivoire (Kouassi et al., 2014), and Ghana (Osei et al., 2010). However, except for some studies mentioning the presence of Gilo and Shum groups in the Beninese agriculture (Achigan?Dako et al., 2010), very little information exist on the diversity of scarlet eggplant throughout the country.
In Republic of Benin, scarlet eggplant production is based on local varieties subjected to various biotic and abiotic constraints, which lead to their low yield. This situation is exacerbated by the fact that research and breeding programs are focused on other Solanaceae economically important. Furthermore, few studies have been done to improve the productivity of this traditional vegetable. A significant risk of extinction of scarlet eggplant local varieties exists in rural areas due to the introduction of exotic commercial eggplant such as brinjal eggplant (S. elongena L.). It is known that scarlet eggplant is an important source of traits of interest (resistance to nematodes, bacterial wilt, and fungi) to improve brinjal eggplant (Hébert, 1985; Rizza et al., 2002). Therefore, it is particularly important to characterize the accessions available and highlight the presence or expression of the criteria of interest before any selection or improvement program. Moreover, the analysis of agromorphological diversity within a species or between several species constitutes a very important asset in the identification of efficient varieties capable of alleviating the problems of producers and consumers (Rodriguez et al., 2008; Orobiyi et al., 2013, Kyriacou et al., 2017). For an effective breeding program, information concerning the extent and nature of genetic diversity within a crop species is essential. In fact, assessing the diversity and relationships of the cultivated species facilitates the establishment of conservation strategies, the use of genetic resources in breeding programmes, and the study of the crop evolution (Hurtado et al., 2012). Controlling agronomic diversity also involves eliminating duplicates, thereby facilitating precise individual knowledge of cultivars and their registration in the national catalogue, making them formal (Chapman, 2020).
This study aims at contributing to a better understanding of the agromorphological diversity of scarlet eggplant in the Republic of Benin. Specifically, it involves: (i) describing the qualitative diversity of the accessions collected using the parameters discriminating the African eggplant for the prospects of varietal improvement; (ii) establishing the level of quantitative structuring of the agronomic diversity of scarlet eggplant from Benin.
MATERIALS AND METHODS
Plant material
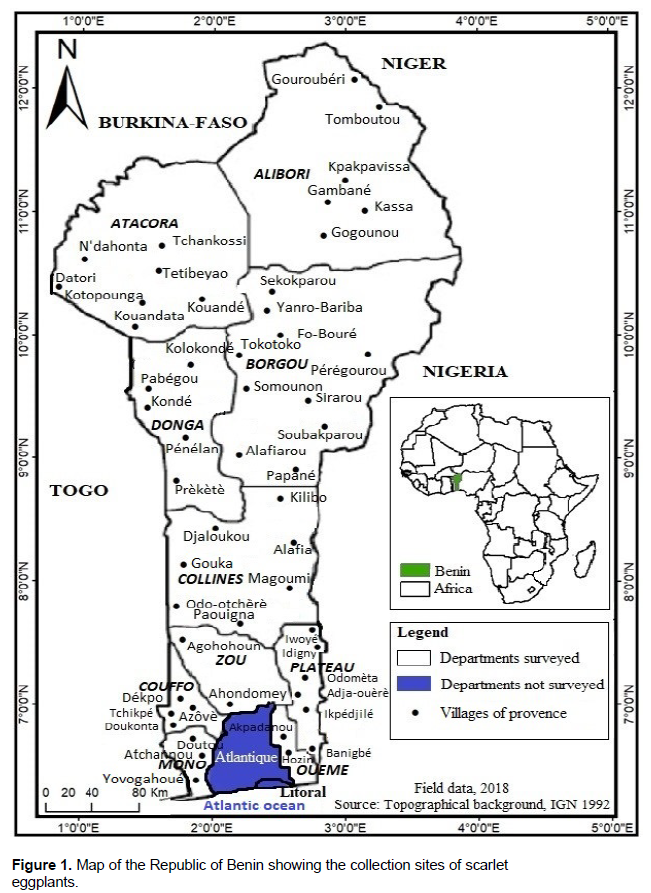
This study's plant material consisted of seeds obtained from 60 accessions of scarlet eggplants knowing under 22 different local names (Table 1). These accessions were collected in farmers’ fields throughout 52 villages (Figure 1).
Study site
Experiments were carried out on the experimental site of the BIORAVE Laboratory located in Massi (6° 56' and 7° 08' north latitude, 1° 58' and 2° 24' east longitude) in southern Benin. The cimate in this region is subequatorial with two rainy and two dry seasons.Rainfall varied between 1000 and 1400 mm. The relief is characterized by vast valleys of the Zou and Ouémé rivers, areas of low altitude plateau and ??the Lama depression area. Several types of soil were found in southern Benin whose vertisols at the experimental site. The vegetation consists of savannahs composed of several strata dominated by Daniella laxiflora and Parkia biglobosa (Adomou et al., 2007).
Growing conditions
The seeds were extracted from fruits, dried for 7 days and sowed in nursery for 45 days using the protocol described by Sawadogo et al. (2016). Hundred normal seeds of each accession were sowed in separated nursery bags loaded with ordinary sand and labelled. Transplanting of the seedlings was carried out at the 5 to 6 leaf stage following the recommendations of Segnou et al. (2012) adopted by Sawadogo et al. (2016). The randomized complete block design with 3 replications was used with regard to the high number of accessions and to control the high level of land heterogeneity conditions in Massi. In each repetition, each accession was planted in a row (elementary plot) of 7 plants. A distance of 0.8 m was respected between elementary plots while 0.4 m was applied between consecutive plants of the same row. To ensure good emergence of the transferred seedlings, regular and daily watering was done for 10 days after transplanting. No organic manure was added. Weeding was done manually without the use of chemicals. In addition, the treatments applied against scarlet eggplant diseases were made with the pesticide namely Dominator Plus at a dose of 15 ml / 10 m2 (15 L / ha) and this every two weeks as recommended by Simeni et al. (2009).
