Full Length Research Paper

ABSTRACT
Twenty one F1s produced from 7x7 diallel mating along with the 7 parents were evaluated to notice the inheritance and combining ability of different traits to obtain high heterotic crosses. Genetic analysis and combing ability were analyzed following Hayman’s and Griffing’s diallel analyses, respectively. Hybrids projecting positive or negative potency ratio with >1.0 value for those traits is also the sign of incidence of over-dominance in desirable direction, and heterosis breeding is important to improve those traits in maize. Hayman’s approach indicated dominance variance and the proportion of +/- genes was higher than additive variance in all characters. Griffing’s analysis also demonstrated the presence of over-dominance governing the traits. The preponderance of dominant gene action coupled with low heritability observed for days to silking, ear length and grain yield suggests the importance of heterosis breeding. Substantial differences in general combing ability and specific combining ability were noticed in all the studied traits except 1000-grain weight. The parental line CML-509 was found to be the best general combiner for days to tasseling and silking, CML-498 for plant height, ear height and grain yield, CML-395 for ear length and grain yield. The crosses CML-498×CML-376, CML-498×CML-395 and CML-376×CML-247 showed significant positive specific combining ability effect for grain yield along with higher mean values over commercial check varieties.
Key words: Inheritance, combining ability, diallel mating, maize.
INTRODUCTION
Maize belongs to the grass family Poaceae. It is one of the most important cereal crops of the world. It is a major grain crop globally, which can be grown in comprehensive climatic conditions. Globally, maize is the third most important crop. It is a versatile crop grown over a wide range of agro-climatic zones. In fact, the suitability of maize to diverse environments is unmatched by any other crop. It is grown from below sea level to altitudes higher than 3000 m, and in areas with 250 mm to more than 5000 mm of rainfall per year; its growing cycle ranges from 3 to 10 months (Sheikh et al., 2017). According to FAO (2016), total area of maize cultivation was 188 million hectare (ha) with production of 1050.1 million ton and average yield of 5.64 ton ha-1. Globally, maize is popular for its multipurpose uses with utmost grain yield. It is used as human food, poultry, livestock and fish feed. Due to increasing poultry and fish feed industry, its demand is increasing continuously.
Keeping in mind the huge demand of maize as feed for poultry and livestock industry as well as food for human, new high yielding hybrid developing program has been going on. To achieve this target, yield improvement through genetic approaches that determine gene action is essential for formulating comprehensive breeding strategies. Yield improvement of any crop depends mostly on understanding the nature of gene action involved in a specific trait to be improved. In addition, the choice of competent breeding program depends on the large knowledge of the nature of gene action of yield related traits. Dominance gene action is desirable for developing hybrids while additive gene action effectively improves character (Edwards et al., 1976). One of the most helpful approaches in this concern is diallel analysis system extensively used in hereditary research to investigate the inheritance of important traits among a set of genotypes (Yan and Hunt, 2002). Components of genetic control and help breeders in the selection of desirable parents for crossing programs, and thus, facilitate in deciding a suitable breeding procedure for genetic improvement of various quantitative traits (Jinks and Hayman, 1963; Walters and Morton, 1978; Reza et al., 2004).
On the other hand, the ability of a line to transfer its performance to others is described as combining ability of inbred line. Combining ability of inbred lines provide information about genetic nature of quantitative traits as well as for selection of suitable parents to be used for heterosis breeding. General combining ability (GCA) is helpful for the improvement of selection efficiency in segregating populations (Bocanski et al., 2009). Specific combining ability (SCA) is specific performance of any two inbreeds in hybrid combination. Variance due to GCA is an indicator of the extent of additive gene action whereas variance due to SCA shows the extent of non-additive gene action (Hayman, 1954; Griffing, 1956).
The diallel cross technique was developed by Sprague and Tatum (1942). Hayman numerical approach (Haymen, 1954) provides information about inheritance pattern of particular character while Griffing (1956) provides a feature on gene action and combining ability of parental lines. The two main genetic parameters of diallel cross analysis are GCA and SCA. Since the GCA effects are endorsed by the preponderance of genes with additive effects and SCA indicates a predominance of genes with non-additive effects (Falconer, 1981), diallel crosses have been used for a long time in genetic research to determinate the inheritance of a trait among a set of genotypes and to identify superior parents for hybrid or cultivar development (Aliu et al., 2009). These methods have been extensively in different crops like maize (Njeri et al., 2017; Owusu et al., 2017; Brahmbhatt et al., 2018), rice (Huang et al., 2015; Kundan et al., 2013), Brassica (Tian et al., 2017) and cassava (Tumuhimbise et al., 2014). Therefore, it is necessary to understand the nature and magnitude of gene action as well as combining ability of yield and its attributes. The present investigation of 7×7 diallel cross I maize without reciprocal crosses was undertaken to supplement genetic parameters interpretations, pinpoint which parents contain the preponderance of dominance/recessive genes with increasing/decreasing character attributes, and isolate superior inbred lines and better combining parents for utilizing them judiciously in future breeding programs. Heterosis using commercial checks was also reported.