Characterization
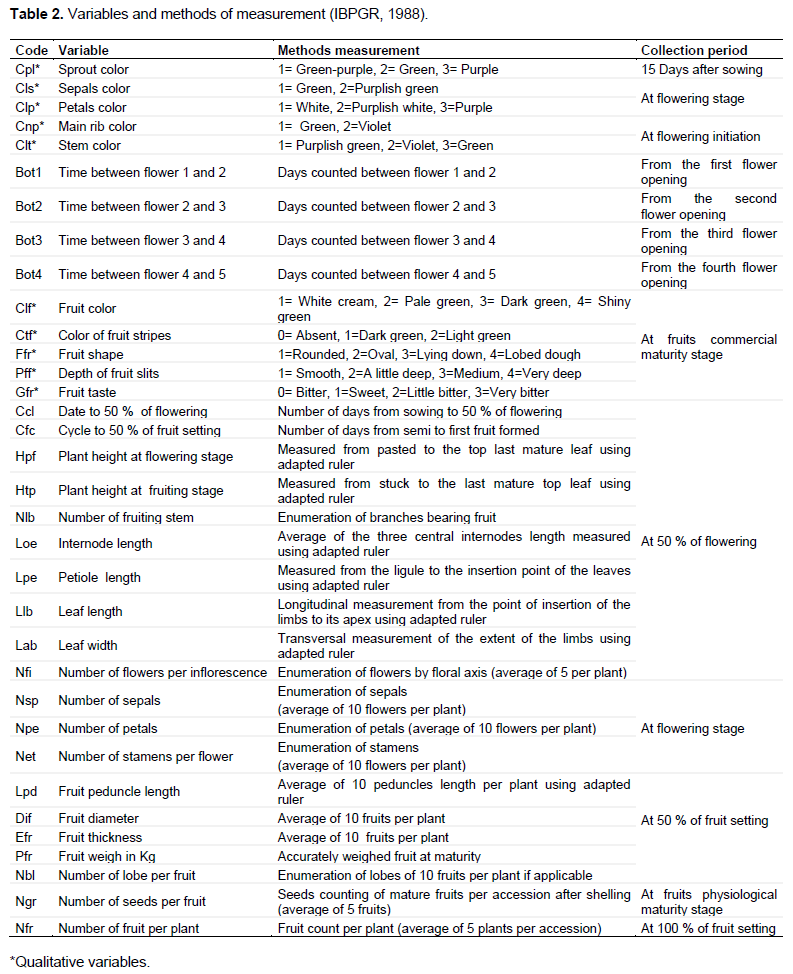
Thirty-four morpho-agronomic traits (10 qualitative and 24 quantitative) from eggplant descriptors developed by the International Board for Plant Genetic Resources (IBPGR, 1990) and revealed by Simeni et al. (2009), Danquah and Ofori (2012) as more discriminating were used. These data were collected on five marked plants in each elementary plot and in the three blocks. The sowing-to-flowering cycle and the color of the seedlings at emergence were noted across the row. The qualitative parameters were evaluated during direct observation sessions on the experimental plots and tasting tests carried out by a team of experienced consumers. The observations were noted as detailed scores in Table 2.
Data analysis
Two databases (categorical and quantitative) were developed through observations and measurements made during the plant development cycle. The frequencies of appearance of the modalities of each qualitative trait were estimated using the formula described by Sawadogo et al. (2016) as followed:
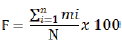
With F the frequency (%), mi indicates number of accessions with the same modalities and N indicates the total number of accessions evaluated. The results were summarized in descriptive table and graphics. As for the quantitative variables, a Principal Component Analysis (PCA) was firstly done to identify the most discriminating traits according to Peres-Neto et al. (2003). The correlation analysis was after that performed as heatmap considering the most discriminant variables using packages ggplot2 (Wickham et al., 2016). The PCA was summarized into a biplot showing accessions with corresponding discriminant variables using FactoMineR (Husson et al., 2016) and factoextra (Kassambara and Mundt, 2017a, b) packages. To classify the studied accessions according to those quantitative variables, a Multiple Correspondence Analysis (MCA) was then carried out and the representative factors was used to perform the Hierarchical Cluster Analysis (HCA) using factoextra (Kassambara, 2017a, b; Coghlan, 2017) and cluster packages (Bates et al., 2018) with R software version 4.0.2 (R Core Team, 2018). Pheatmap (Kolde and Kolde, 2015) and ggplot2 (Wickham et al., 2016) packages were used to plot dendrogram.
RESULTS
Morphological diversity of scarlet eggplants
Two main stem colours (green and purple) were observed during the plants sprouting with an absolute predominance of the green colour (86.43 % of accessions). However, 3.24% of the accessions displayed a mixture of green and purple colour (Figure 2). The great majority of accessions (71.66%) were characterized by leaves with green veins against 28.33% purple both at the seedling stage and at physiological maturity.The floral pieces, in particular the petals, presented three distinctive colour (white, purplish white, purple) with a predominance of flowers with purplish white petals (58.33%), 26.67% of the accessions presented inflorescences with purple petals, while only 15% presented white petals (Figure 2). The sepals have a more or less uniform light green coloration (80%). Only 20% of the accessions are distinguished from the others by the sepals purplish green colouring. At fruiting stage (at least 2 month after planting), accessions were mainly identified by the colour and shape of the fruits.

In terms of fruits colour, four different modalities were observed (Table 3). The studied germplasm was dominated by accessions with pale green fruits (36.67%), followed respectively by those with dark green (26.67%), shiny green (20%), and white cream fruits (16.67%). Four types of accessions were identified concerning fruits taste (Table 3). The germplasm was dominated by accessions with bitter fruits (51.67%), while those with sweet (26.67%), slightly bitter (16.67%), and very bitter (5%) fruits were little represented.
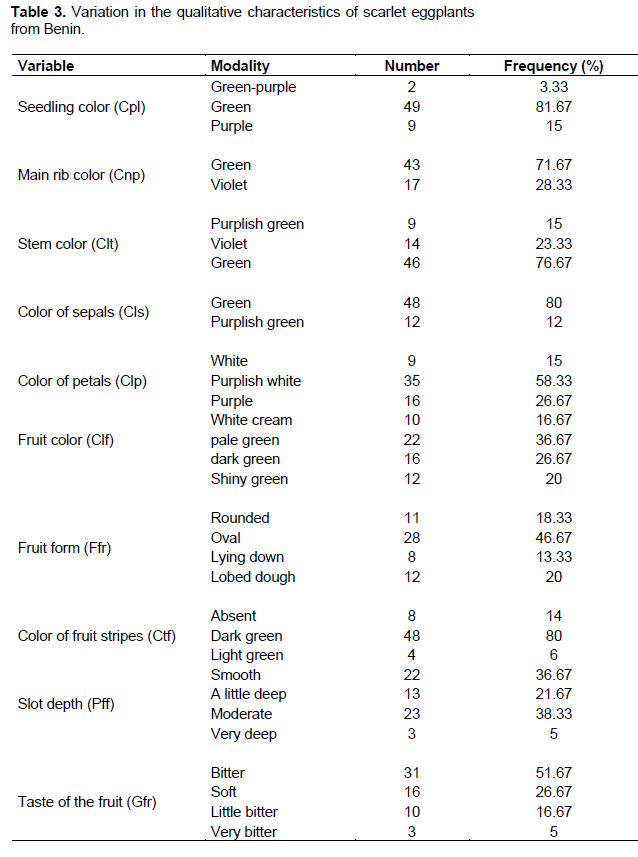
Stripes of different colours on the fruit are also part of the eggplant's distinctive quality traits (Table 3). Thus, the collection studied in Benin presents for most of the accessions an overlap of dark green stripes with a dominant proportion (80%) on the fruits. Fruit accessions with light green stripes are around 6% of the current Beninese collection. However, 14% of this collection were without remarkable stripes and are either entirely white or pale green. The description of fruits shape revealed three different modalities clearly dominated by the oval fruits (46.67%). Accessions with lobed and non-lobed flat fruits represent 20 and 13.33% while rounded fruit accessions are 18.33%. Within the lobed fruit accessions, there is also a variability in the depth of the side slits of the fruit delimiting the lobes. A significant proportion (36.67%) of the accessions have fruits without slits (smooth fruits). Accessions with shallow and moderately deep lobed fruits represent respectively 21.67 and 38.33%. Only 5% of accessions have fruits with very deep lobes.
In summary three morphological groups are observed (Gilo, Kumba and Shum) (Figure 3). The Gilo groups are characterized by smooth to slightly lobed oval fruits of shiny green, white and striped green (dark green longitudinal stripes; Figure 3a). However, the Kumba group's accessions have strongly lobed and flat fruits with dark green, white to glossy green colourings, devoid of stripes (Figure 3b). As for Shum, the fruits are generally small, rounded and green with dark stripes. The latter are the most bitter (Figure 3c).

Phenotypic scarlet eggplant diversity using quantitative variables
Eighteen discriminant variables were correlated to the first five principal components identified as most discriminating the germplasm collection (Table 4). Those components explained 76.86% of the total quantitative variation in the studied collection. Thirteen variables (flowering cycle (Ccl), fruiting cycle (Cfc); internodes length (LoE); leaf length (Llb); width of leaf (Lab); number of sepals (Nsp); number of petals ([Npe); number of stamens (Net); number of fruits (Nfr); diameter of the fruit (Dif); weight of the fruit (Pfr); number of seeds per fruit (Ngr) and number of flower per inflorescence (Nfi)) were significantly correlated with the first components accounted for 35.76% of the total variation. The second principal component explained 15.81% of the total variation and associated with the number of fruiting stem (Nbf) and Fruits thickness (Efr). The third factor accounted for 10.38% of the total variation and was mainly correlated with number of seeds per fruit (Table 4). The fourth component is linked to the plant height at flowering (Hpf) stage, while the last component (PCA 5) is only associated with fruits weight (Pfr) (Table 4).