MATERIALS AND METHODS
Plant materials
Seven maize inbred lines (CML 498, CML 376, CML 247, CML 509, CML 502, CML 144 and CML 395) collected from International Maize and Wheat Improvement Center (CIMMYT ) were crossed in a diallel fashion excluding the reciprocals during the rabi season in 2014-15 at Bangladesh Agricultural Research Institute (BARI), Gazipur, Bangladesh. The resulting 21 F1’s and their 7 parents were evaluated along with two commercial checks (900M GOLD and NK40) in a randomized complete block design (RCBD) with three replications at the same location in the following rabi (winter) season of 2015- 2016.
Experiment settings, crop management and data recording
Seeds of each entry were sown in two rows of 4 m plot. The spacing between rows was 60 cm and plant to plant distance was 25 cm. Fertilizers were applied at 250, 55, 110, 40, 5 and 1.5 kg ha-1 of N, P, K, S, Zn, and B respectively. One plant per hill was maintained after proper thinning. Observations were recorded on five randomly selected competitive plants from each plot for days to 50% tasseling, days to 50% silking, plant height (cm), ear height (cm), ear length, number of grains/row, 1000-grain weight (g) and grain yield (t ha-1). Data for 50% days to tasseling and 50% silking as well as grain yield were recorded on whole plot basis and finally, grain yield converted to t ha-1. Data were analyzed for the variance for all the characters studied.
Statistical analysis
The mean performances of all characters were analyzed using Crop Stat software. Gene action was clarified by genetical analysis as proposed by Hayman’s numerical approach (1954a, b). According to him, the size of mean degree of dominance (H1/D).5 was categorized as (H1/D).5 = 0, mean no dominance, (H1/D).5 = 1, mean complete dominance, (H1/D).5 >1, mean over dominance and (H1/D).5 <1 mean partial dominance. The proportion of dominant and recessive alleles is ascertained by the ration (4DH1).5 + F/(4DH1).5 – F. Its value explain (4DH1).5 + F/(4DH1).5 – F @ 1.0 means nearly equal proportion of dominance and recessive alleles in parents that is symmetrical distribution; p=q=0.5. If its value is >1.0 refers to an excess of dominant alleles and the minority of recessive alleles (p>q). If (4DH1).5 + F/ (4DH1).5 – F <1.0 means minority of dominant alleles and excess of recessive alleles (p<q). Mean covariance of additive and dominant variance was expressed by F. The value of F express if F=0 means balanced distribution (p=q=.5); F>0 (+) means dominant alleles are more frequent than recessive alleles (p>q); F>0 (-) means recessives are more prevalent than dominant alleles (p<q). The proportion of dominant genes with positive or negative effects in parents is determined by the ratio: H2/4H1 with the maximum theoretical value of 0.25, which stands up when p=q=0.5 in all loci. A deviation from 0.25 would stem when p q. Complete dominance was indicated when p = ±1; while partial dominance is indicated when “P” is between (-1 and +1), except the value zero, which indicates absence of dominance. Over-dominance was considered when potency ratio exceeds ±1. The positive and negative signs indicate the direction of dominance of either parent (Pujer and Badiger, 2017). Heritability values were categorized as follows: low, <30%; moderate, 30-60% and high, >60% (Johnson et al., 1955a).
General combining ability (GCA) and specific combining ability (SCA) were estimated following Model I, Method II of Griffing (1956). The standard heterosis (against the best standard check variety) was estimated and tested according to Singh and Singh (1994). Potency ratio was calculated according to Smith (1952) to determine the degree of dominance as follows:
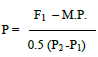
Where, P: relative potency of gene set, F1: first generation mean, P1: the mean of lower parent, P2: the mean of higher parent, M.P.: mid-parents value = (P1 + P2)/2.