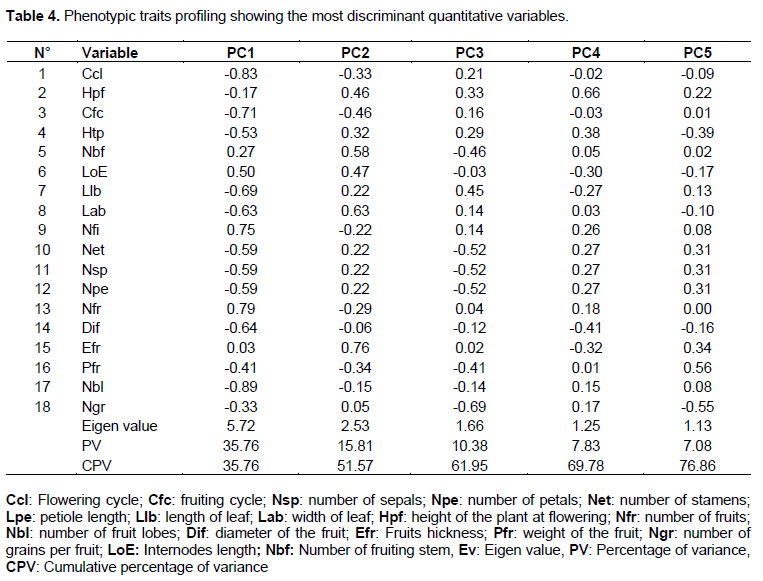
Variation between the most discriminatory quantitative variables
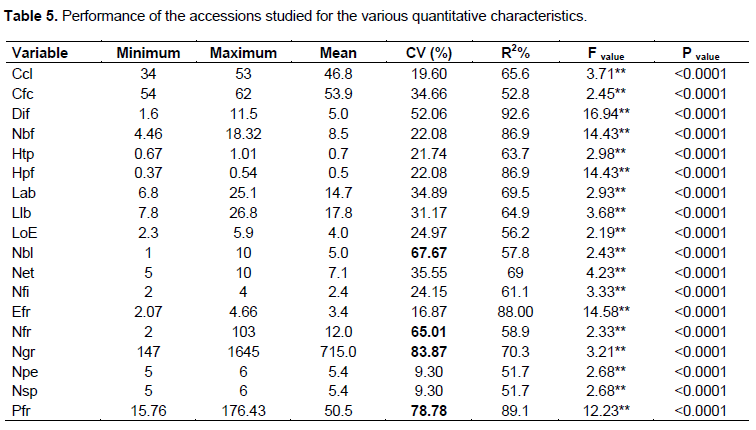
All discriminant variables are highly determinate (R2 ≥ 30%). The large differences observed between the statistical parameters (minimum and maximum) and the large values ??of coefficients of variation explain the great heterogeneity existing within Benins accessions of scarlet eggplants. The most significant variations were observed for the fruiting criteria, in particular the number of fruits per plant, the weight of the fruits, the number of seeds per fruit and the number of fruit lobes while the smallest variations are recorded for the floral parts (petals, sepals and stamens number). The flowering cycles of the studied accessions varied from 34 to 53 days after transplanting with the Gbitchangodo (A2), Gboyigouroto (A12) and Koklozingbo (A23) accessions having the early flowering cycle and Sambini (Djougou, A57) and Gabta (A48) accessions the later cycle. It is very important to point out that flowering occurs when the plants have an average height of 0.37 m and fruiting when they are 0.67 m in height. The leaves length generally varies between 7.8 and 26.8 cm while their width varies from 6.8 to 25.1 cm. Accessions Yekan (A28), Yekantchan’tchayé (A42), Kpaanoulaka (A24) and Gabta (Cotiakou, A56) presented the longer and wider leaves. A minimum value of 1.6 cm was recorded for the diameter of the fruit, while the maximum value was 11.5 cm. The net weight of the fruits varied from 15.76 to 176.43 g with an average number of 715 seeds per fruit (Table 5). Accessions Samibi (N’dali,
A19), Ikangougou (A53), Kannan (A34) and Yètchanmiyé (A27) presented the higher fruit weight and number of seed per fruit.
Phenotypic correlations among the quantitative variables
Positive correlations were observed between the flowering cycle (Ccl), the fruiting cycle (Cfc), the length of the blade (Llb) and the height of the plant at flowering stage (Hpf) (Figure 4). An important positive link exists between the floral parts (sepal, petal and stamen) and some fruits characteristics including the number of lobes (Nbl), seeds, weight (Pfr) and fruits diameter (Dif) (Figure 5).
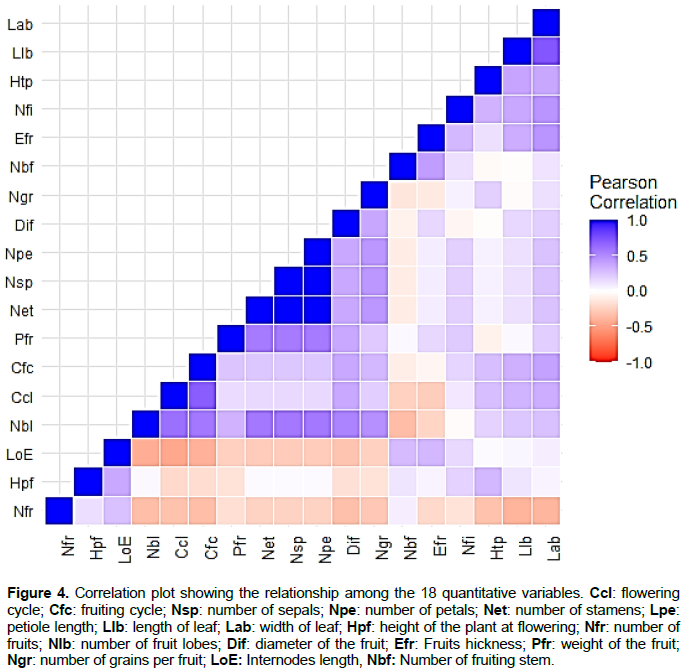
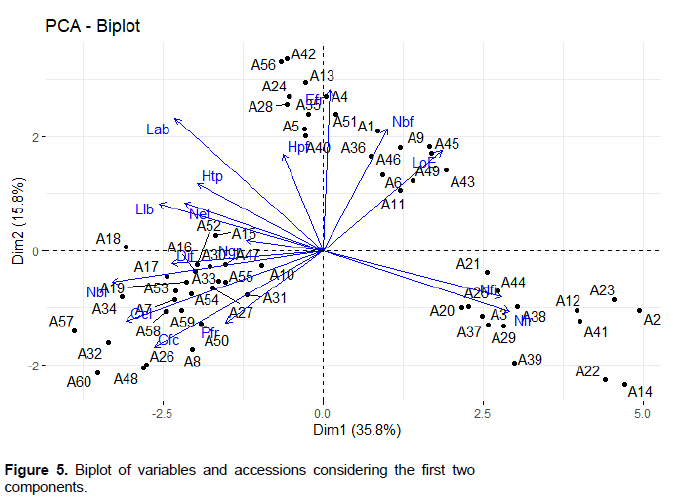
The number of fruits per plant (Nfr) was positively linked to the number of fruiting branches (Nbf). There is also a slight correlation between the leaves length (Llb) and width (Lab) as well as between the number of fruit lobes (Nbl) and the number of seeds (Ngr) and the fruits weight (Pfr). In addition, a very high correlation was observed between flowers parameters including the number of sepal (Nsp), petal (Npe) and stamen (Net) per flower (Figure 4).