RESULTS AND DISCUSSION
Mean performance
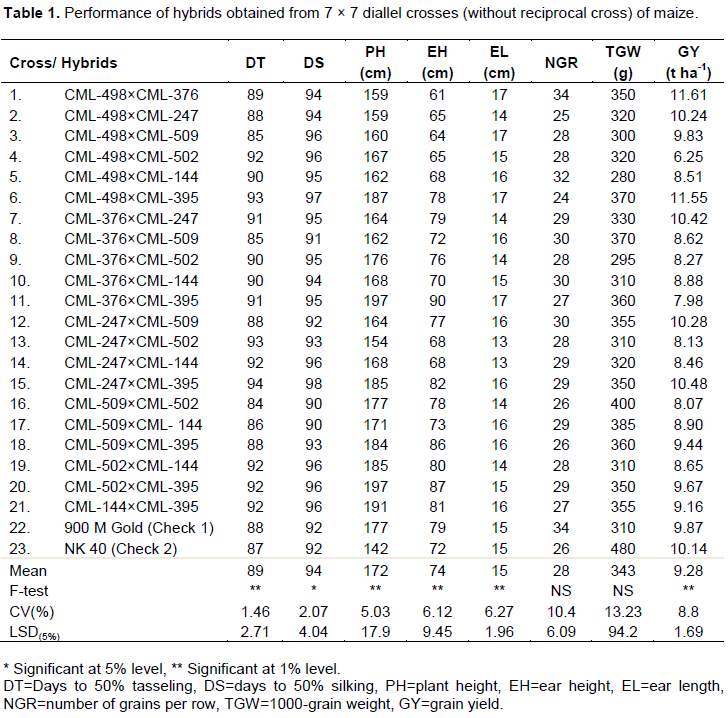
Significant differences were found among the genotypes for days to tasseling (DT), days to silking (DS), plant height (PH), ear height (EH) and grain yield (GY) (Table 1). Though none of the hybrids showed significantly higher yield over the best check NK-40, six cross combinations, CML-498×CML-376 (11.61 t ha-1), CML-498×CML-247 (10.24 t ha-1), CML-498×CML-395 (11.55 t ha-1), CML-376×CML-247 (10.42 t ha-1), CML-247×CML-509 (10.28 t ha-1) and CML-247×CML-395 (10.48 t ha-1) showed better yield than commercial check NK40 (10.14 t ha-1). The highest yielder cross CML-498×CML-376 had higher NGR (34) with shortest EH (61 cm) compared to the best check. Although none of the higher yielder crosses were earlier than the best check, the highest cross was almost similar.
Nature of genetic variance
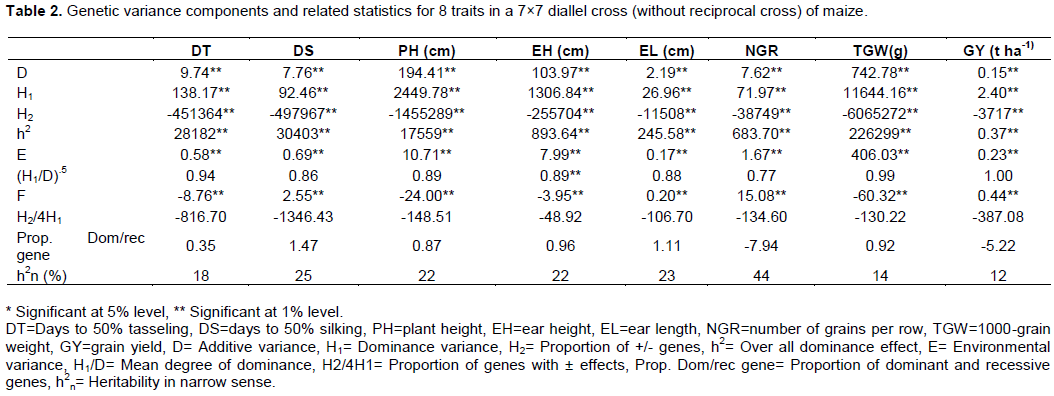
The analysis of genetic variance components indicated that both additive variance (D) and dominance variance (H1 and h2) were significant for all the traits (Table 2). These results focused that the expression of all characters was conditioned by both additive and dominance gene action. However, dominant variance (H1) was more predominant than additive variance (D) for all traits indicating the presence of over dominance controlling the traits (Radha, 2014). The dominance is also reflected by the high degree of dominance effect, that is sum total of all loci in the heterozygous state (h2). The dominance was in partial dominance range because of (H1/D).5 < 1.0 for all traits except grain yield. The dominance is complete dominance in case of grain yield because of (H1/D).5 = 1.0. Though, the environmental variance (E) was significant but much lower than additive variance (D) and dominant variance (H1) for all traits. From proportion of dominant (p) and recessive (q) alleles days to tasseling, plant height, ear height and 1000-grain weight showed asymmetry of distribution (p ≠ q) and the minority of dominant alleles and excess of recessive alleles (p<q) because of the proportion of dominance and recessive gene <1. On the other hand, days to silking, ear length, number of grain per row and grain yield also showed asymmetry of distribution, but excess of dominant alleles and minority of recessive alleles (p>q) as proportion of dominance and recessive gene >1. The symmetry of dominant and recessive allele distribution in parents is further established by relative sizes of dominance variance (H1) and proportion of (+/-) genes (H2) as H1 ≠ H2 means asymmetry of distribution (p ≠ q. In the present study, H1<H2 for all traits indicated an uneven distribution of dominant and recessive alleles. The symmetry of distribution of dominant and recessive alleles in parents is also verified by the direction (sign) of F (mean covariance of D and H1). In the present study, F<0 (-) for days to tasseling, plant height, ear height and 1000-grain weight which indicated recessives were more prevalent than dominant alleles (p<q). On the other hand, F>=0 (+) for days to silking, ear length, number of grain per row and grain yield which means dominant alleles were more frequent than recessive alleles (p>q). The proportion of dominant genes with positive or negative effects in parents is determined by the ratio: H2/4H1 with the maximum theoretical value 0.25. H2/4H1 ≠ 0.25 means asymmetry of the distribution. In the present study, H2/4H1 ≠ 0.25 and negative sign for all studied traits, hence dominant genes having decreasing and increasing effects on all characters were irregularly distributed in parents. Heritability estimate (h2n) was <30% for the studied traits except for number of grain per row (44%) that indicated these traits are less heritable and highly influenced by environment. On the other hand, heritability for number of grain per row was moderate. The predominance of dominant gene action coupled with low heritability observed for days to silking, ear length and grain yield suggesting the importance of heterosis breeding (Radha, 2014).
Analysis of variance
The mean square of genotypes (diallel hybrids) was highly significant for all the traits except 1000-grain weight (Table 3). Further, analysis of variance for combining ability showed that estimates of mean squares due to GCA and SCA were also highly significant for all the characters except 1000-grain weight. This indicated the importance of both additive and non-additive components of genetic variance in controlling these traits. Importance of both GCA and SCA variances for yield and yield contributing traits in maize was reported in various previous studies (Ahmed et al., 2008; Gurung et al., 2008; Mousa, 2014; Hoque et al., 2016). However, in the present study variances due to GCA were much higher in magnitude than SCA for the characters of days to 50% tasseling and silking, ear height, ear length, number of grains per row and 1000-grain weight, which revealed the prevalence of additive gene action for controlling these traits. The predominance of additive gene action for days to tasseling, days to silking and number of grain per row was reported by Hoque et al. (2016) which supports the present study. On the other hand, non additive gene action for ear height and 1000-grain weight was supported by Hoque et al. (2016) and Kadir (2010). On the other hand, the magnitude of SCA was higher than GCA for plant height, number of grain per row and grain yield, indicating non-additive gene action in controlling these traits. Non-additive gene action was also reported on plant height (Kadir, 2010), number of kernel per row (Abdel-Moneam et al., 2009) and grain yield (Abdel-Moneam et al., 2009; Kadir, 2010; Barakat and Osman, 2008; Gouda et al., 2013; Hoque et al., 2016) in their study. These investigations supported the present study.

General combining ability (GCA) effects
The GCA effects were shown in Table 4. None of the parents were found to be a good general combiner for all the characters studied. A wide range of variability in GCA effects was observed among the parents. In case of maize, the inbred lines with significant and negative GCA effects are considered as good general combiners for days to 50% tasseling, days to 50% silking, plant height as well as ear height to utilize these for developing early and short stature plants. On the other hand, for yield and other yield components, those with significant and positive GCA effects are considered as good general combiners.
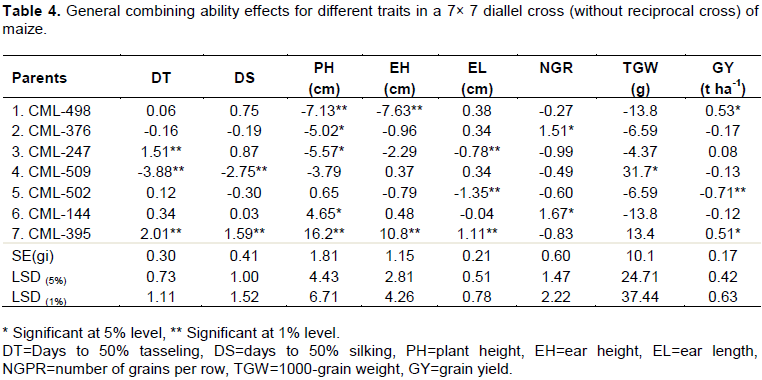
In the present study, the parent CML-509 was a good general combiner for days to tasseling and silking due to its significant negative GCA value. In addition, it was also a good general combiner for 1000-grain weight for its significant positive GCA value. Parent CML-498 and CML-376 showed expected significant negative GCA value for plant height where CML-498 had significant negative value for both plant and ear height. So these parents could be a good source for the development of short stature plant. Significant and negative GCA for ear height was observed in different studies (Malik et al., 2004; Alam et al., 2008; Amiruzzaman, 2010). Inbred CML-498 and CML 395 exhibited significant positive GCA for grain yield. This result was supported by different studies (Malik et al., 2004; Uddin et al., 2006; Ahmed et al., 2008; Abdel-Moneam et al., 2009). They reported that parents with good general combiners for grain yield showed good performance for various yield components.
Higher significant positive GCA for ear length and yield were found in parent CML-395 while in case of number of grains per row and 1000-grain weight, parents CML-144 and CML-509 expressed positive significant GCA. Higher significant and positive GCA effect for 1000-grain weight was also observed in different studies (Alam et al., 2008; Abdel-Moneam et al., 2009; Uddin et al., 2006).