Classification of scarlet eggplant accessions
The projection of variables and individuals in the principal component analysis plan revealed heterogeneity within the accessions studied (Figure 5). However, a more or less absolute similarity can be observed between the accessions around three groups of variables (Figure 5). Thus, the number of fruits per plant (Nfr) and the number
of flowers per inflorescence (Nfi), positively associated with axis 1 (Dim1) seem to be characteristic of a given accession group. Likewise, this deduction is valid for the group of variables positively correlated with this same axis and associated with axis 2 (Dim2).
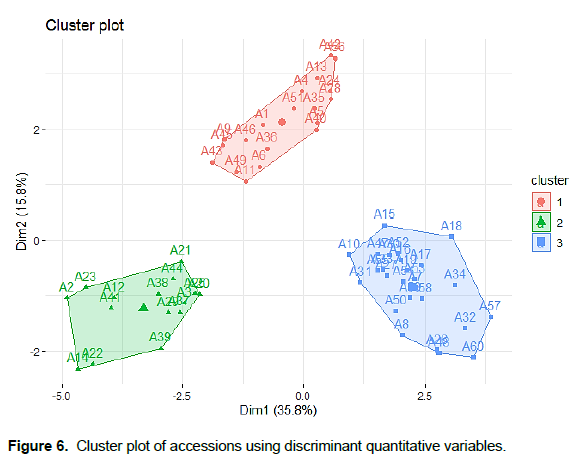
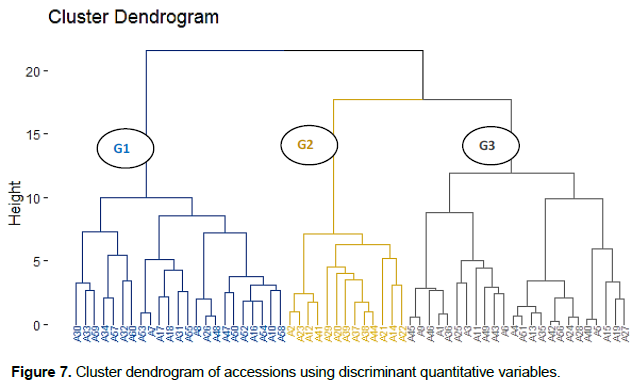
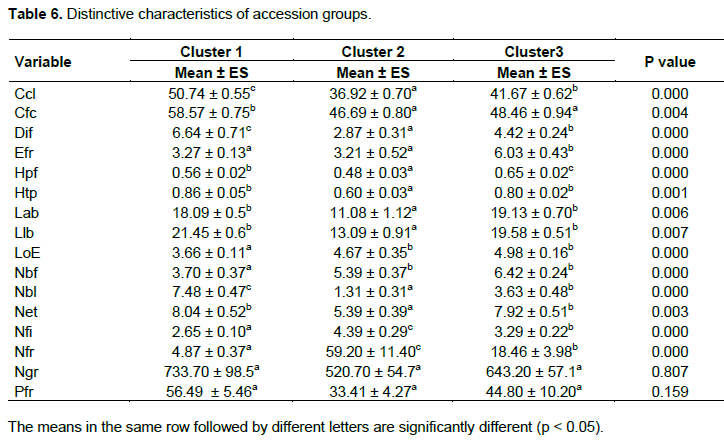
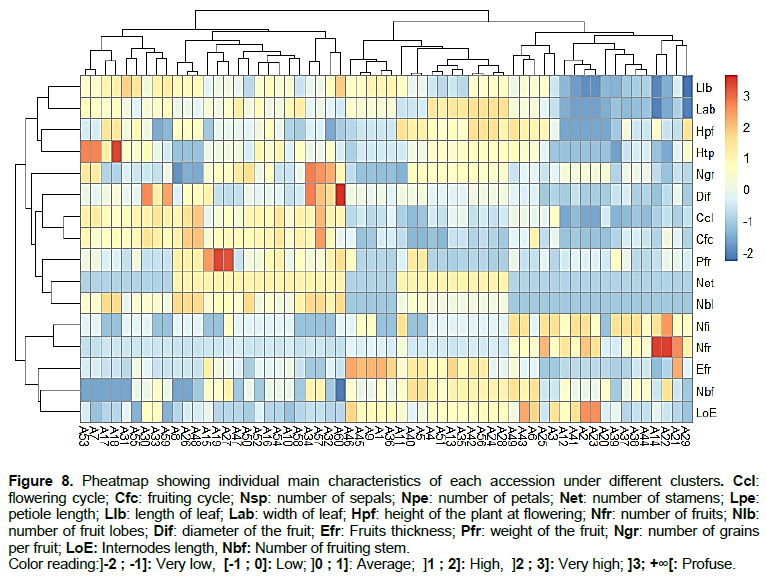
From the cluster analysis based on the similarity using the discriminatory phenotypic variables, the 60 accessions evaluated were grouped into 3 clusters (Figure 6). The first cluster was made up of 23 accessions while the second and the third were contained 13 and 24 accessions (Figures 6 and 7). The three clusters of scarlet accessions obtained were significantly different in terms of flowering cycle, height of the plant at flowering, number of flowers per inflorescence, number of fruit lobes, number of fruits, and fruit diameter (Table 6). The accessions grouped in the first cluster were characterised by their late flowering, late fruit setting, large fruits, high number of fruit lobes, short internodes length, low number of fruiting stem, low number of flowers per inflorescence, and low number of fruits per plant. Accessions characterised the second cluster with early flowering, small fruit, low height of the plant at flowering and fruiting stage, short leaves, low number of stamens per flower, high number of flowers per inflorescence, and high number of fruits per plant. The third cluster grouped accessions with thick fruits and high height of the plant at flowering stage compared to the other cluster. However, there is also a slight variability between different accessions of the same cluster considering the most 17 discriminant variables (Figure 8).
DISCUSSION
The diversity of cultivated plant based on morphological and agronomic parameters reveals diversity as local farmers perceive (Sawadogo et al., 2016; Djaha et al., 2017; Gmakouba et al., 2018). It is the most practical approach for phenotypic eggplant differentiation when combined with a statistical tool, which estimates the effects linked to production conditions (Kouassi et al., 2014; Sabatino et al., 2019). This study qualitative descriptors revealed significant morphological diversity within the eggplant accessions evaluated in Benin Republic. Several authors have also reported a high morphological diversity within scarlet eggplant in several West Africa countries (Osei et al., 2010; Bationo-Kandoet al., 2015; Sawadogo et al., 2016). This high diversity is possibly caused by allogamy rating more than 65% in scarlet eggplant accorging to Sawadogo et al. (2016). In addition, most of the morphological characters are under the control of genes with additive effects. This statement seems to be suitable for explaining the phenotypic variabilities observed with regard to categorical phenotypic traits in Beninese scarlet eggplants.