Specific combining ability (SCA) effects
The SCA effects of the crosses for eight characters are presented in Table 5. The hybrid CML-498×CML-247 exhibited significant negative SCA effects for days to tasseling and days to silking. In addition, the hybrid CML-509×CML-502 and CML-247×CML-502 exhibited significant negative SCA effects for days to tasseling and days to silking respectively, indicating earliness of the hybrids. These crosses mostly involved average × low, high × average, low × average general combining parents. These findings are consistent with the results of Ahmed et al. (2008).
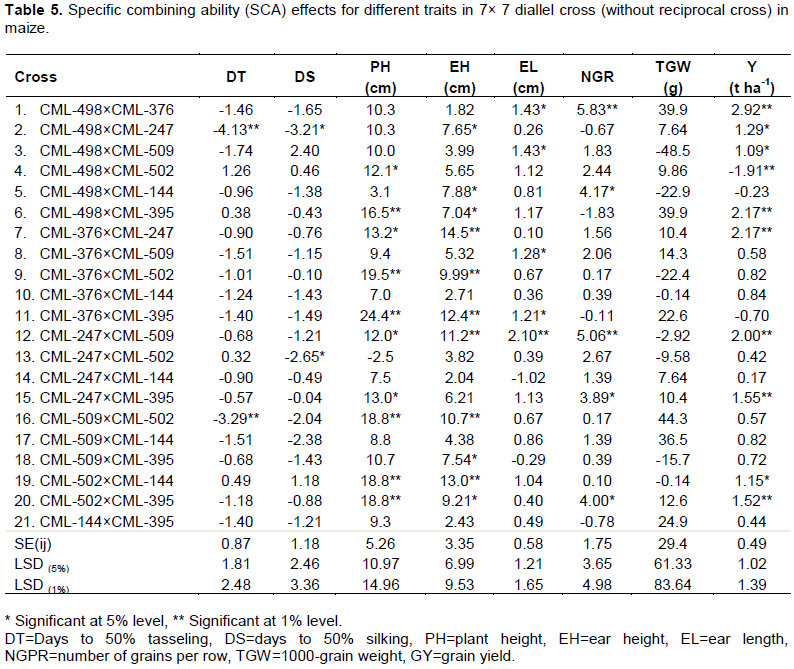
Considering the results, out of 21 hybrids, nine crosses (viz.,CML-498×CML-376, CML-498×CML-247, CML-498×CML-509, CML-498×CML-395, CML-376×CML-247, CML-247×CML-509, CML-247×CML-395, CML-502×CML-144 and CML-502×CML-395) exhibited significant positive SCA effects for grain yield (Table 5), and most of them also possessed high per se performance for the same trait (Table 1). These crosses involved high × high, high × average, high × low, average × high and average × average general combining parents. These crosses involving parents with one or both parents were related to good combiners, indicating GCA of the parental lines plays a key role in producing high yield. Vasal (1998) recommended to include one good combiner (especially female parent) during the crossing to obtain higher heterosis. Xingming et al. (2002) also drew a similar conclusion. On the other hand, an appreciable amount of the SCA effects expressed by low × low crosses might be ascribed to dominance × dominance type of non-allelic gene action produced over-dominance and are non-fixable. It appears that superior performance of most hybrids may be largely due to epistatic interaction. If the inbreed does not show good GCA effect but have good SCA effect, these materials could be successively used for further breeding (Aliu et al., 2009). The SCA effects of the crosses exhibited no specific trends in cross combinations between parents having high, medium and low GCA effects. Any combination of the parents may produce hybrid vigor over the parents which might be due to dominance, over dominance or epistatic gene action. So, the crosses which showing desirable SCA effects can be used in the future breeding program.
None of the crosses exhibited significant and negative SCA effects for plant height and ear height. In case of ear length and number of grains per row, five crosses for each character expressed significant and positive SCA effect. For 1000-grain weight, none of the crosses showed significant and positive SCA effects.
Heterosis and potence ratio
Standard heterosis is important for selecting new variety (Amiruzzaman, 2010; Kadir, 2010). The standard/ economic heterosis expressed by the F1 hybrids over the best commercial check variety NK 40 for yield and yield related traits are shown in Table 6. All the traits showed more or less significant heterosis in different crosses.