As for the quantitative criteria, the significant differences between the means of each discriminant variable and the high coefficients of variation indicate a high heterogeneity within the evaluated accessions. Several studies have also revealed a great diversity between scarlet eggplant accessions with regard to quantitative characters (Sawadogo et al., 2016). This diversity is much more determined by the fruits criteria such as the number of fruits per plant, the diameter, the weight of the fruits and the number of seeds per fruit. Several authors have reported similar results (Osei et al., 2010; Kouassi et al., 2014; Plazas et al., 2016; Wahua and Olaleye, 2017; Sseremba et al., 2017). However, in Burkina Faso, the number of flowers per inflorescence was mentioned as the most discriminating variable (Sawadogo et al., 2016). Accessions Samibi (N’dali, A19), Ikangougou (A53), Kannan (A34) and Yètchanmiyé (A27), which presented the high fruit weight could be used as source of genes in a breeding program for creation of high yielding varieties. Similarly, the significant positive correlations between scarlet eggplant yield parameters must be considered during improvement through selection (Plazas et al., 2016).
Being approved that there is a great interdependence between different stages of plant development (Anjarwalla et al., 2016), it is reasonable to observe strong correlations between the measured quantitative variables (Danquah and Ofori, 2012). The yield in photosynthetic reserve depends not only on the available vegetative mass (Adeniji et al., 2019) but also on geophysical conditions (Chiemela and Ricardo, 2015). This assertion was confirmed on the accessions of Kumba (bitter eggplant) from Burkina Faso (Sawadogo et al., 2016). According to Chiemela and Ricardo (2015) there are very often important interactions notably between environments and plant phenotypes. However, the principal component analysis and the hierarchical clustering showed a random distribution of accessions throughout the study area.
The three eggplant groups (Gilo, Kumba, and Shum) revealed in our collection approves the existing diversity in West Africa (Fondioet al., 2015; Wahua and Olaleye, 2017). It is known that here is a highly significant difference between scarlet eggplant groups (Bationo-Kandoet al., 2015; Wahua and Olaleye, 2017). In Benin, the Gilo produces more fruits of medium size and weight than the Kumba, whose fruits are large with a high weight. Similarly, within the Shum group's accessions, the fruits are very small compared to the Gilo and the Kumba. The Shum group has the bitterest fruits and the seed content is associated with this bitter (San José et al., 2016). This is also due to the fact that the mucilage surrounding the seeds contains high level of saponins and alkaliods (Chinedu et al. 2011; Ayodele, 2018). These observations confirm the results of recent studies conducted in Burkina Faso (Sawadogo et al., 2016), and Tanzania (Adeniji et al., 2019).
CONCLUSION
Our results showed that scarlet eggplant from the Republic of Benin offer enormous potential for breeding programs through significant phenotypic variation revealed within the 60 accessions. Scarlet eggplant accessions was classified in three groups corresponding to Gilo (sweet), Kumba (bitter) and Shum (very bitter) groups. Yekan and Yekantchan’tchayé accessions with their long and broad leaves presented valuable potential for future scarlet breeding program as vegetables. While, Samibi, Ikangougou, Kannan and Yètchanmiyé accessions could be promoted to boost the production of scarlet eggplant fruits. The various information obtained on the morphological diversity of scarlet eggplants cultivated in the Republic of Benin will be very useful for the conservation and improvement of this vegetable.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Achigan?Dako EG, Pasquini MW, Assogba?Komlan F, N'danikou S, Yédomonhan H, Dansi A, Ambrose?Oji B (2010). Traditional vegetables in Benin. Institut National des Recherches Agricoles du Bénin. Imprimeries du CENAP, Cotonou P 285. |
|
|
Adeniji OT, Kusolwa PM, Reuben SWOM (2019). Quantitative analysis of fruit size and fruit number in Solanum aethiopicum group. Analysis of fruit in Solanum aethiopicum. Agrieast: Journal Agricultural Science 13:27-37. |
|
|
Adomou AC, Akoègninou A, Sinsin B, de Foucault B, van der Maesen LJ (2007). Notulae Florae Beninensis, 13-Biogeographical analysis of the vegetation in Benin. Acta Botanica Gallica 154:221-233. |
|
|
Anjarwalla P, Belmain S, Sola P, Jamnadass R, Stevenson PC (2016). Handbook on Pesticidal Plants.World Agroforestry Centre (ICRAF), Nairobi, Kenya P 64. |
|
|
Ayodele OD (2018). Mini review on two species of garden egg (S. aethiopicum L. and S. macrocarpon L.) found in Nigeria. Journal of Animal Research 7(2):237-238. |
|
|
Bates DM, Maechler M, Bolker BM, Walker S, Christensen RHB, Singmann H, Green P (2018). Package "lme4." Retrieved April 3, 2020. |
|
|
Bationo-Kando P, Sawadogo B, Nanéma KR, Kiébré Z, Sawadogo N, Kiébré M, Traoré RE, Sawadogo M, Zongo JD (2015). Characterization of Solanum aethiopicum (Kumba group) in Burkina Faso. International Journal of Security and Networks 6(2):169-176. |
|
|
Chapman MA (2020). Eggplant breeding and improvement for future climates. In Genomic Designing of Climate-Smart Vegetable Crops. Springer, Cham. pp. 257-276. |
|
|
Chiemela FA, Ricardo LI (2015). Performance and adaptability of two yam (Dioscerea spp) varieties under ifugao condition. International Journal of Advanced Research 3:110-116. |
|
|
Chinedu SN, Olasumbo CA, Eboji KO, Emiloju OC, Arinola OK, Dania I (2011). Proximate and phytochemical analyses of Solanum aethiopicum L. and Solanum macrocarpon L. fruits. Research Journal of Chemical Sciences 1(3):63-71. |
|
|
Coghlan A (2017). A little book for R for multivariate analysis. Wellcome Sanger Institute. |
|
|
Danquah JA, Ofori K (2012). Variation and correlation among agronomic traits in 10 accesssions of garden eggplant (Solanum gilo Raddi) in Ghana. International Journal of Nature and Science 3(2):373-379. |
|
|
Djaha KE, Abo K, Bonny BS, Kone T, Amouakon WJL, Kone D, Kone M (2017). Caractérisation agromorphologique de 44 accessions de manioc (Manihot esculenta Crantz) cultivés en Côte d'Ivoire. International Journal of Biological and Chemical Sciences 11(1):174-184. |
|
|
Fondio L, N'tamon LN, Hala FN, Djidji HA (2015). Evaluation agronomique de six cultivars d'aubergine africaine (Solanumspp.) de la nouvelle collection des plantes légumières du CNRA. African Journal of Agronomy 20(1):69-79. doi: 10.4314/aga.v20i1.1737. |
|
|
Gmakouba T, Koussao S, Traore ER, Kpemoua KE, Zongo JD (2018). Analyse de la diversité agromorphologique d'une collection de manioc (Manihot esculenta Crantz) du Burkina Faso. International Journal of Biological and Chemical Sciences 12(1):402-421. |
|
|
Hébert Y (1985). Comparative resistance of nine species of the genes Solanum to bacterial wilt Psedomonas solanacearum and the nematode Meloidogyne incognita. Implications for the breeding of aubergine (S. melongena) in the humid tropical zone. Agronomie 5(1):27-32. |
|
|
Hurtado M, Vilanova S, Plazas M, Fonseka HH, Fonseka R, Prohens J (2012). Diversity and relationships of eggplants from three geographically distant secondary centers of diversity. Plos ONE 7(7):41748. |
|
|
Husson F, Josse J, Le S, Mazet J, Husson MF (2016). Package 'FactoMineR'. An R package 96:698. |
|
|
IBPGR (1990). Descriptors for eggplant/descripteurs pour aubergine. International Board for Plant Genetic Resources, Rome P 46 |
|
|
Kassambara A (2017a). Practical guide to cluster analysis in R: Unsupervised machine learning. Sthda, Vol. 1. |
|
|
Kassambara A (2017b). Practical guide to principal component methods in R: PCA, M (CA), FAMD, MFA, HCPC, factoextra. Sthda, Vol. 2. |