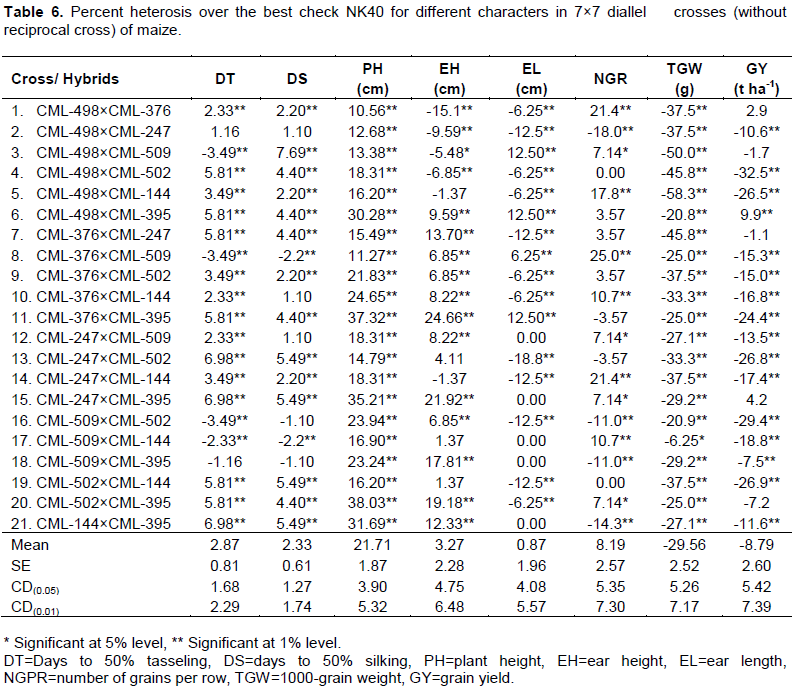
For grain yield (t ha-1), only one cross CML-498×CML-395 (9.9%) showed significant positive heterosis over the standard check variety NK-40. Significant negative heterosis was exhibited by four and two crosses for days to tasseling as well as days to silking respectively, indicating earliness (Table 6). Heterosis ranged from -3.4 to 6.98% and -2.2 to 7.69% for days to tasseling and silking, respectively. Similarly, significant and negative heterosis was exhibited by four crosses for ear height ranging from -15.1 to -5.48%, indicating short stature. All crosses expressed significant and negative heterosis for 1000-grain weight.
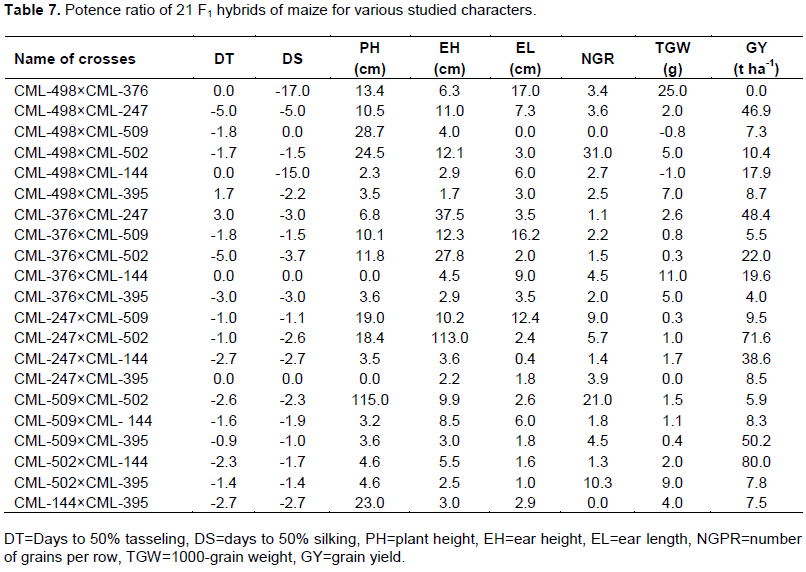
The potency ratio of 21 F1 crosses is presented in Table 7. Positive values ratio specified the degrees of dominance, that is partial to over-dominance and negative values ratio signposted the degrees of recessiveness, that is partial to under recessiveness (Solieman et al., 2013). For Days to 50% tasseling (DT), the potency ratio ranged from -5 (CML-498 × CML-247 and CML-376 × CML-502) to 3 (CML-376 × CML-247). Among them the value of potency ratio was zero (0) for four crosses indicating absence of dominance; two crosses showed complete dominance (-1.0), one cross namely CML-509 × CML-395 exposed partial dominance (-0.9) and the rest 14 crosses exhibited over-dominance (>±1). For days to 50% silking (DS), the range of potency ratio was -17.0 (CML-498 × CML-376) to 0.0 with three crosses showing absence of dominance (0); one cross (CML-509 × CML-395) showed complete dominance (-1) and the rest 17 crosses showed over-dominance (>±1). For plant height (PH), the range of potency ratio was 0.0 (CML-376 × CML-144 and CML-247 × CML-395) to 115 (CML-509 × CML-502), with two crosses showing absence of dominance while the rest 19 crosses showed over-dominance (>±1). For ear height (EH) the range of potency ratio was 1.7 (CML-498 × CML-395) to 113.0 (CML-247 × CML-502) with all crosses exhibiting over-dominance (>+1). For ear length (EL) the range of potency ratio was 0.0 (CML-498 × CML-509) to 17.0 (CML-498 × CML-376) with one cross showing absence of dominance (0), one exhibiting complete dominance (+1) and one exhibiting partial dominance (0.4) and the rest 18 crosses showed over-dominance (>+1). For the number of grain per row (NGR), the range of potency ratio was 0.0 (CML-498 × CML-509 and CML-144 × CML-395) to 31.0 (CML-498 × CML-502) with two crosses showing absence of dominance and the rest 19 crosses showed over-dominance (>+1). For 1000-grain weight, the range of potency ratio was 0.0 (CML-247 × CML-395) to 25 (CML-498 × CML-376) with two crosses showing complete dominance (±1), one showed absence of dominance, five crosses showed partial dominance (-1 to +1) and the rest 13 crosses showed over-dominance. For grain yield (GY) the range of potency ratio was 0.0 (CML-498 × CML-376) to 80 (CML-502 × CML-144) with only one cross showing absence of dominance and the rest 20 cross exhibited over dominance.