|
|
Kassambara A, Mundt F (2017a). Factoextra: extract and visualize the results of multivariate data analyses. R package version 1.0.4.999. |
|
|
Kassambara A, Mundt F (2017b). Package 'factoextra'. Extract and visualize the results of multivariate data analyses. R Package Version 1(5):337-354. |
|
|
Kolde R, Kolde MR (2015). Package 'Pheatmap'. R Package 1(7):790. |
|
|
Kouassi A, Béli-Sika E, Tian-Bi TYN, Alla-N'Nan O, Kouassi AB, N'Zi JC, N'Guetta ASP, Tio-Touré B (2014). Identification de trois sous-groupes d'aubergines distincts au sein du groupe Solanum aethiopicum Gilo de Côte d'Ivoire par morpho-agronomiecaractérisation. Agriculture 4(4):1-14 |
|
|
Kyriacou MC, Rouphael Y, Colla G, Zrenner R, Schwarz D (2017). Vegetable grafting: The implications of a growing agronomic imperative for vegetable fruit quality and nutritive value. Frontiers in Plant Science 8:741. |
|
|
Orobiyi A, Dansi A, Assogba P, Loko LY, Dansi M, Vodouhè R, Akouègninou A, Sanni A (2013). Chile (Capsicum annuum L.) in southern Benin: production constraints, varietal diversity, criteria and participatory evaluation. International Research Journal of Advanced Science 3(4):107-120. |
|
|
Osei MK, Banful B, Osei CK, Oluoch MO (2010). Characterization of African eggplant for morphological characteristics Journal of Agriculture, Science and Technology 4(3):28-36. |
|
|
Peres-Neto PR, Jackson DA, Somers KM (2003). Giving meaningful interpretation to ordination axes: assessing loading significance in principal component analysis. Ecology 84(9):2347-2363. |
|
|
Plazas M, Kaushik P, Prohens J, Vilanova S, Gramazio P (2016). Phenotyping of eggplant wild relatives andinterspecific hybrids with conventional and phenomics descriptors provides insight for their potentialutilization in breeding. Frontiers in Plant Science 7:677. |
|
|
R Core Team R (2018). A Language and Environment for Statistical Computing, R Foundation for Statistical Computing: Vienna, Austria, 2018, |
|
|
Rizza F, Mennella G, Collonnier C, Shiachakr D, Kashyap V, Rajam MV, Prestera M, Rotino, GL (2002). Androgenic dihaploids from somatic hybrids between Solanum melongena and S. aethiopicum group gilo as a source of resistance to Fusarium oxysporum f. sp. melongenae. Plant Cell Report 20(11):1022-1032. |
|
|
Rodriguez E, Forster MP, Diaz Romero C (2008). Differential characteristics in the chemical composition of bananas from Tenerife (Canary Islands) and Ecuador.Journal of Agricultural and Food Chemistry 50(25):7586-7592. |
|
|
Sabatino L, Iapichino G, Rotino GL, Palazzolo E, Mennella G, D'Anna F (2019). Solanum aethiopicum gr. gilo and Its Interspecific Hybrid with S. melongena as Alternative Rootstocks for Eggplant: Effects on Vigor, Yield, and Fruit Physicochemical Properties of Cultivar ′Scarlatti′. Agronomy 9:223. |
|
|
San José R, Plazas Ávila MDLO, Sánchez-Mata MC, Cámara Hurtado MM, ProhensTomás J (2016). Diversity in composition of scarlet (S. aethiopicum) and gboma (S. macrocarpon) eggplants and of interspecific hybrids between S. aethiopicum and common eggplant (S. melongena). Journal of Food Composition and Analysis 45:130-140. |
|
|
Sawadogo B, Bationo-Kando P, Sawadogo N, Kiebre Z, Kiebre M, Nanema KR, Traore RE, Sawadogo M, Zongo JD (2016). Variation, correlation and heritability of interest characters for selection of African eggplant. African Crop Science Journal 24(2):213-221. |
|
|
Segnou J, Akoa A,Youmbi E (2012). Viabilité et développementvégétatif des plantules de piment (Capsicum annuum L.) suivant différents matériels de conditionnement des semences. Tropicultura 30(1):15-23. |
|
|
Simeni GT, Adeoti R, Abiassi E, Kodjo MK, Coulibaly O (2009). Caractérisation des systèmes de cultures maraîchères des zones urbaine et périurbaine dans la ville de Djougou au Nord-Ouest du Bénin. Bulletin de la Recherche Agronomique du Bénin 64:34-49. |
|
|
Sseremba G, Tongoona P, Eleblu JSY, Danquah EY, Kabod NP, Kizito EB (2017). Morphological distinctiveness between Solanum aethiopicum Shum group and its progenitor. Journal of Plant Breeding and Crop Science 9(8):118-129. |
|
|
Tunwagun AD, Akinyemi OA, Ayoade TE, Oyelere SY (2020). Qualitative analysis of the phytochemical contents of different anatomical parts of ripe Solanum aethiopicum Linneaus Fruits. Journal of Complementary and Alternative Medical Research 9(3):14-22. |
|
|
Wahua C, Olaleye MS (2017). Comparative Taxonomic Studies on Solanum aethiopicum Linn. and Solanum nigrum Linn. (Solanaceae), Greener Journal of Agricultural Sciences 3(12):849-854. |
|
|
Wickham H, Chang W, Wickham MH (2016). Package 'ggplot2'. Create Elegant Data Visualisations Using the Grammar of Graphics. R package version 2(1):1-189. |
|
|
IBPGR (1988). Descriptors for eggplants. International Board for Plant Genetic Resources, Rome, Italia 23 p. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