CONCLUSION
The results of the study revealed that the value of dominance variance (H1) and the proportion of +/- genes (H2) were higher than additive variance (D) in all characters. Therefore, over-dominance controlled the studied traits. Hybrids projecting positive or negative potence ratio with >1.0 value for those traits is the sign of incidence of over dominance in desirable direction and heterosis breeding is important to improve those traits in maize. The parental lines CML-498 and CML-395 were found to be the best general combiner for yield. The good combiner parents for different trait could be used in hybridization to improve yield as well as with other desirable traits as donor parents for the accumulation of favorable genes. Three hybrids namely, CML-498×CML-376, CML-498×CML-395 and CML-376×CML-247 need to be further evaluated at different agro-ecological conditions in a multi-year to evaluate their performance.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Abdel-Moneam MA, Attia AN, El-Emery MI, Fayed EA (2009). Combining ability and heterosis for some agronomic traits in crosses of maize. Pakistan Journal of Biological Science 12(5):433-438. |
|
|
Ahmed S, Khatun F, Uddin MS, Banik BR, Ivy NA (2008). Combining ability and heterosis in maize (Zea mays L.). Bangladesh Journal of Plant Breeding and Genetics 21(2):27-32. |
|
|
Alam AKMM, Ahmed S, Begum M, Sultan MK (2008). Heterosis and combining ability for grain yield and its contributing characters in maize. Bangladesh Journal of Agricultural Research 33(3):375-379. |
|
|
Aliu S, Fetahu S, Kaciu S, Salillari A (2009). Combining ability study for kernel yield per ear of maize (Zea mays L.) hybrid. 44th Croatian and 4th International Symposium on Agriculture pp. 476-480. |
|
|
Amiruzzaman M (2010). Exploitation of hybrid vigor from normal and quality protein maize crosses. Ph.D Dissertation, Dept. Genetics and Plant Breeding, Bangladesh Agricultural University, Mymensingh P 200. |
|
|
Barakat AA, Osman MMA (2008). Evaluation of some newly developed yellow maize inbred lines for combining ability in two locations. Journal of Agricultural Science Mansoura University 33:4667-4679. |
|
|
Bocanski J, Zorana S, Aleksandra N (2009). Genetic and phenotypic relationship between grain yield and components of grain yield of maize (Zea mays L.). Original science, paper. Genetika 41:145-154. |
|
|
Brahmbhatt BN, Kuchhadiya GV, Gosai MA, Joshi NR, Kanjariya KG (2018). Study of combining ability through diallel crosses in maize (Zea mays L.) for grain yield and protein content. Journal of Pharmacognosy and Phytochemistry 7(2):1754-1755. |
|
|
Edwards LH, Ketata H, Smith EL (1976). Gene action of heading date, plant height, and other characters in tow winter wheat crosses. Crop Science 16:275-277. |
|
|
Falconer DC (1981). Introduction to quantitative Genetics, 2nd edition, Longman Group Ltd. New York, 67-68. |
|
|
Food and Agriculture Organization (FAO) (2016). Agriculture Organization of the United Nations Statistics Division. |
|
|
Gouda RK, Kage U, Lohithaswa HC, Shekara BG, Shobha D (2013). Combining ability studies in maize (Zea mays L.). Molecular Plant Breeding 3:116-127. |
|
|
Griffing B (1956). Concept of general and specific combining ability in relation to diallel crossing systems. Australian Journal of Biological Science 9:463-493. |
|
|
Gurung DB, George MLC, Delacruz QD (2008). Heterosis and combining ability of Nepalese Yellow maize Populations. Book of Abstracts. The 10th Asian Reg. Maize Workshop. October 20-23, Makassar, Indonesia P 88. |
|
|
Hayman BI (1954). The analysis of variance of diallel tables. Biometrics 10:235-244. |
|
|
Hoque M, Akhter F, Kadir M, Begum HA, Ahmed S (2016). Study on combining ability and heterosis for earliness and short statured plant in maize. Bangladesh Journal of Agricultural Research 41(2):365-376. |
|
|
Huang M, Chen L, Chen Z (2015). Diallel analysis of combining ability and heterosis for yield and yield components in rice by using positive loci. Euphytica 205:37. |
|
|
Jinks JL, Hayman BL (1963). The analysis of diallel crosses. Maize Genetic Cooperative Newsletter 27:48:54. |
|
|
Johnson HW, Robinson HF, Comstock RE (1955). Estimates of genetic and environmental variability in soybean. Journal of Agronomy 47:314-318. |
|
|
Kadir MM (2010). Development of quality protein maize hybrids and their adaptation in Bangladesh. PhD Dissertation, Dept. Genetics & Plant Breeding, Bangladesh Agricultural University, Mymensingh. |
|
|
Kundan K, Shailesh M, Binod K (2013). Combining ability analysis in diallel crosses of wheat (Triticum aestivum L). The Bioscan 8:1557-1560. |
|
|
Malik SI, Malik HN, Minhas NM, Munir M (2004). General and specific combining ability studies in maize diallel crosses. International Journal of Agriculture and Biology 6(5):856-859. |
|
|
Mousa STM (2014). Diallel analysis for physiological traits and grain yield of seven white maize inbred lines. Alexandria Journal of Agricultural Research 59(1):9-17. |
|
|
Njeri SG, Makumbi D, Warburton ML, Diallo A, Jumbo MB, Chemining'wa G (2017). Genetic analysis of tropical quality protein maize (Zea mays L.) germplasm. Euphytica 213:261 |
|
|
Owusu GA, Nyadanu D, Obeng-Antwi K, Amoah RA, Danso FC, Amissah S (2017). Estimating gene action, combining ability and heterosis for grain yield and agronomic traits in extra-early maturing yellow maize single-crosses under three agro-ecologies of Ghana. Euphytica 213: 287. |
|
|
Pujer P, Badiger M (2017). Heterosis and potence ratios for growth, earliness, yield and quality traits in cherry tomato (Solanum lycopersicum L. var. Cerasiforme Mill). International Journal of Chemical Studies 5(4):1000-1006. |
|
|
Radha RK (2014). Diallel analysis for different horticultural traits in bitter gourd (Momordica charantia L.) using Hayman's numerical and graphical approach. Tropical Plant Research 1(2):60-64. |
|
|
Reza A, Yazdisamadi B, Zali A, Tallei A, Zeinali H, Rezael A (2004). Estimate of heterosis and combining ability in maize (Zea mays L.) using diallel crossing method. pp. 395-397. In: Genetic Variation for plant breeding. 2004 EUCARPIA, BOKU, Vienna, Austria. |
|
|
Sheikh FA, Dar ZA, Sofi PA, Lone AA (2017). Recent Advances in Breeding for Abiotic Stress (Drought) Tolerance in Maize. International Journal of Current Microbiology and Applied Sciences 6(4):2226-2243. |
|
|
Singh RK, Singh PK (1994). A manual on Genetics and Plant Breeding. Experimental Techniques. Kalyani Publishers. Ludiana, New Delhi pp. 99-107. |
|
|
Smith HH (1952). Fixing Transgressive Vigour in Nicotiana rustica. In: Gowen JW, editor. Heterosis. Ames (IA), Iowa State University Press. pp. 161-174. |
|
|
Solieman THI, El-Gabry MAH, Abido AI (2013). Heterosis, potence ratio and correlation of some important characters in tomato (Solanum lycopersicon L.). Scientia Horticulturae 150:25-30. |
|
|
Sprague GF, Tatum LA (1942). General vs. specific combining ability in single crosses of corn. Journal of American Society of Agronomy 34:923-932. |
|
|
Tian HY, Channa SA, Hu SW (2017). Relationships between genetic distance, combining ability and heterosis in rapeseed (Brassica napus L.) Euphytica 213:1. |
|
|
Tumuhimbise R, Melis R, Shanahan P (2014). Diallel analysis of early storage root yield and disease resistance traits in cassava (Manihot esculenta Crantz). Field Crops Research 167:86-93. |
|
|
Uddin SM, Khatun F, Ahmed S, Ali MR, Begum SA (2006). Heterosis and combining ability in corn (Zea mays L.). Bangladesh Journal of Botany 35:109-116. |
|
|
Vasal SK (1998). Hybrid maize technology: Challenges and expanding possibilities for research in the next century. In: Vasal SK, Gonzalez CF, Xingming F (ed). Proc. 7th Asian Reg. Maize workshop.Los Banos, Philippines, February 23-27, pp.58-62. |
|
|
Walters DS, Morton JR (1978). On the analysis of variance of a half diallel table. Biometrics 34:91-94. |
|
|
Xingming F, Tan J, Chen Z, Yang J (2002). Combining ability and heterotic grouping of ten temperate, tropical and subtropical quality protein maize. In: Srinivasan, G, Zaidi PH, Prasanna BN, Gonzalez FC, Lesnick K (ed). Proc. 8th Asian Reg. Maize Workshop. Bangkok, Thailand, August 5-8, pp. 10-18. |
|
|
Yan W, Hunt LA (2002). Biplot analysis of diallel data. Crop Science 42: 21-30. |
|
|
Zare M, Choukan R, Bihamta MR, Heravan EM, Kamelmanesh MM (2011). Gene action for some agronomic traits in maize (Zea mays L.). Crop Breeding Journal 1(2):133-141. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


